Genetic Analysis of Milk Production Traits and Mid-Infrared Spectra in Chinese Holstein Population

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Original Data
2.2. Data Editing
2.3. Models for Estimates of Non-Genetic Factors
2.4. Models for Genetic Analyses
3. Results and Discussion
3.1. Descriptive Statistics
3.2. Estimates of Fixed Effects on Milk Production Traits
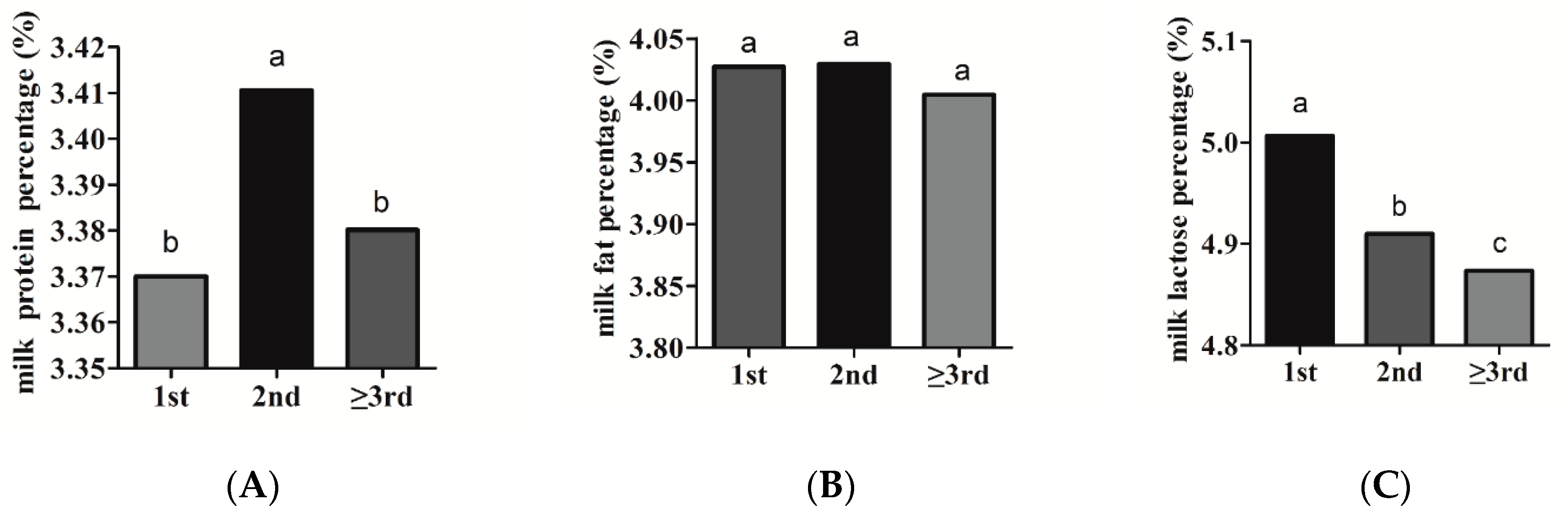
3.2.1. Parity
3.2.2. Calving Season
3.2.3. Lactation Stage
3.3. Estimates of Fixed Effects on Milk Infrared Spectra
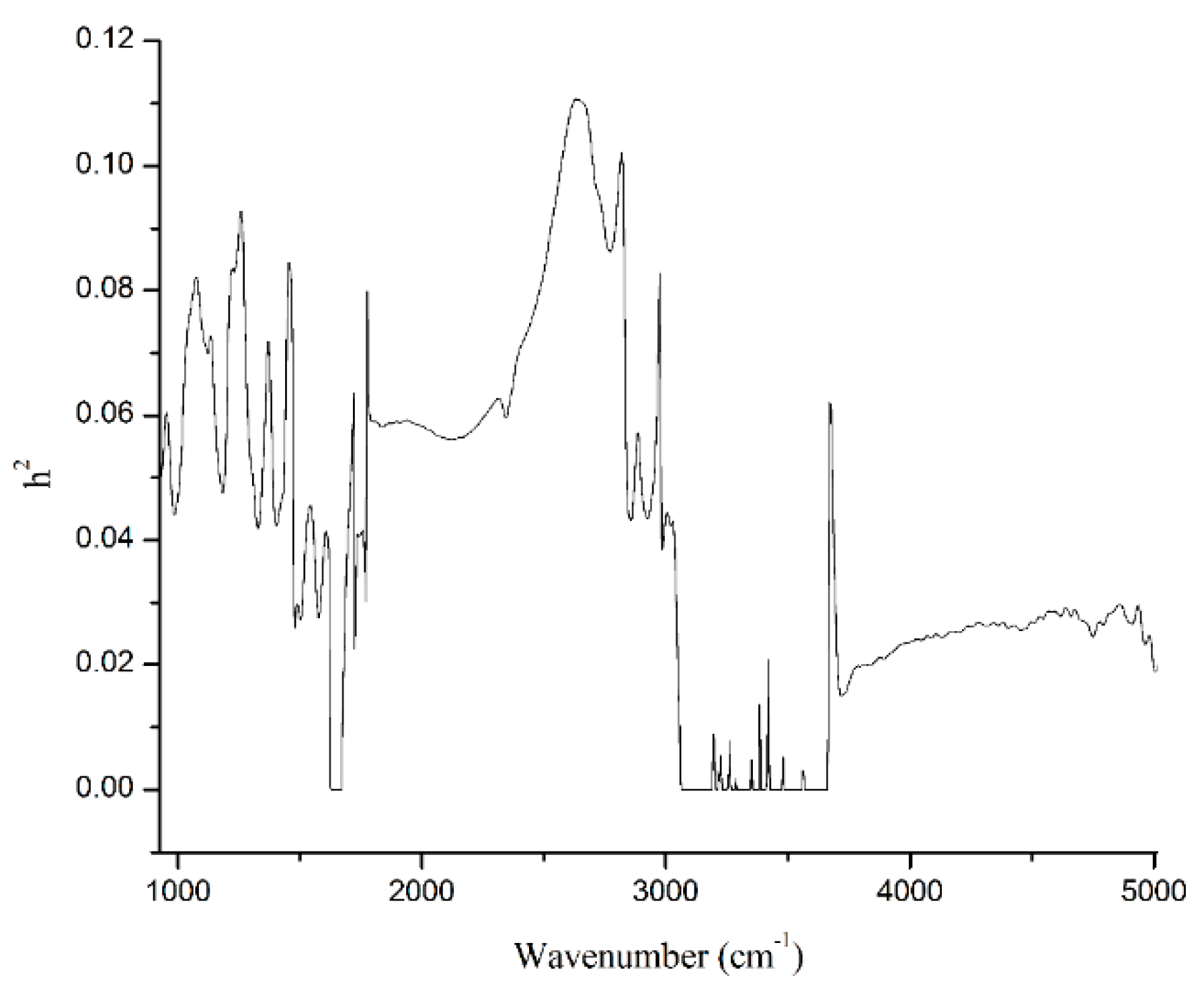
3.4. Heritability
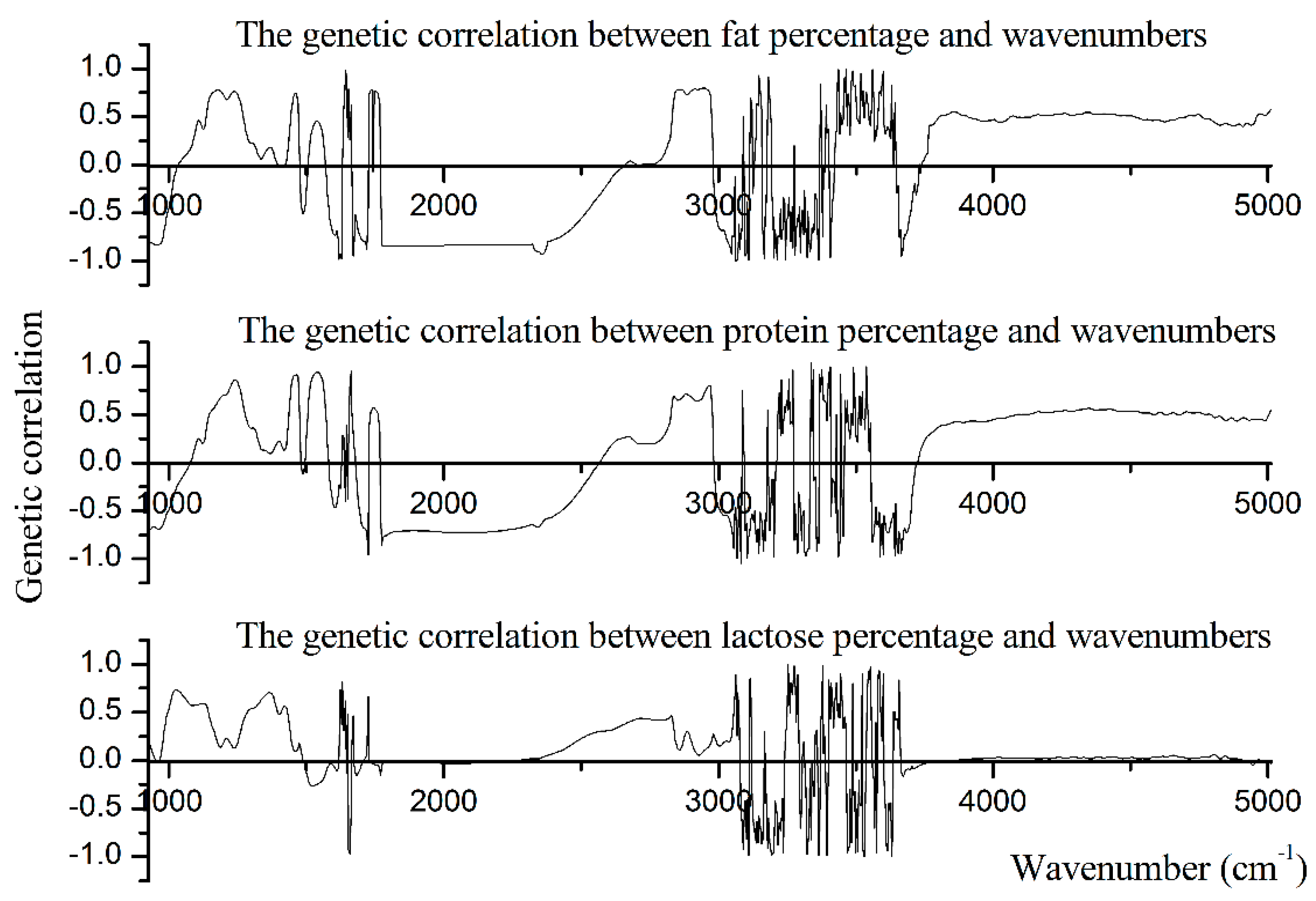
3.5. Genetic Correlation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Statements
References
- De Marchi, M.; Toffanin, V.; Cassandro, M.; Penasa, M. Invited review: Mid-infrared spectroscopy as phenotyping tool for milk traits. J. Dairy Sci. 2014, 97, 1171–1186. [Google Scholar] [CrossRef] [PubMed]
- Gottardo, P.; De Marchi, M.; Cassandro, M.; Penasa, M. Technical note: Improving the accuracy of mid-infrared prediction models by selecting the most informative wavelengths. J. Dairy Sci. 2015, 98, 4168–4173. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.; Visentin, G.; De Marchi, M.; Berry, D.P.; Fenelon, M.A.; O’Connor, P.M.; Kenny, O.A.; McParland, S. Prediction of individual milk proteins including free amino acids in bovine milk using mid-infrared spectroscopy and their correlations with milk processing characteristics. J. Dairy Sci. 2016, 99, 3171–3182. [Google Scholar] [CrossRef] [PubMed]
- Margolies, B.J.; Barbano, D.M. Determination of fat, protein, moisture, and salt content of Cheddar cheese using mid-infrared transmittance spectroscopy. J. Dairy Sci. 2018, 101, 924–933. [Google Scholar] [CrossRef]
- De Marchi, M.; Penasa, M.; Cecchinato, A.; Mele, M.; Secchiari, P.; Bittante, G. Effectiveness of mid-infrared spectroscopy to predict fatty acid composition of Brown Swiss bovine milk. Animal 2011, 5, 1653–1658. [Google Scholar] [CrossRef]
- Visentin, G.; Penasa, M.; Niero, G.; Cassandro, M.; De March, M. Phenotypic characterisation of major mineral composition predicted by mid-infraredspectroscopy in cow milk. Italian J. Anim. Sci. 2018, 17, 549–556. [Google Scholar] [CrossRef]
- Vanlierde, A.; Vanrobays, M.L.; Dehareng, F.; Froidmont, E.; Soyeurt, H.; McParland, S.; Lewis, E.; Deighton, M.H.; Grandl, F.; Kreuzer, M.; et al. Hot topic: Innovative lactation-stage-dependent prediction of methane emissions from milk mid-infrared spectra. J. Dairy Sci. 2015, 98, 5740–5747. [Google Scholar] [CrossRef]
- McParland, S.; Kennedy, E.; Lewis, E.; Moore, S.G.; McCarthy, B.; O’Donovan, M.; Berry, D.P. Genetic parameters of dairy cow energy intake and body energy status predicted using mid-infrared spectrometry of milk. J. Dairy Sci. 2015, 98, 1310–1320. [Google Scholar] [CrossRef]
- Klaffenböck, M.; Steinwidder, A.; Fasching, C.; Terler, G.; Gruber, L.; Mészáros, G.; Sölkner, J. The use of mid-infrared spectrometry to estimate the ration composition of lactating dairy cows. J. Dairy Sci. 2017, 100, 5411–5421. [Google Scholar] [CrossRef]
- Norup, L.R.; Hansen, P.W.; Ingvartsen, K.L.; Friggens, N.C. An attempt to detect estrus from changes in Fourier transform infrared spectra of milk from dairy heifers. Anim. Reprod. Sci. 2001, 65, 43–50. [Google Scholar] [CrossRef]
- Martin, A.D.; Afseth, N.K.; Kohler, A.; Randby, Å.; Eknæs, M.; Waldmann, A.; Dørum, G., 2nd; Måge, I.; Reksen, O. The relationship between fatty acid profiles in milk identified by Fourier transform infrared spectroscopy and onset of luteal activity in Norwegian dairy cattle. J. Dairy Sci. 2015, 98, 5374–5384. [Google Scholar] [CrossRef]
- Lainé, A.; Bastin, C.; Grelet, C.; Hammami, H.; Colinet, F.G.; Dale, L.M.; Gillon, A.; Vandenplas, J.; Dehareng, F.; Gengler, N. Assessing the effect of pregnancy stage on milk composition of dairy cows using mid-infrared spectra. J. Dairy Sci. 2017, 100, 2863–2876. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Alvarado, H.; Vazquez, A.I.; De Los Campos, G.; Tempelman, R.J.; Bittante, G.; Cecchinato, A. Diagnosing pregnancy status using infrared spectra and milk composition in dairy cows. J. Dairy Sci. 2018, 101, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Soyeurt, H.; Misztal, I.; Gengler, N. Genetic variability of milk components based on mid-infrared spectral data. J. Dairy Sci. 2010, 93, 1722–1728. [Google Scholar] [CrossRef]
- Bittante, G.; Cecchinato, A. Genetic analysis of the Fourier-transform infrared spectra of bovine milk with emphasis on individual wavelengths related to specific chemical bonds. J. Dairy Sci. 2013, 96, 5991–6006. [Google Scholar] [CrossRef]
- Wang, Q.; Hulzebosch, A.; Bovenhuis, H. Genetic and environmental variation in bovine milk infrared spectra. J. Dairy Sci. 2016, 99, 6793–6803. [Google Scholar] [CrossRef] [PubMed]
- Zaalberg, R.M.; Shetty, N.; Janss, L.; Buitenhuis, A.J. Genetic analysis of Fourier transform infrared milk spectra in Danish Holstein and Danish Jersey. J. Dairy Sci. 2019, 102, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Coffey, E.L.; Horan, B.; Evans, R.D.; Berry, D.P. Milk production and fertility performance of Holstein, Friesian, and Jersey purebred cows and their respective crosses in seasonal-calving commercial farms. J. Dairy Sci. 2016, 99, 5681–5689. [Google Scholar] [CrossRef] [PubMed]
- Mele, M.; Macciotta, N.P.P.; Cecchinato, A.; Conte, G.; Schiavon, S.; Bittante, G. Multivariate factor analysis of detailed milk fatty acid profile: Effects of dairy system, feeding, herd, parity, and stage of lactation. J. Dairy Sci. 2016, 99, 9820–9833. [Google Scholar] [CrossRef] [PubMed]
- Bovenhuis, H.; Visker, M.H.; van Valenberg, H.J.; Buitenhuis, A.J.; van Arendonk, J.A. Effects of the DGAT1 polymorphism on test-day milk production traits throughout lactation. J. Dairy Sci. 2015, 98, 6572–6582. [Google Scholar] [CrossRef]
- Zhao, F.P.; Guo, G.; Wang, Y.C.; Guo, X.Y.; Zhang, Y.; Du, L.X. Genetic parameters for somatic cell score and production traits in the first three lactations of Chinese Holstein cows. J. Integr. Agric. 2015, 14, 125–130. [Google Scholar] [CrossRef]
- Haile-Mariam, M.; Pryce, J.E. Genetic parameters for lactose and its correlation with other milk production traits and fitness traits in pasture-based production systems. J. Dairy Sci. 2017, 100, 3754–3766. [Google Scholar] [CrossRef] [PubMed]
- Miglior, F.; Sewalem, A.; Jamrozik, J.; Bohmanova, J.; Lefebvre, D.M.; Moore, R.K. Genetic analysis of milk urea nitrogen and lactose and their relationships with other production traits in Canadian Holstein cattle. J. Dairy Sci. 2007, 90, 2468–2479. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.K.; Sachdeva, G.K.; Yadav, A.K.; Gautam, S.; Ali, M.M.; Bindal, S. Effect of genetic and non-genetic factors on milk yield and milk constituents of Sahiwal cattle. Indian J. Anim. Res. 2015, 6, 44–47. [Google Scholar]
- Weller, J.I.; Ezra, E. Genetic and phenotypic analysis of daily Israeli Holstein milk, fat, and protein production as determined by a real-time milk analyzer. J. Dairy Sci. 2016, 99, 9782–9795. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.; Schenkel, F.S.; Malchiodi, F.; Ali, R.A.; Mallard, B.; Sargolzaei, M.; Jamrozik, J.; Johnston, J.; Miglior, F. Genetic correlations of mid-infrared-predicted milk fatty acid groups with milkproduction traits. J. Dairy Sci. 2018, 101, 4295–4306. [Google Scholar] [CrossRef] [PubMed]
- Buitenhuis, B.; Poulsen, N.A.; Gebreyesus, G.; Larsen, L.B. Estimation of genetic parameters and detection of chromosomal regions affecting the major milk proteins and their post translational modifications in Danish Holstein and Danish Jersey cattle. BMC Genet. 2016, 17, 114. [Google Scholar] [CrossRef]
- Costa, A.; Lopez-Villalobos, N.; Visentin, G.; De Marchi, M.; Cassandro, M.; Penasa, M. Heritability and repeatability of milk lactose and its relationships with traditional milk traits, somatic cell score and freezing point in Holstein cows. Animal 2019, 13, 909–916. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, A.; Li, X.; Xu, W.; Shi, R.; Luo, H.; Su, G.; Dong, G.; Guo, G.; Wang, Y. Genetic analysis of skinfold thickness and its association with body condition score and milk production traits in Chinese Holstein population. J. Dairy Sci. 2019, 102, 2347–2352. [Google Scholar] [CrossRef]
- Sneddon, N.W.; Lopez-Villalobos, N.; Davis, S.R.; Hickson, R.E.; Shalloo, L. Genetic parameters for milk components including lactose from test day records in the New Zealand dairy herd. New Zeal. J. Agric. Res. 2015, 58, 97–107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Traits | Records | Mean | SD | CV | Minimum | Maximum |
|---|---|---|---|---|---|---|
| PP | 9893 | 3.45 | 0.44 | 0.13 | 2.06 | 4.92 |
| FP | 9871 | 4.19 | 1.13 | 0.27 | 0.70 | 7.84 |
| LP | 9804 | 4.91 | 0.26 | 0.05 | 3.94 | 5.62 |
| Season of Calving | PP | FP | LP | |||
|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | n | Mean ± SE | |
| Spring | 2029 | 3.37 ± 0.01 b,c | 2024 | 3.97 ± 0.03 a | 2017 | 4.93 ± 0.01 a,b |
| Summer | 2198 | 3.34 ± 0.01 c | 2183 | 4.01 ± 0.03 a | 2178 | 4.95 ± 0.01 a |
| Autumn | 2405 | 3.41 ± 0.01 a,b | 2404 | 4.04 ± 0.03 a | 2375 | 4.93 ± 0.01 b |
| Winter | 3261 | 3.43 ± 0.01 a | 3260 | 4.06 ± 0.03 a | 3234 | 4.92 ± 0.01 b |
| Lactation Stage | PP | FP | LP | |||
|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | n | Mean ± SE | |
| 1–50 days | 1204 | 3.20 ± 0.01 d | 1194 | 4.06 ± 0.04 b | 1203 | 4.96 ± 0.01 b |
| 51–100 days | 1018 | 3.17 ± 0.02 d | 1010 | 3.77 ± 0.04 d | 1012 | 5.01 ± 0.01 a |
| 101–200 days | 1613 | 3.36 ± 0.01 c | 1608 | 3.88 ± 0.03 c | 1606 | 4.96 ± 0.01 b |
| 201–305 days | 3500 | 3.52 ± 0.01 b | 3487 | 4.11 ± 0.03 b | 3473 | 4.90 ± 0.01 c |
| >305 days | 2558 | 3.68 ± 0.01 a | 2572 | 4.28 ± 0.03 a | 2510 | 4.83 ± 0.01 d |
| h2 | Spectral Points Number | Heritability | |||
|---|---|---|---|---|---|
| Average | SD | Minimum | Maximum | ||
| <0.01 | 167 | 0.00046 | 0.0015 | 0 | 0.009 |
| 0.01–0.04 | 395 | 0.026 | 0.0045 | 0.01 | 0.04 |
| 0.04–0.07 | 328 | 0.055 | 0.0073 | 0.04 | 0.07 |
| 0.07–0.1 | 132 | 0.083 | 0.0084 | 0.07 | 0.1 |
| >0.1 | 38 | 0.107 | 0.0037 | 0.1 | 0.11 |
| All | 1060 | 0.041 | 0.029 | 0 | 0.11 |
| MIR Region | Heritability | ||||
|---|---|---|---|---|---|
| Name | Range (cm−1) | Average | SD | Minimum | Maximum |
| Lactose | 925–1200 | 0.06 | 0.01 | 0.04 | 0.08 |
| Protein | 1240–1600 | 0.05 | 0.02 | 0.03 | 0.09 |
| Fat-I | 1680–1770 | 0.04 | 0.01 | 0.02 | 0.06 |
| Fat-II | 2800–3015 | 0.06 | 0.02 | 0.04 | 0.10 |
| Total | 925–5011 | 0.04 | 0.03 | 0.00 | 0.11 |
| Traits | MIR Regions (cm−1) | Negative Correlation | Positive Correlation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Average | SD | Min | Max | Average | SD | Min | Max | ||
| PP | 925–1200 (lactose-region) | −0.43 | 0.24 | −0.02 | −0.70 | 0.40 | 0.22 | 0.01 | 0.71 |
| 1240–1600 (protein-region) | −0.22 | 0.14 | −0.06 | −0.42 | 0.47 | 0.32 | 0.01 | 0.94 | |
| 1680–1770 (fat-region) | −0.57 | 0.23 | −0.10 | −0.96 | 0.50 | 0.09 | 0.32 | 0.57 | |
| 2800–3015 (fat-region) | −0.42 | 0.12 | −0.17 | −0.53 | 0.62 | 0.15 | 0.11 | 0.80 | |
| FP | 925–1200 (lactose-region) | −0.60 | 0.28 | −0.02 | −0.85 | 0.43 | 0.28 | 0.02 | 0.78 |
| 1240–1600 (protein-region) | −0.28 | 0.25 | −0.02 | −0.71 | 0.32 | 0.22 | 0.02 | 0.76 | |
| 1680–1770 (fat-region) | −0.67 | 0.21 | −0.07 | −0.88 | 0.74 | 0.04 | 0.63 | 0.78 | |
| 2800–3015 (fat-region) | −0.54 | 0.19 | −0.11 | −0.69 | 0.65 | 0.22 | 0.10 | 0.80 | |
| LP | 925–1200 (lactose-region) | −0.01 | 0 | −0.01 | −0.02 | 0.44 | 0.23 | 0.01 | 0.73 |
| 1240–1600 (protein-region) | −0.17 | 0.09 | −0.03 | −0.26 | 0.44 | 0.21 | 0.01 | 0.70 | |
| 1680–1770 (fat-region) | −0.06 | 0.05 | −0.01 | −0.15 | 0.13 | 0.27 | 0.01 | 0.67 | |
| 2800–3015 (fat-region) | / | / | / | / | 0.21 | 0.12 | 0.06 | 0.47 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, C.; Nan, L.; Yan, L.; Bu, Q.; Ren, X.; Zhang, Z.; Sabek, A.; Zhang, S. Genetic Analysis of Milk Production Traits and Mid-Infrared Spectra in Chinese Holstein Population. Animals 2020, 10, 139. https://doi.org/10.3390/ani10010139
Du C, Nan L, Yan L, Bu Q, Ren X, Zhang Z, Sabek A, Zhang S. Genetic Analysis of Milk Production Traits and Mid-Infrared Spectra in Chinese Holstein Population. Animals. 2020; 10(1):139. https://doi.org/10.3390/ani10010139
Chicago/Turabian StyleDu, Chao, Liangkang Nan, Lei Yan, Qiuyue Bu, Xiaoli Ren, Zhen Zhang, Ahmed Sabek, and Shujun Zhang. 2020. "Genetic Analysis of Milk Production Traits and Mid-Infrared Spectra in Chinese Holstein Population" Animals 10, no. 1: 139. https://doi.org/10.3390/ani10010139
APA StyleDu, C., Nan, L., Yan, L., Bu, Q., Ren, X., Zhang, Z., Sabek, A., & Zhang, S. (2020). Genetic Analysis of Milk Production Traits and Mid-Infrared Spectra in Chinese Holstein Population. Animals, 10(1), 139. https://doi.org/10.3390/ani10010139