Effects of Wheat Bran Applied to Maternal Diet on the Intestinal Architecture and Immune Gene Expression in Suckling Piglets

 , , , , , , ,
, , , , , , ,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals and Experimental Design
2.3. qPCR
2.4. Statistical Analyses
3. Results
3.1. Zootechnical Results
3.2. Milk Composition
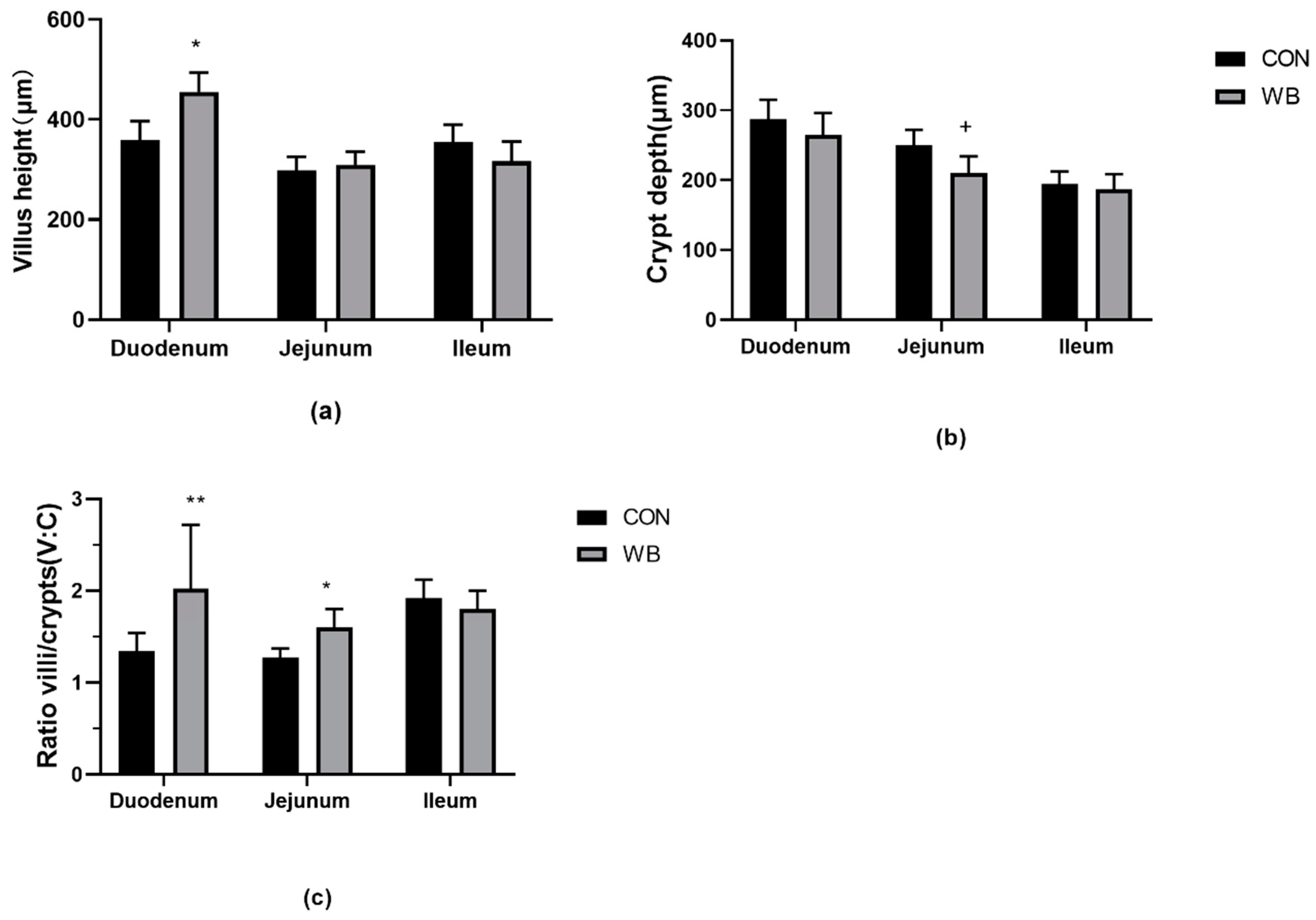
3.3. Histomorphology Analysis of the Small Intestine
3.4. Relative Gene Expression in Ileum and Colon Mucosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Annison, G. The role of wheat non-starch polysaccharides in broiler nutrition. Aust. J. Agric. Res. 1993, 44, 405–422. [Google Scholar] [CrossRef]
- Jha, R.; Rossnagel, B.; Pieper, R.; Van Kessel, A.; Leterme, P. Barley and oat cultivars with diverse carbohydrate composition alter ileal and total tract nutrient digestibility and fermentation metabolites in weaned piglets. Animal 2010, 4, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Leterme, P. Feed ingredients differing in fermentable fibre and indigestible protein content affect fermentation metabolites and faecal nitrogen excretion in growing pigs. Animal 2012, 6, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Berrocoso, J. Dietary fiber utilization and its effects on physiological functions and gut health of swine. Animal 2015, 9, 1441–1452. [Google Scholar] [CrossRef]
- Saqui-Salces, M.; Huang, Z.; Vila, M.F.; Li, J.; Mielke, J.A.; Urriola, P.E.; Shurson, G.C. Modulation of intestinal cell differentiation in growing pigs is dependent on the fiber source in the diet1. J. Anim. Sci. 2017, 95, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Molist, F.; de Segura, A.G.; Pérez, J.; Bhandari, S.; Krause, D.; Nyachoti, C. Effect of wheat bran on the health and performance of weaned pigs challenged with Escherichia coli K88+. Livest. Sci. 2010, 133, 214–217. [Google Scholar] [CrossRef]
- Chen, H.; Mao, X.; He, J.; Yu, B.; Huang, Z.; Yu, J.; Zheng, P.; Chen, D. Dietary fibre affects intestinal mucosal barrier function and regulates intestinal bacteria in weaning piglets. Br. J. Nutr. 2013, 110, 1837–1848. [Google Scholar] [CrossRef]
- Chen, H.; Chen, D.; Qin, W.; Liu, Y.; Che, L.; Huang, Z.; Luo, Y.; Zhang, Q.; Lin, D.; Liu, Y.; et al. Wheat bran components modulate intestinal bacteria and gene expression of barrier function relevant proteins in a piglet model. Int. J. Food Sci. Nutr. 2017, 68, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Van Craeyveld, V.; Swennen, K.; Dornez, E.; Van de Wiele, T.; Marzorati, M.; Verstraete, W.; Delaedt, Y.; Onagbesan, O.; Decuypere, E.; Buyse, J.; et al. Structurally different wheat-derived arabinoxylooligosaccharides have different prebiotic and fermentation properties in rats. Livest. Sci. 2008, 138, 2348–2355. [Google Scholar] [CrossRef]
- Koo, B.; Hossain, M.M.; Nyachoti, C.M. Effect of dietarywheat bran inclusion on nutrient and energy digestibility and microbial metabolites in weaned pigs. Livest. Sci. 2017, 203, 110–113. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, P.; Wu, Y.; Guo, P.; Liu, L.; Ma, N.; Levesque, C.; Chen, Y.; Zhao, J.; Zhang, J. Dietary Fiber Increases Butyrate-Producing Bacteria and Improves the Growth Performance of Weaned Piglets. J. Agric. Food. Chem. 2018, 66, 7995–8004. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Van Hée, V.F.; Piront, N.; De Backer, F.; Toussaint, O.; Cani, P.D.; Delzenne, N.M. Wheat-derived arabinoxylan oligosaccharides with prebiotic effect increase satietogenic gut peptides and reduce metabolic endotoxemia in diet-induced obese mice. Nutr. Diabetes 2012, 2, e28. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.; Ashworth, C.J. The role of dietary fibre in pig production, with a particular emphasis on reproduction. J. Anim. Sci. Biotechnol. 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Feng, C.; Tao, S.; Li, N.; Zuo, B.; Han, D.; Wang, J. Maternal imprinting of the neonatal microbiota colonization in intrauterine growth restricted piglets: A review. J. Anim. Sci. Biotechnol. 2019, 10, 88. [Google Scholar] [CrossRef]
- Leblois, J.; Massart, S.; Li, B.; Wavreille, J.; Bindelle, J.; Everaert, N. Modulation of piglets’ microbiota: Differential effects by a high wheat bran maternal diet during gestation and lactation. Sci. Rep. 2017, 7, 7426. [Google Scholar] [CrossRef] [PubMed]
- Schokker, D.; Zhang, J.; Zhang, L.L.; Vastenhouw, S.A.; Heilig, H.G.H.J.; Smidt, H.; Rebel, J.M.J.; Smits, M.A. Early-life environmental variation affects intestinal microbiota and immune development in new-born piglets. PLoS ONE 2014, 9, e100040. [Google Scholar] [CrossRef]
- Everaert, N.; Van Cruchten, S.; Weström, B.; Bailey, M.; Van Ginneken, C.; Thymann, T.; Pieper, R. A review on early gut maturation and colonization in pigs, including biological and dietary factors affecting gut homeostasis. Anim. Feed Sci. Technol. 2017, 233, 89–103. [Google Scholar] [CrossRef]
- Leonard, S.; Sweeney, T.; Bahar, B.; O’Doherty, J. Effect of maternal seaweed extract supplementation on suckling piglet growth, humoral immunity, selected microflora, and immune response after an ex vivo lipopolysaccharide challenge. J. Anim. Sci. 2011, 90, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Liu, H.; Liu, S.; He, T.; Piao, X. Effects of dietary fiber sources during late gestation and lactation on sow performance, milk quality, and intestinal health in piglets1. J. Anim. Sci. 2019, 97, 4922–4933. [Google Scholar] [CrossRef]
- Leblois, J.; Massart, S.; Soyeurt, H.; Grelet, C.; Dehareng, F.; Schroyen, M.; Li, B.; Wavreille, J.; Bindelle, J.; Everaert, N. Feeding sows resistant starch during gestation and lactation impacts their faecal microbiota and milk composition but shows limited effects on their progeny. PLoS ONE 2018, 13, e0199568. [Google Scholar] [CrossRef]
- Uerlings, J.; Schroyen, M.; Bautil, A.; Courtin, C.; Richel, A.; Sureda, E.A.; Bruggeman, G.; Tanghe, S.; Willems, E.; Bindelle, J. In vitro prebiotic potential of agricultural by-products on intestinal fermentation, gut barrier and inflammatory status of piglets. Br. J. Nutr. 2020, 123, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Loisel, F.; Farmer, C.; Ramaekers, P.; Quesnel, H. Effects of high fiber intake during late pregnancy on sow physiology, colostrum production, and piglet performance1. J. Anim. Sci. 2013, 91, 5269–5279. [Google Scholar] [CrossRef]
- Guillemet, R.; Hamard, A.; Quesnel, H.; Père, M.C.; Etienne, M.; Dourmad, J.Y.; Meunier-Salaün, M.C. Dietary fibre for gestating sows: Effects on parturition progress, behaviour, litter and sow performance. Animal 2007, 1, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.P.; Johnston, L.J.; Baidoo, S.K.; Shurson, G.C. Effects of a high-fiber diet and frequent feeding on behavior, reproductive performance, and nutrient digestibility in gestating sows. J. Anim. Sci. 2006, 84, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Hedemann, M.S.; Eskildsen, M.; Lærke, H.N.; Pedersen, C.; Lindberg, J.E.; Laurinen, P.; Bach Knudsen, K.E. Intestinal morphology and enzymatic activity in newly weaned pigs fed contrasting fiber concentrations and fiber properties. J. Anim. Sci. 2006, 84, 1375–1386. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Liu, H.; Yang, Y.; He, J.; Cao, M.; Yang, M.; Zhong, W.; Lin, Y.; Zhuo, Y.; et al. Effects of the Ratio of Insoluble Fiber to Soluble Fiber in Gestation Diets on Sow Performance and Offspring Intestinal Development. Animals 2019, 9, 422. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Naik, S. Compartmentalized and systemic control of tissue immunity by commensals. Nat. Immunol. 2013, 14, 646–653. [Google Scholar] [CrossRef]
- Xu, H.; Hu, M.B.; Bai, P.D.; Zhu, W.H.; Liu, S.H.; Hou, J.Y.; Xiong, Z.Q.; Ding, Q.; Jiang, H.W. Pro-inflammatory cytokines in prostate cancer development and progression promoted by high-fat diet. BioMed. Res. Int. 2015, 249741. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. (Landmark Ed.) 2009, 14, 2765–2778. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani, A.I.O.; Beswa, D. Composition and functionality of wheat bran and its application in some cereal food products. Int. J. Food Sci. Tech. 2015, 50, 2509–2518. [Google Scholar] [CrossRef]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-γ agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef]
- Opal, S.M.; DePalo, V.A.J.C. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta. 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Xue, X.; Falcon, D.M. The Role of Immune Cells and Cytokines in Intestinal Wound Healing. Int. J. Mol. Sci. 2019, 20, 6097. [Google Scholar] [CrossRef] [PubMed]
- Dann, S.M.; Spehlmann, M.E.; Hammond, D.C.; Iimura, M.; Hase, K.; Choi, L.J.; Hanson, E.; Eckmann, L. IL-6-dependent mucosal protection prevents establishment of a microbial niche for attaching/effacing lesion-forming enteric bacterial pathogens. J. Immunol. 2008, 180, 6816–6826. [Google Scholar] [CrossRef]


{kind=link}
| Sample | Treatment | Parity | Protein (%) | Fat (%) | Lactose (%) | IgA (mg/mL) | IgG (mg/mL) | IgM (mg/mL) |
|---|---|---|---|---|---|---|---|---|
| Colostrum | CON | 1 | 19.49 | 6.48 | 2.6 | 12.63 | 68.58 | 4.99 |
| CON | ≥2 | 18.77 | 6.21 | 2.6 | 14.81 | 59.74 | 4.73 | |
| WB | 1 | 18.36 | 6.63 | 2.66 | 11.48 | 65.69 | 4.23 | |
| WB | ≥2 | 20.4 | 6.08 | 2.56 | 15.25 | 71.31 | 4.21 | |
| Global SEM | 0.3 | 0.14 | 0.03 | 0.81 | 3.46 | 0.32 | ||
| Milk W1 | CON | 1 | 6.1 | 10.26 | 4.75 | 1.84 | 0.42 | 0.92 |
| CON | ≥2 | 6.22 | 9.29 | 4.59 | 2.3 | 0.39 | 1.21 | |
| WB | 1 | 5.83 | 10.37 | 4.71 | 2.49 | 0.34 | 0.95 | |
| WB | ≥2 | 6.1 | 8.59 | 4.77 | 2.57 | 0.48 | 1.35 | |
| Global SEM | 0.1 | 0.51 | 0.05 | 0.16 | 0.05 | 0.11 | ||
| Milk W2 | CON | 1 | 5.95 | 10.93 | 4.79 | 2.26 | 0.27 | 1.1 |
| CON | ≥2 | 6.1 | 9.02 | 4.82 | 2.55 | 0.32 | 1.09 | |
| WB | 1 | 5.43 | 9.68 | 4.85 | 2.36 | 0.24 | 0.97 | |
| WB | ≥2 | 5.8 | 9.55 | 4.93 | 2.96 | 0.27 | 1.19 | |
| Global SEM | 0.1 | 0.28 | 0.02 | 0.17 | 0.02 | 0.07 | ||
| Milk W3 | CON | 1 | 6.31 | 10.84 | 4.81 | 3.22 | 0.21 | 1.01 |
| CON | ≥2 | 6.15 | 8.88 | 4.84 | 3.56 | 0.2 | 0.95 | |
| WB | 1 | 5.74 | 8.31 | 4.95 | 3.77 | 0.16 | 0.98 | |
| WB | ≥2 | 5.98 | 9.4 | 4.91 | 3.37 | 0.16 | 1.03 | |
| Global SEM | 0.08 | 0.38 | 0.03 | 0.23 | 0.02 | 0.08 | ||
| Overall p-values | ||||||||
| Treatment | 0.16 | 0.23 | <0.05 | 0.77 | 0.52 | 0.58 | ||
| Time | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
| Parity | 0.04 | 0.03 | 0.75 | 0.06 | 0.82 | 0.76 | ||
| Treatment*Week | 0.48 | 0.48 | 0.74 | 0.68 | 0.8 | 0.31 | ||
| Treatment*Parity | <0.01 | 0.16 | 0.58 | 0.82 | 0.28 | 0.73 | ||
| Time*Parity | 0.4 | 0.63 | 0.2 | 0.03 | 0.34 | 0.19 | ||
| Treatment*Time*Parity | 0.12 | 0.06 | 0.25 | 0.04 | 0.28 | 0.9 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leblois, J.; Zhang, Y.; Wavreille, J.; Uerlings, J.; Schroyen, M.; Sureda, E.A.; Soyeurt, H.; Dehareng, F.; Grelet, C.; Oswald, I.P.; et al. Effects of Wheat Bran Applied to Maternal Diet on the Intestinal Architecture and Immune Gene Expression in Suckling Piglets. Animals 2020, 10, 2051. https://doi.org/10.3390/ani10112051
Leblois J, Zhang Y, Wavreille J, Uerlings J, Schroyen M, Sureda EA, Soyeurt H, Dehareng F, Grelet C, Oswald IP, et al. Effects of Wheat Bran Applied to Maternal Diet on the Intestinal Architecture and Immune Gene Expression in Suckling Piglets. Animals. 2020; 10(11):2051. https://doi.org/10.3390/ani10112051
Chicago/Turabian StyleLeblois, Julie, Yuping Zhang, José Wavreille, Julie Uerlings, Martine Schroyen, Ester Arévalo Sureda, Hélène Soyeurt, Frédéric Dehareng, Clément Grelet, Isabelle P. Oswald, and et al. 2020. "Effects of Wheat Bran Applied to Maternal Diet on the Intestinal Architecture and Immune Gene Expression in Suckling Piglets" Animals 10, no. 11: 2051. https://doi.org/10.3390/ani10112051
APA StyleLeblois, J., Zhang, Y., Wavreille, J., Uerlings, J., Schroyen, M., Sureda, E. A., Soyeurt, H., Dehareng, F., Grelet, C., Oswald, I. P., Li, B., Bindelle, J., Zhang, H., & Everaert, N. (2020). Effects of Wheat Bran Applied to Maternal Diet on the Intestinal Architecture and Immune Gene Expression in Suckling Piglets. Animals, 10(11), 2051. https://doi.org/10.3390/ani10112051