
Surface Properties of Parabacteroides distasonis and Impacts of Stress-Induced Molecules on Its Surface Adhesion and Biofilm Formation Capacities

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Method
2.1. Bacterial Strains and Culture Conditions
2.2. Sample Purity and Bacterial Enumeration by Quantitative Polymerase Chain Reaction (qPCR)
2.3. Survivability of P. distasonis under Experimental Conditions
2.4. Homo-Aggregation Assays
2.5. Abiotic Support and Microscopic Observation of the Resulting Microbial Organization
2.5.1. Microtiter Plate for Adhesion and Biofilm Formation Capacity
2.5.2. Microscopic Observation
2.6. Impact of Stress-Induced Molecules on Bacterial Adhesion and Biofilm Formation
2.7. Electrophoretic Mobility Measurements
2.8. Statistical Analysis
3. Results
3.1. Survival Capacity of P. distasonis during Stressing Conditions Linked to Experiments: Aerobic Condition, Saline Solution, Diluted MH and NaNO3 electrolyte
3.2. Homo-Aggregation Capacity of P. Distasonis
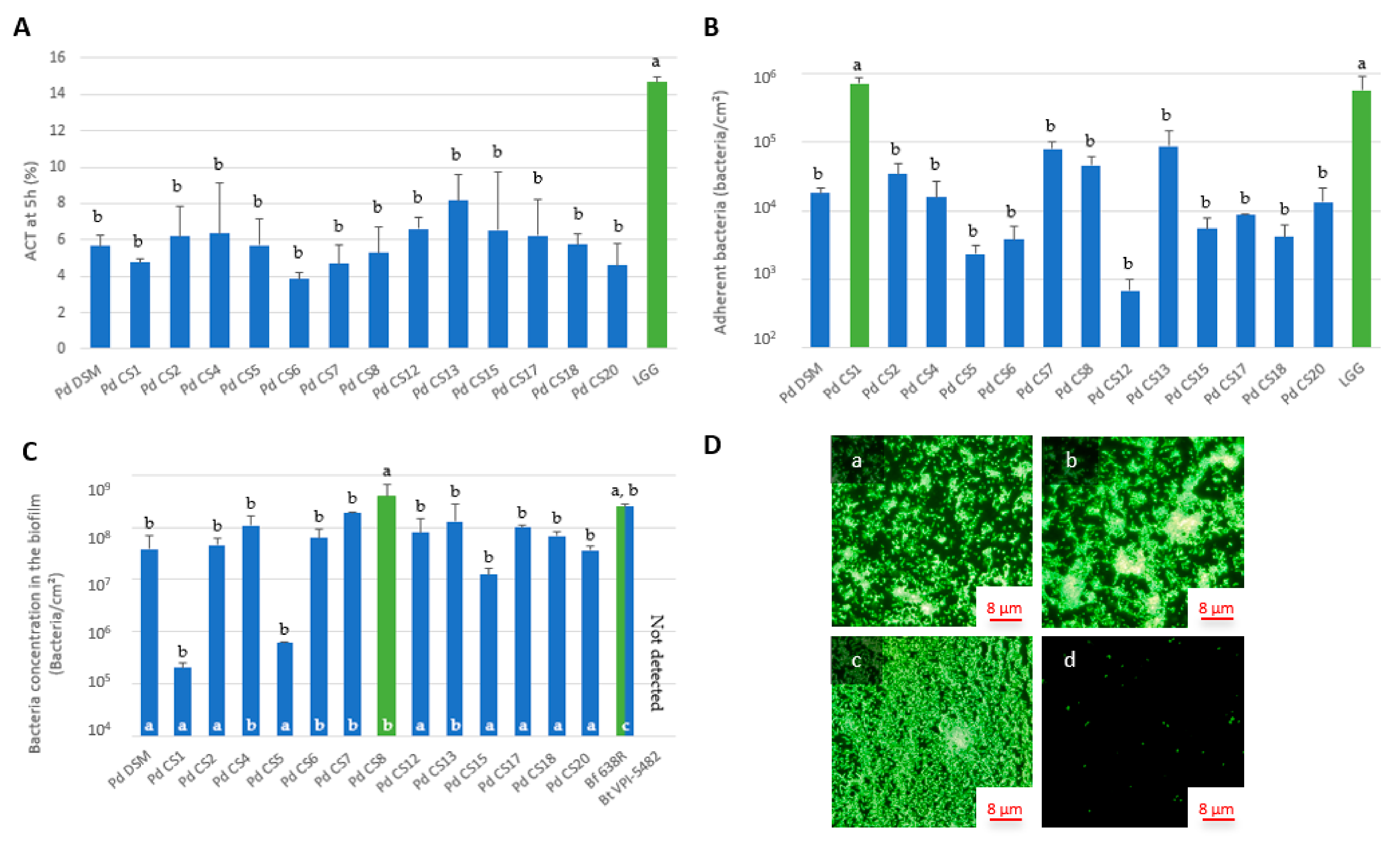
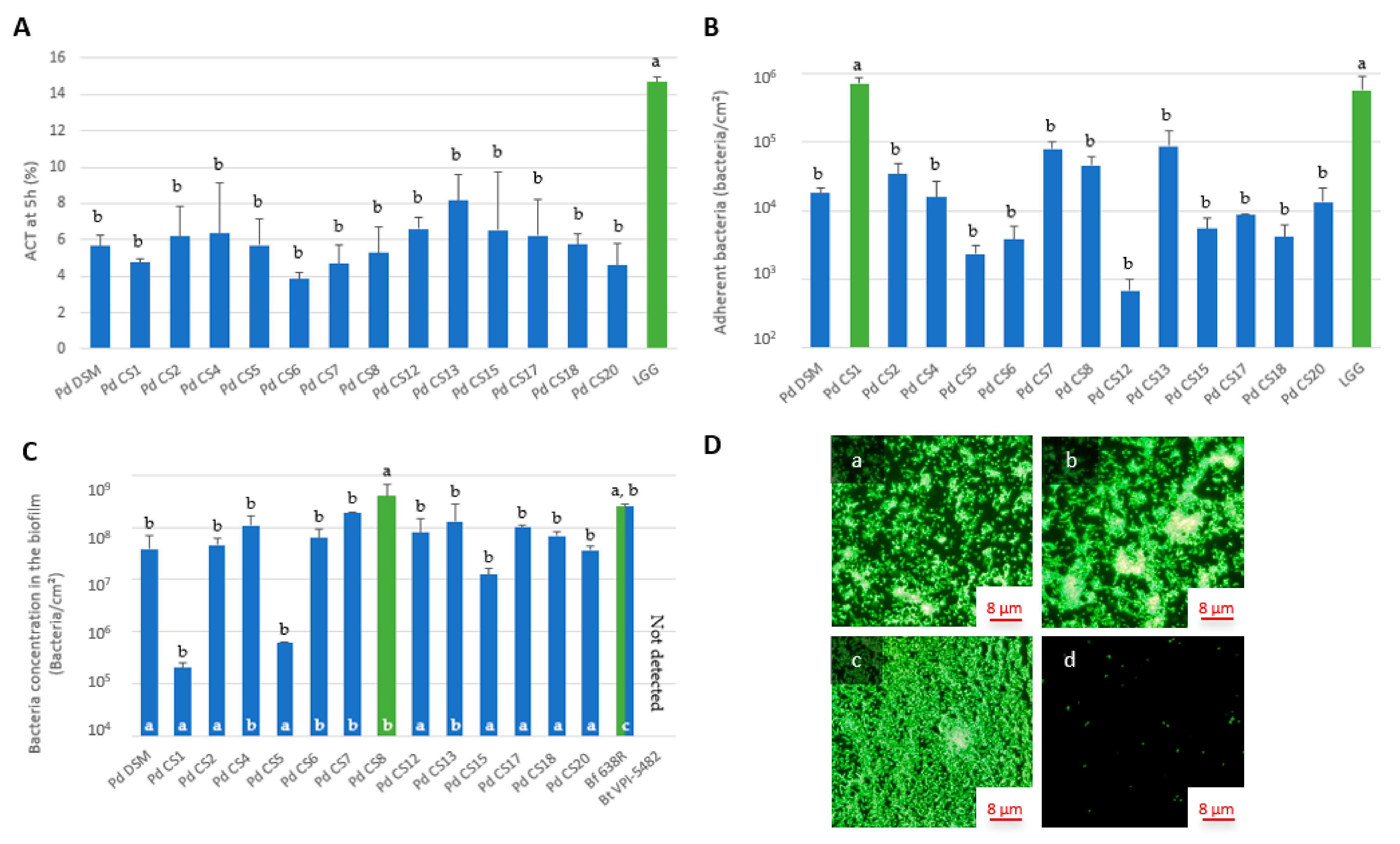
3.3. Adhesion and Biofilm Formation Capacities of P. distasonis Strains on Abiotic Support
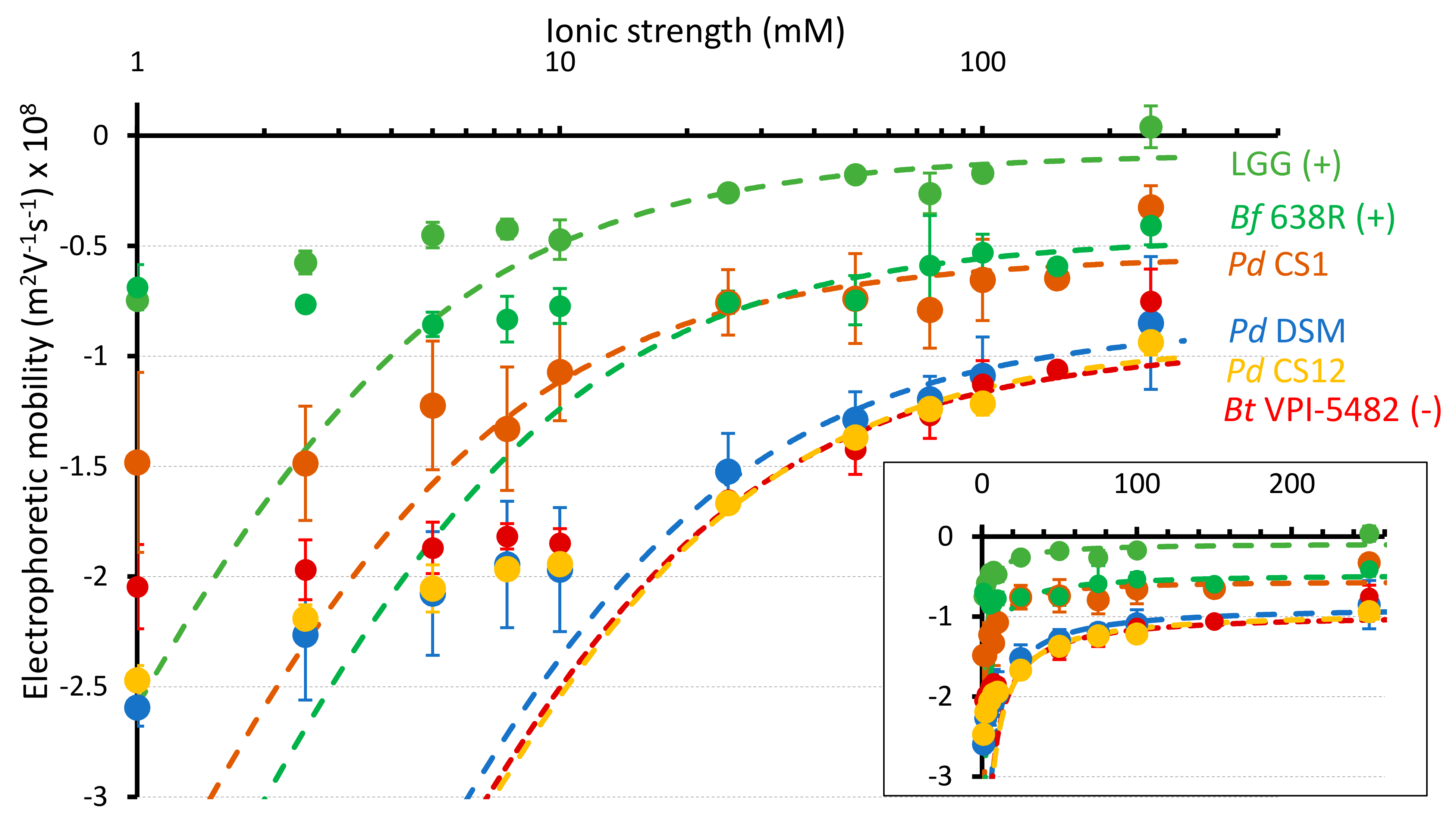
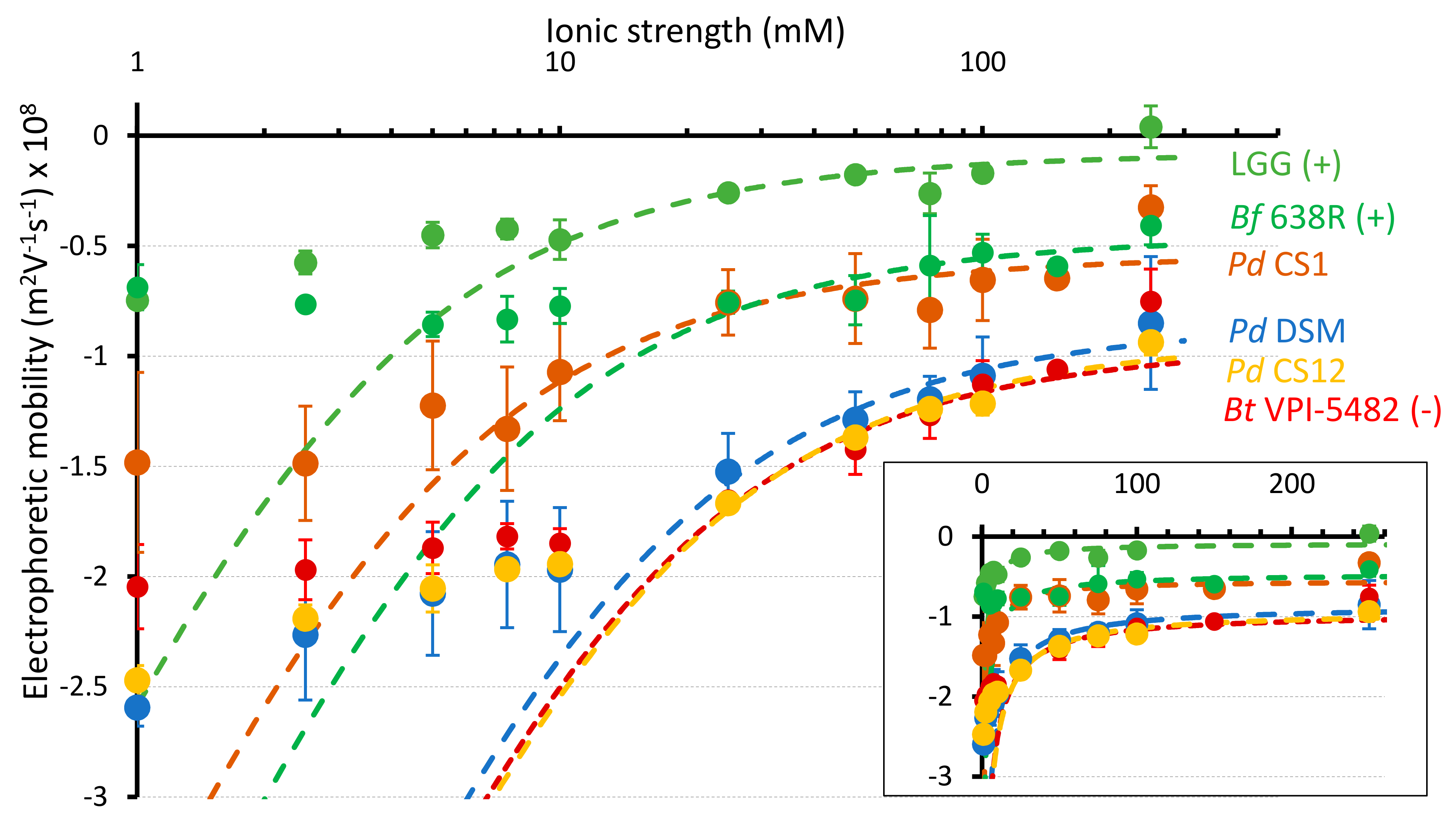
3.4. Electrokinetic Properties of P. distasonis
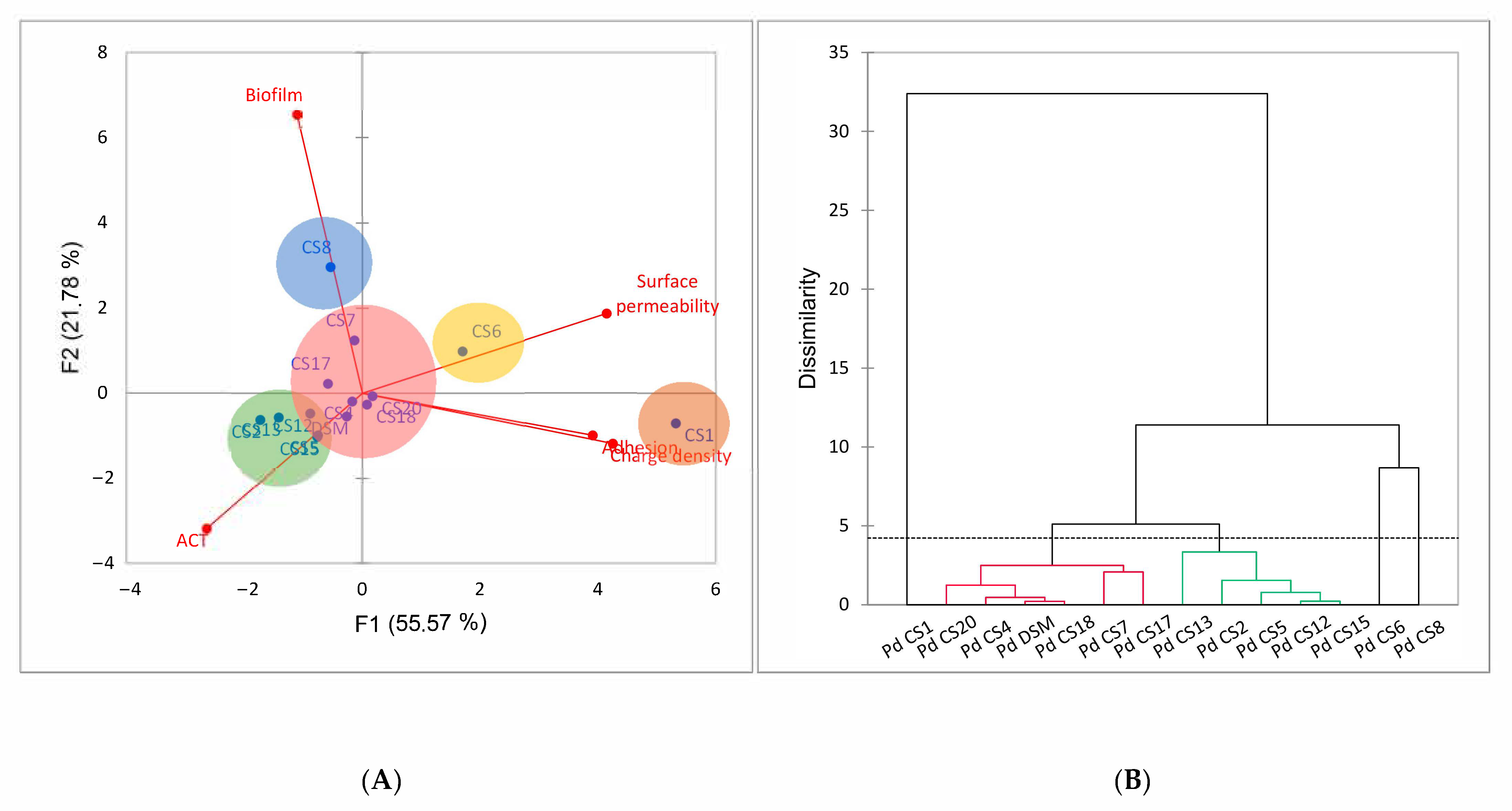
3.5. Correlation between ACT, Electrokinetic Surface, Adhesion and Biofilm Formation Properties of P. distasonis
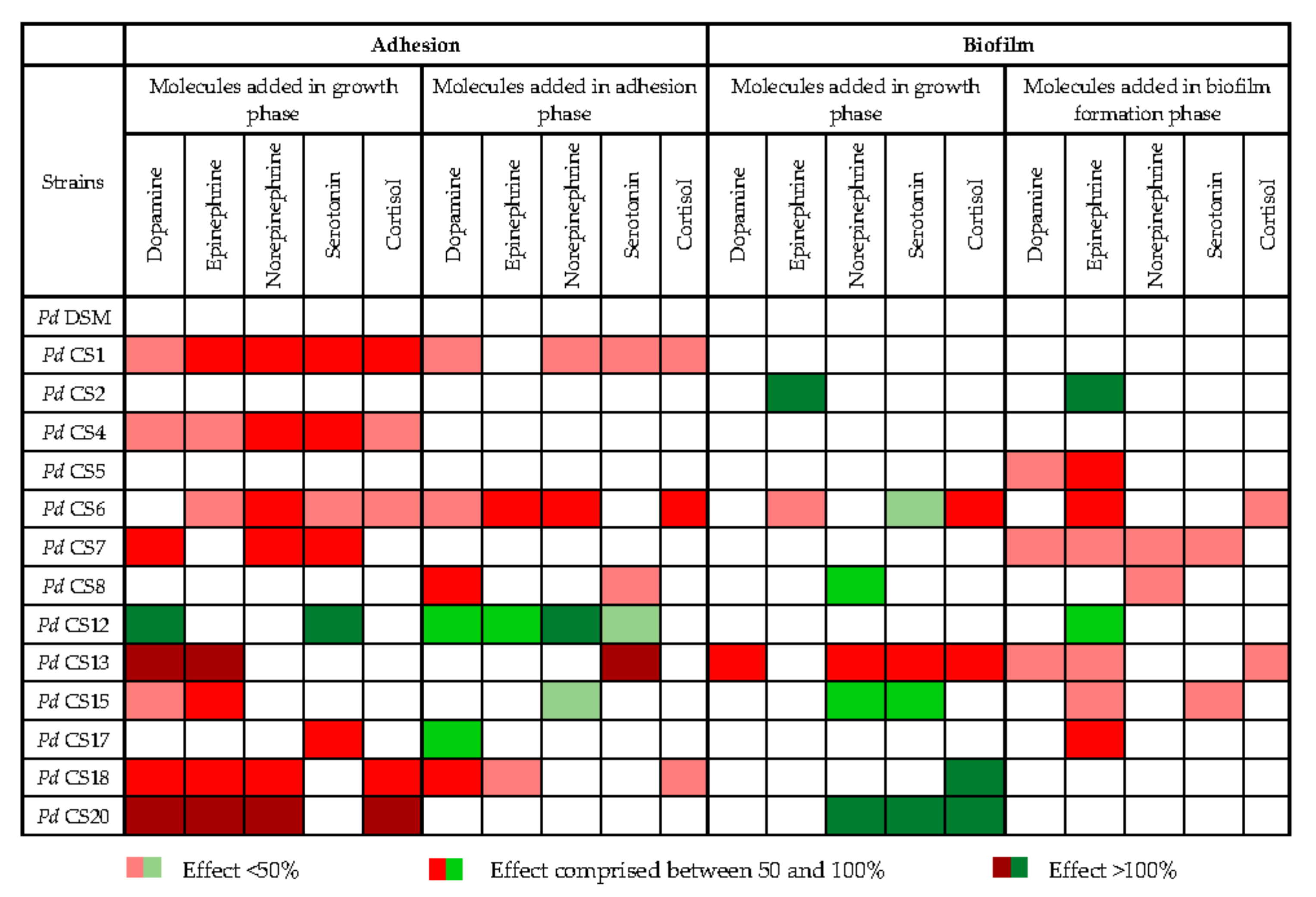
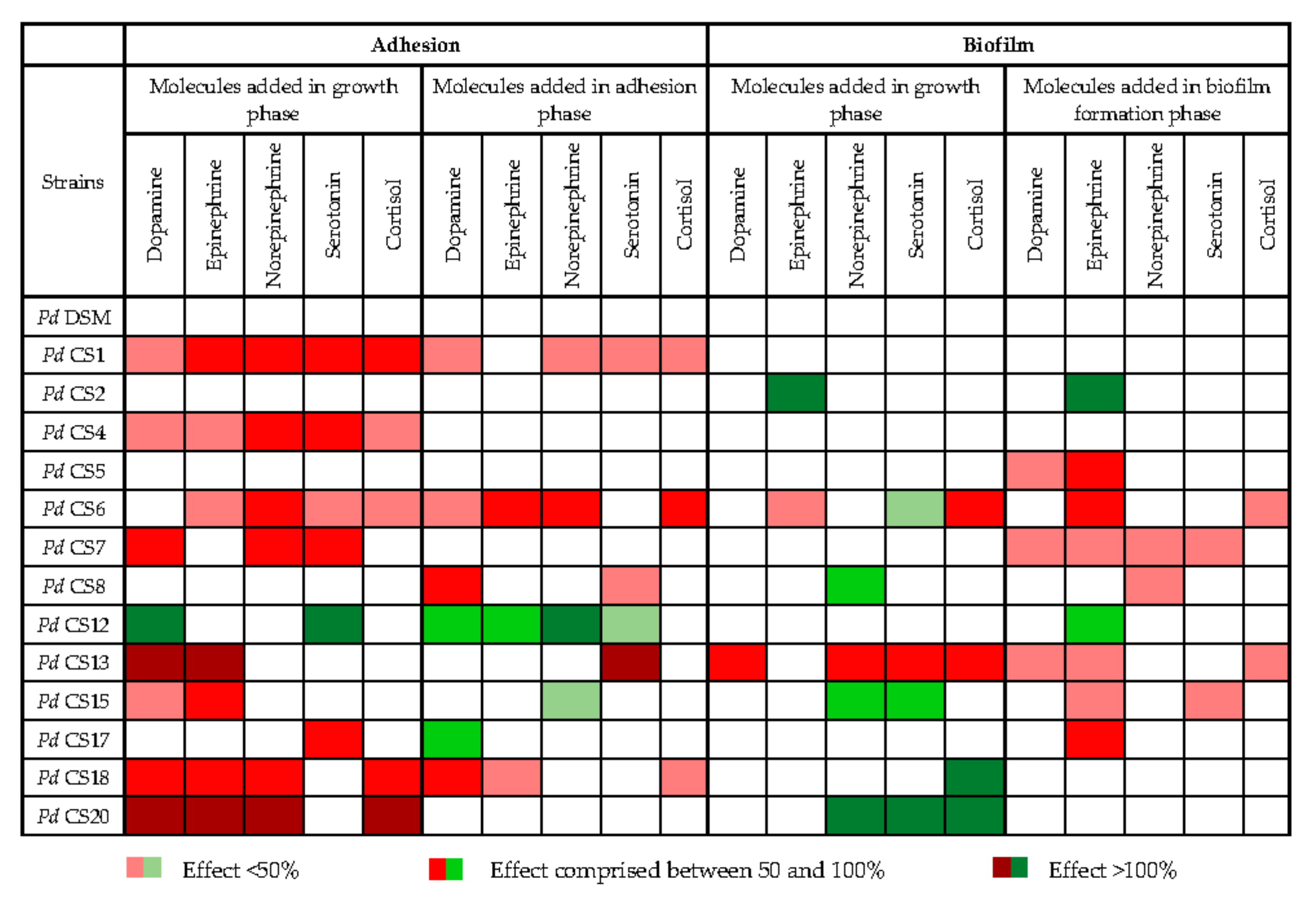
3.6. Impact of Stress-Induced Molecules on Adhesion and Biofilm Formation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Hara, A.M.; Shanahan, F. The Gut Flora as a Forgotten Organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 46. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a Social Stressor Alters the Structure of the Intestinal Microbiota: Implications for Stressor-Induced Immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut Microbiota: Role in Pathogen Colonization, Immune Responses and Inflammatory Disease. Immunol. Rev. 2018, 39, 70–89. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-Based Metagenomics Analysis Reveals Markers for Gut Microbiome Composition and Diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Galley, J.D.; Parry, N.M.; Ahmer, B.M.M.; Fox, J.G.; Bailey, M.T. The Commensal Microbiota Exacerbate Infectious Colitis in Stressor-Exposed Mice. Brain Behav. Immun. 2017, 60, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siopi, E.; Chevalier, G.; Katsimpardi, L.; Saha, S.; Bigot, M.; Moigneu, C.; Eberl, G.; Lledo, P.-M. Changes in Gut Microbiota by Chronic Stress Impair the Efficacy of Fluoxetine. Cell Rep. 2020, 30, 3682–3690.e6. [Google Scholar] [CrossRef] [PubMed]
- Alauzet, C.; Cunat, L.; Wack, M.; Lozniewski, A.; Busby, H.; Agrinier, N.; Cailliez-Grimal, C.; Frippiat, J.-P. Hypergravity Disrupts Murine Intestinal Microbiota. Sci. Rep. 2019, 9, 9410. [Google Scholar] [CrossRef] [PubMed]
- Alauzet, C.; Cunat, L.; Wack, M.; Lanfumey, L.; Legrand-Frossi, C.; Lozniewski, A.; Agrinier, N.; Cailliez-Grimal, C.; Frippiat, J.-P. Impact of a Model Used to Simulate Chronic Socio-Environmental Stressors Encountered during Spaceflight on Murine Intestinal Microbiota. Int. J. Mol. Sci. 2020, 21, 7863. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-Level Analysis of Gut Microbiome Variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Gutiérrez-Repiso, C.; Molina-Vega, M.; Bernal-López, M.R.; Garrido-Sánchez, L.; García-Almeida, J.M.; Sajoux, I.; Moreno-Indias, I.; Tinahones, F.J. Different Weight Loss Intervention Approaches Reveal a Lack of a Common Pattern of Gut Microbiota Changes. Pers. Med. 2021, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Mahowald, M.A.; Ley, R.E.; Lozupone, C.A.; Hamady, M.; Martens, E.C.; Henrissat, B.; Coutinho, P.M.; Minx, P.; Latreille, P.; et al. Evolution of Symbiotic Bacteria in the Distal Human Intestine. PLoS Biol. 2007, 5, e156. [Google Scholar] [CrossRef]
- Wang, X.; Conway, P.L.; Brown, I.L.; Evans, A.J. In Vitro Utilization of Amylopectin and High-Amylose Maize (Amylomaize) Starch Granules by Human Colonic Bacteria. Appl. Environ. Microbiol. 1999, 65, 4848–4854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiippala, K.; Kainulainen, V.; Suutarinen, M.; Heini, T.; Bowers, J.R.; Jasso-Selles, D.; Lemmer, D.; Valentine, M.; Barnes, R.; Engelthaler, D.M.; et al. Isolation of Anti-Inflammatory and Epithelium Reinforcing Bacteroides and Parabacteroides spp. from A Healthy Fecal Donor. Nutrients 2020, 12, 935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, G.Y.; Kane, A.; Lee, K.; Xu, Q.; Wu, X.; Roper, J.; Mason, J.B.; Crott, J.W. Parabacteroides distasonis Attenuates Toll-like Receptor 4 Signaling and Akt Activation and Blocks Colon Tumor Formation in High-fat Diet-fed Azoxymethane-treated Mice. Int. J. Cancer 2018, 143, 1797–1805. [Google Scholar] [CrossRef] [Green Version]
- Kverka, M.; Zakostelska, Z.; Klimesova, K.; Sokol, D.; Hudcovic, T.; Hrncir, T.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Verdu, E.F.; et al. Oral Administration of Parabacteroides distasonis Antigens Attenuates Experimental Murine Colitis through Modulation of Immunity and Microbiota Composition: Bacterial Components and Experimental Colitis. Clin. Exp. Immunol. 2011, 163, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Súkeníková, L.; Desramaut, J.; Boudebbouze, S.; Salomé-Desnoulez, S.; Hrdý, J.; Waligora-Dupriet, A.-J.; Maguin, E.; et al. In Vitro Characterization of Gut Microbiota-Derived Commensal Strains: Selection of Parabacteroides distasonis Strains Alleviating TNBS-Induced Colitis in Mice. Cells 2020, 9, 2104. [Google Scholar] [CrossRef]
- Koh, G.Y.; Kane, A.V.; Wu, X.; Crott, J.W. Parabacteroides distasonis Attenuates Tumorigenesis, Modulates Inflammatory Markers and Promotes Intestinal Barrier Integrity in Azoxymethane-Treated A/J Mice. Carcinogenesis 2020, 41, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Peucelle, V.; Desramaut, J.; Boudebbouze, S.; Croyal, M.; Waligora-Dupriet, A.-J.; Rhimi, M.; Grangette, C.; et al. Identification of New Potential Biotherapeutics from Human Gut Microbiota-Derived Bacteria. Microorganisms 2021, 9, 565. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbial. 1995, 35, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J. Introduction to Biofilm. Int. J. Antimicrob. Agents 1999, 11, 217–221. [Google Scholar] [CrossRef]
- Nikolaev, Y.A.; Plakunov, V.K. Biofilm—“City of Microbes” or an Analogue of Multicellular Organisms? Microbiology 2007, 76, 125–138. [Google Scholar] [CrossRef]
- Roberts, A.; Matthews, J.B.; Socransky, S.S.; Freestone, P.P.E.; Williams, P.H.; Chapple, I.L.C. Stress and the Periodontal Diseases: Effects of Catecholamines on the Growth of Periodontal Bacteria In Vitro: Effects of Catecholamines on Periodontal Bacteria. Oral Microbiol. Immunol. 2002, 17, 296–303. [Google Scholar] [CrossRef]
- Freestone, P.P.E.; Lyte, M. Chapter 2 Microbial Endocrinology: Experimental Design Issues in the Study of Interkingdom Signalling in Infectious Disease. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2008; Volume 64, pp. 75–105. ISBN 978-0-12-374338-1. [Google Scholar]
- Knecht, L.D.; O’Connor, G.; Mittal, R.; Liu, X.Z.; Daftarian, P.; Deo, S.K.; Daunert, S. Serotonin Activates Bacterial Quorum Sensing and Enhances the Virulence of Pseudomonas aeruginosa in the Host. EBioMedicine 2016, 9, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Torabi Delshad, S.; Soltanian, S.; Sharifiyazdi, H.; Bossier, P. Effect of Catecholamine Stress Hormones (Dopamine and Norepinephrine) on Growth, Swimming Motility, Biofilm Formation and Virulence Factors of Yersinia ruckeri in Vitro and an in Vivo Evaluation in Rainbow Trout. J. Fish Dis. 2019, 42, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Declercq, A.M.; Cai, W.; Naranjo, E.; Thongda, W.; Eeckhaut, V.; Bauwens, E.; Arias, C.; De La Fuente, L.; Beck, B.H.; Lange, M.D.; et al. Evidence That the Stress Hormone Cortisol Regulates Biofilm Formation Differently among Flavobacterium columnare Isolates. Vet. Res. 2019, 50, 24. [Google Scholar] [CrossRef] [Green Version]
- Hiller, C.C.; Lucca, V.; Carvalho, D.; Borsoi, A.; Borges, K.A.; Furian, T.Q.; do Nascimento, V.P. Influence of Catecholamines on Biofilm Formation by Salmonella enteritidis. Microb. Pathog. 2019, 130, 54–58. [Google Scholar] [CrossRef]
- Francius, G.; Polyakov, P.; Merlin, J.; Abe, Y.; Ghigo, J.-M.; Merlin, C.; Beloin, C.; Duval, J.F.L. Bacterial Surface Appendages Strongly Impact Nanomechanical and Electrokinetic Properties of Escherichia coli Cells Subjected to Osmotic Stress. PLoS ONE 2011, 6, e20066. [Google Scholar] [CrossRef]
- Gosselin, F.; Duval, J.F.L.; Simonet, J.; Ginevra, C.; Gaboriaud, F.; Jarraud, S.; Mathieu, L. Impact of the Virulence-Associated MAb3/1 Epitope on the Physicochemical Surface Properties of Legionella pneumophila Sg1: An Issue to Explain Infection Potential? Colloids Surf. B Biointerfaces 2011, 82, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Hyono, A.; Gaboriaud, F.; Mazda, T.; Takata, Y.; Ohshima, H.; Duval, J.F.L. Impacts of Papain and Neuraminidase Enzyme Treatment on Electrohydrodynamics and IgG-Mediated Agglutination of Type A Red Blood Cells. Langmuir 2009, 25, 10873–10885. [Google Scholar] [CrossRef]
- Pagnout, C.; Sohm, B.; Razafitianamaharavo, A.; Caillet, C.; Offroy, M.; Leduc, M.; Gendre, H.; Jomini, S.; Beaussart, A.; Bauda, P.; et al. Pleiotropic Effects of Rfa-Gene Mutations on Escherichia coli Envelope Properties. Sci. Rep. 2019, 9, 9696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquot, A.; Sakamoto, C.; Razafitianamarahavo, A.; Caillet, C.; Merlin, J.; Fahs, A.; Ghigo, J.-M.; Duval, J.F.L.; Beloin, C.; Francius, G. The Dynamics and PH-Dependence of Ag43 Adhesins’ Self-Association Probed by Atomic Force Spectroscopy. Nanoscale 2014, 6, 12665–12681. [Google Scholar] [CrossRef] [Green Version]
- Van Belkum, A.; Kluytmans, J.; van Leeuwen, W.; Bax, R.; Quint, W.; Peters, E.; Fluit, A.; Vandenbroucke-Grauls, C.; van den Brule, A.; Koeleman, H. Multicenter Evaluation of Arbitrarily Primed PCR for Typing of Staphylococcus aureus Strains. J. Clin. Microbiol. 1995, 33, 1537–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of Repetitive DNA Sequences in Eubacteria and Application to Fingerprinting of Bacterial Genomes. Nucleic Acids Res. 1991, 9, 6823–6831. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Waśko, A.; Paduch, R.; Skrzypek, T.; Sroka-Bartnicka, A. The Effect of Cell Surface Components on Adhesion Ability of Lactobacillus rhamnosus. Antonie van Leeuwenhoek 2014, 106, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sá Almeida, J.S.; de Oliveira Marre, A.T.; Teixeira, F.L.; Boente, R.F.; Domingues, R.M.C.P.; de Paula, G.R.; Lobo, L.A. Lactoferrin and Lactoferricin B Reduce Adhesion and Biofilm Formation in the Intestinal Symbionts Bacteroides fragilis and Bacteroides thetaiotaomicron. Anaerobe 2020, 64, 102232. [Google Scholar] [CrossRef]
- Mihajlovic, J.; Bechon, N.; Ivanova, C.; Chain, F.; Almeida, A.; Langella, P.; Beloin, C.; Ghigo, J.-M. A Putative Type V Pilus Contributes to Bacteroides thetaiotaomicron Biofilm Formation Capacity. J. Bacteriol. 2019, 201, e00650-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and Aggregation Properties of Probiotic and Pathogen Strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- Tasse, J.; Cara, A.; Saglio, M.; Villet, R.; Laurent, F. A Steam-Based Method to Investigate Biofilm. Sci. Rep. 2018, 8, 13040. [Google Scholar] [CrossRef] [PubMed]
- Donelli, G.; Vuotto, C.; Cardines, R.; Mastrantonio, P. Biofilm-Growing Intestinal Anaerobic Bacteria. FEMS Immunol. Med. Microbiol. 2012, 65, 318–325. [Google Scholar] [CrossRef]
- Duval, J.F.L.; Gaboriaud, F. Progress in Electrohydrodynamics of Soft Microbial Particle Interphases. Curr. Opin. Colloid Interface Sci. 2010, 15, 184–195. [Google Scholar] [CrossRef]
- Ohshima, H. Electrophoresis of Soft Particles. Adv. Colloid Interface Sci. 1995, 47, 189–235. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut Bacteria from Multiple Sclerosis Patients Modulate Human T Cells and Exacerbate Symptoms in Mouse Models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef] [Green Version]
- Zitomersky, N.L.; Atkinson, B.J.; Franklin, S.W.; Mitchell, P.D.; Snapper, S.B.; Comstock, L.E.; Bousvaros, A. Characterization of Adherent Bacteroidales from Intestinal Biopsies of Children and Young Adults with Inflammatory Bowel Disease. PLoS ONE 2013, 8, e63686. [Google Scholar] [CrossRef] [Green Version]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A Microbial Signature for Crohn’s Disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef]
- Pfalzer, A.C.; Nesbeth, P.-D.C.; Parnell, L.D.; Iyer, L.K.; Liu, Z.; Kane, A.V.; Chen, C.-Y.O.; Tai, A.K.; Bowman, T.A.; Obin, M.S.; et al. Diet- and Genetically-Induced Obesity Differentially Affect the Fecal Microbiome and Metabolome in Apc1638N Mice. PLoS ONE 2015, 10, e0135758. [Google Scholar] [CrossRef] [Green Version]
- Pérez, P.F.; Minnaard, Y.; Disalvo, E.A.; De Antoni, G.L. Surface Properties of Bifidobacterial Strains of Human Origin. Appl. Environ. Microbiol. 1998, 64, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thewes, N.; Loskill, P.; Jung, P.; Peisker, H.; Bischoff, M.; Herrmann, M.; Jacobs, K. Hydrophobic Interaction Governs Unspecific Adhesion of Staphylococci: A Single Cell Force Spectroscopy Study. Beilstein J. Nanotechnol. 2014, 5, 1501–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobley, L.; Harkins, C.; MacPhee, C.E.; Stanley-Wall, N.R. Giving Structure to the Biofilm Matrix: An Overview of Individual Strategies and Emerging Common Themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bircher, L.; Schwab, C.; Geirnaert, A.; Greppi, A.; Lacroix, C. Planktonic and Sessile Artificial Colonic Microbiota Harbor Distinct Composition and Reestablish Differently upon Frozen and Freeze-Dried Long-Term Storage. mSystems 2020, 5, e00521-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlanga, M.; Guerrero, R. Living Together in Biofilms: The Microbial Cell Factory and Its Biotechnological Implications. Microb. Cell. Fact. 2016, 15, 165. [Google Scholar] [CrossRef] [Green Version]
- Patrick, S.; Blakely, G.W.; Houston, S.; Moore, J.; Abratt, V.R.; Bertalan, M.; Cerdeño-Tárraga, A.M.; Quail, M.A.; Corton, N.; Corton, C.; et al. Twenty-Eight Divergent Polysaccharide Loci Specifying within- and amongst-Strain Capsule Diversity in Three Strains of Bacteroides fragilis. Microbiology 2010, 156, 3255–3269. [Google Scholar] [CrossRef] [Green Version]
- Pumbwe, L.; Skilbeck, C.A.; Wexler, H.M. The Bacteroides fragilis Cell Envelope: Quarterback, Linebacker, Coach—or All Three? Anaerobe 2006, 12, 211–220. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Nakano, V.; Piazza, R.M.F.; Cianciarullo, A.M.; Bueris, V.; Santos, M.F.; Menezes, M.A.; Mendes-Ledesma, M.R.B.; Szulczewski, V.; Elias, W.P.; Pumbwe, L.; et al. Adherence and Invasion of Bacteroidales Isolated from the Human Intestinal Tract. Clin. Microbiol. Infect. 2008, 14, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Coyne, M.J.; Weinacht, K.G.; Krinos, C.M.; Comstock, L.E. Mpi Recombinase Globally Modulates the Surface Architecture of a Human Commensal Bacterium. Proc. Natl. Acad. Sci. USA 2003, 100, 10446–10451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, C.M.; Coyne, M.J.; Bentley, D.L.; Villa, O.F.; Comstock, L.E. Phase-Variable Expression of a Family of Glycoproteins Imparts a Dynamic Surface to a Symbiont in Its Human Intestinal Ecosystem. Proc. Natl. Acad. Sci. USA 2007, 104, 2413–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konduri, R.; Saiabhilash, C.R.; Shivaji, S. Biofilm-Forming Potential of Ocular Fluid Staphylococcus aureus and Staphylococcus epidermidis on Ex Vivo Human Corneas from Attachment to Dispersal Phase. Microorganisms 2021, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Burtseva, O.; Baulina, O.; Zaytseva, A.; Fedorenko, T.; Chekanov, K.; Lobakova, E. In Vitro Biofilm Formation by Bioluminescent Bacteria Isolated from the Marine Fish Gut. Microb. Ecol. 2021, 81, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Brown, D.R.; Xie, Y.; Green, B.T.; Lyte, M. Catecholamines Modulate Escherichia coli O157:H7 Adherence to Murine Cecal Mucosa. Shock 2003, 20, 183–188. [Google Scholar] [CrossRef]
- Clarke, M.B.; Hughes, D.T.; Zhu, C.; Boedeker, E.C.; Sperandio, V. The QseC Sensor Kinase: A Bacterial Adrenergic Receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 10420–10425. [Google Scholar] [CrossRef] [Green Version]
- Sarkodie, E.K.; Zhou, S.; Baidoo, S.A.; Chu, W. Influences of Stress Hormones on Microbial Infections. Microb. Pathog. 2019, 131, 270–276. [Google Scholar] [CrossRef]
- Gerits, E.; Verstraeten, N.; Michiels, J. New Approaches to Combat Porphyromonas gingivalis Biofilms. J. Oral Microbiol. 2017, 9, 1300366. [Google Scholar] [CrossRef] [Green Version]
- Pumbwe, L.; Skilbeck, C.A.; Wexler, H.M. Presence of Quorum-Sensing Systems Associated with Multidrug Resistance and Biofilm Formation in Bacteroides fragilis. Microb. Ecol. 2008, 56, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Boyanova, L. Stress Hormone Epinephrine (Adrenaline) and Norepinephrine (Noradrenaline) Effects on the Anaerobic Bacteria. Anaerobe 2017, 44, 13–19. [Google Scholar] [CrossRef]
- Saito, T.; Inagaki, S.; Sakurai, K.; Okuda, K.; Ishihara, K. Exposure of P. gingivalis to Noradrenaline Reduces Bacterial Growth and Elevates ArgX Protease Activity. Arch. Oral Biol. 2011, 56, 244–250. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | 1/λo (nm) | |
|---|---|---|
| P. distasonis DSM 20701T | −26.6 | 1.79 |
| P. distasonis CS1 | −8.6 | 2.51 |
| P. distasonis CS2 | −37.0 | 1.60 |
| P. distasonis CS4 | −30.4 | 1.80 |
| P. distasonis CS5 | −23.5 | 1.85 |
| P. distasonis CS6 | −23.7 | 2.23 |
| P. distasonis CS7 | −29.0 | 1.84 |
| P. distasonis CS8 | −32.3 | 1.98 |
| P. distasonis CS12 | −29.1 | 1.78 |
| P. distasonis CS13 | −33.8 | 1.80 |
| P. distasonis CS15 | −29.1 | 1.78 |
| P. distasonis CS17 | −33.8 | 2.04 |
| P. distasonis CS18 | −24.0 | 1.88 |
| P. distasonis CS20 | −27.4 | 1.87 |
| L. rhamnosus GG ATCC 53103 | −7.0 | 1.07 |
| B. fragilis 638R | −12.5 | 1.91 |
| B. thetaiotaomicron VPI-5482 | −27.3 | 1.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamarande, J.; Cunat, L.; Caillet, C.; Mathieu, L.; Duval, J.F.L.; Lozniewski, A.; Frippiat, J.-P.; Alauzet, C.; Cailliez-Grimal, C. Surface Properties of Parabacteroides distasonis and Impacts of Stress-Induced Molecules on Its Surface Adhesion and Biofilm Formation Capacities. Microorganisms 2021, 9, 1602. https://doi.org/10.3390/microorganisms9081602
Chamarande J, Cunat L, Caillet C, Mathieu L, Duval JFL, Lozniewski A, Frippiat J-P, Alauzet C, Cailliez-Grimal C. Surface Properties of Parabacteroides distasonis and Impacts of Stress-Induced Molecules on Its Surface Adhesion and Biofilm Formation Capacities. Microorganisms. 2021; 9(8):1602. https://doi.org/10.3390/microorganisms9081602
Chicago/Turabian StyleChamarande, Jordan, Lisiane Cunat, Céline Caillet, Laurence Mathieu, Jérôme F. L. Duval, Alain Lozniewski, Jean-Pol Frippiat, Corentine Alauzet, and Catherine Cailliez-Grimal. 2021. "Surface Properties of Parabacteroides distasonis and Impacts of Stress-Induced Molecules on Its Surface Adhesion and Biofilm Formation Capacities" Microorganisms 9, no. 8: 1602. https://doi.org/10.3390/microorganisms9081602
APA StyleChamarande, J., Cunat, L., Caillet, C., Mathieu, L., Duval, J. F. L., Lozniewski, A., Frippiat, J.-P., Alauzet, C., & Cailliez-Grimal, C. (2021). Surface Properties of Parabacteroides distasonis and Impacts of Stress-Induced Molecules on Its Surface Adhesion and Biofilm Formation Capacities. Microorganisms, 9(8), 1602. https://doi.org/10.3390/microorganisms9081602