
It Takes Two to Tango: A Bacterial Biofilm Provides Protection against a Fungus-Feeding Bacterial Predator

 , , and
, , and 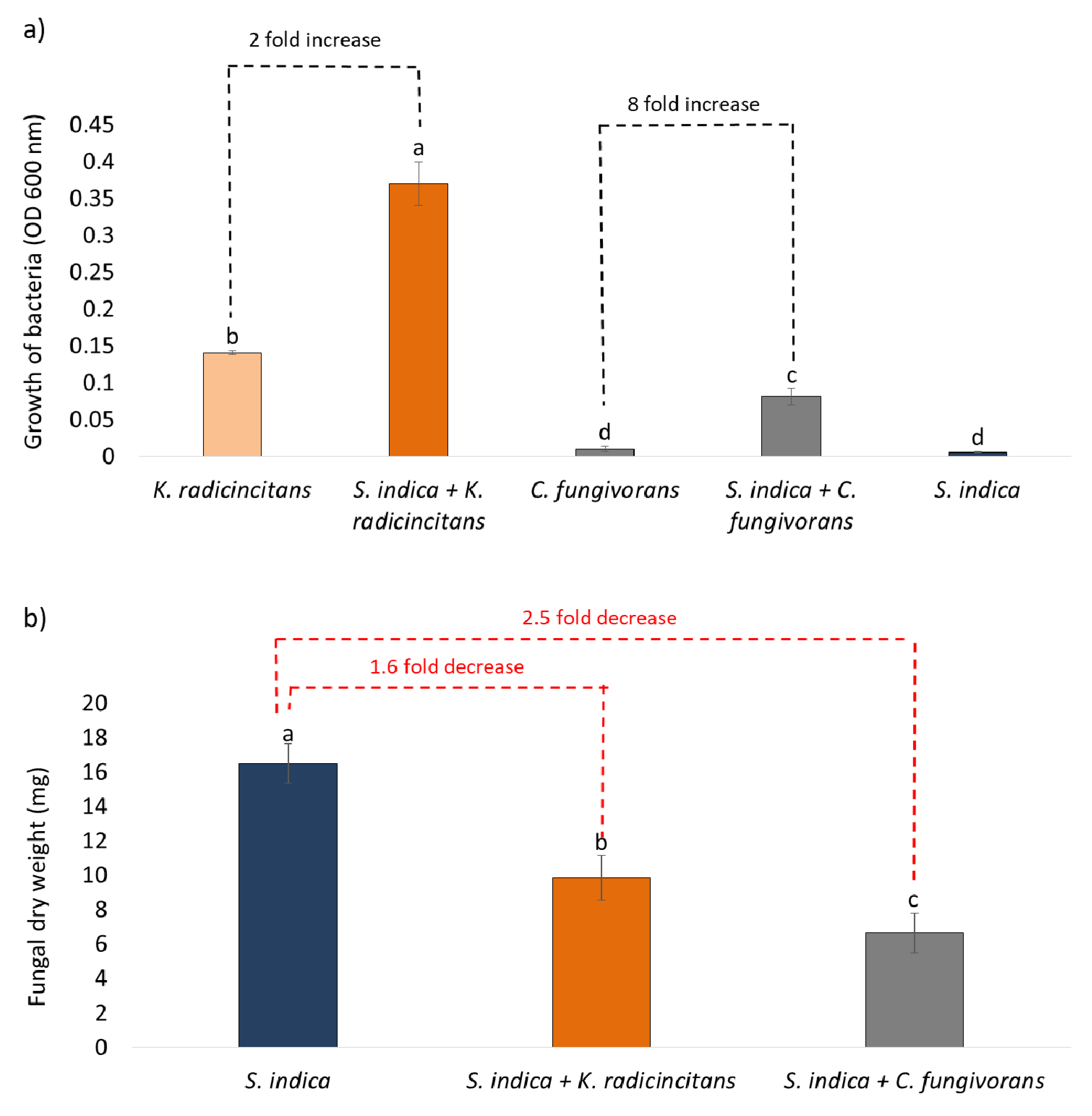{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms Used in This Study
2.2. Cultivation of Microorganisms
2.3. Fungal Growth Inhibition Assay
2.4. Mycophagy Assay
2.5. Extraction of Metabolites of S. indica
2.6. Growth of the Bacteria on Metabolites of S. indica
2.7. Co-Cultivation of Bacteria and Fungus (with or without Physical Contact)
2.8. Tripartite Interaction: K. radicincitans, S. indica and C. fungivorans on Solid Media
2.9. Study of Tripartite Interaction: K. radicincitans, S. indica, and C. fungivorans in Liquid Media
2.10. DNA Extraction and Quantitative PCR (qPCR)
2.11. Confocal Laser Scanning Microscopy (CLSM) Analysis
2.12. Data Analysis
3. Results
3.1. Fungal Growth Inhibition Assays
3.2. Mycophagy Assay
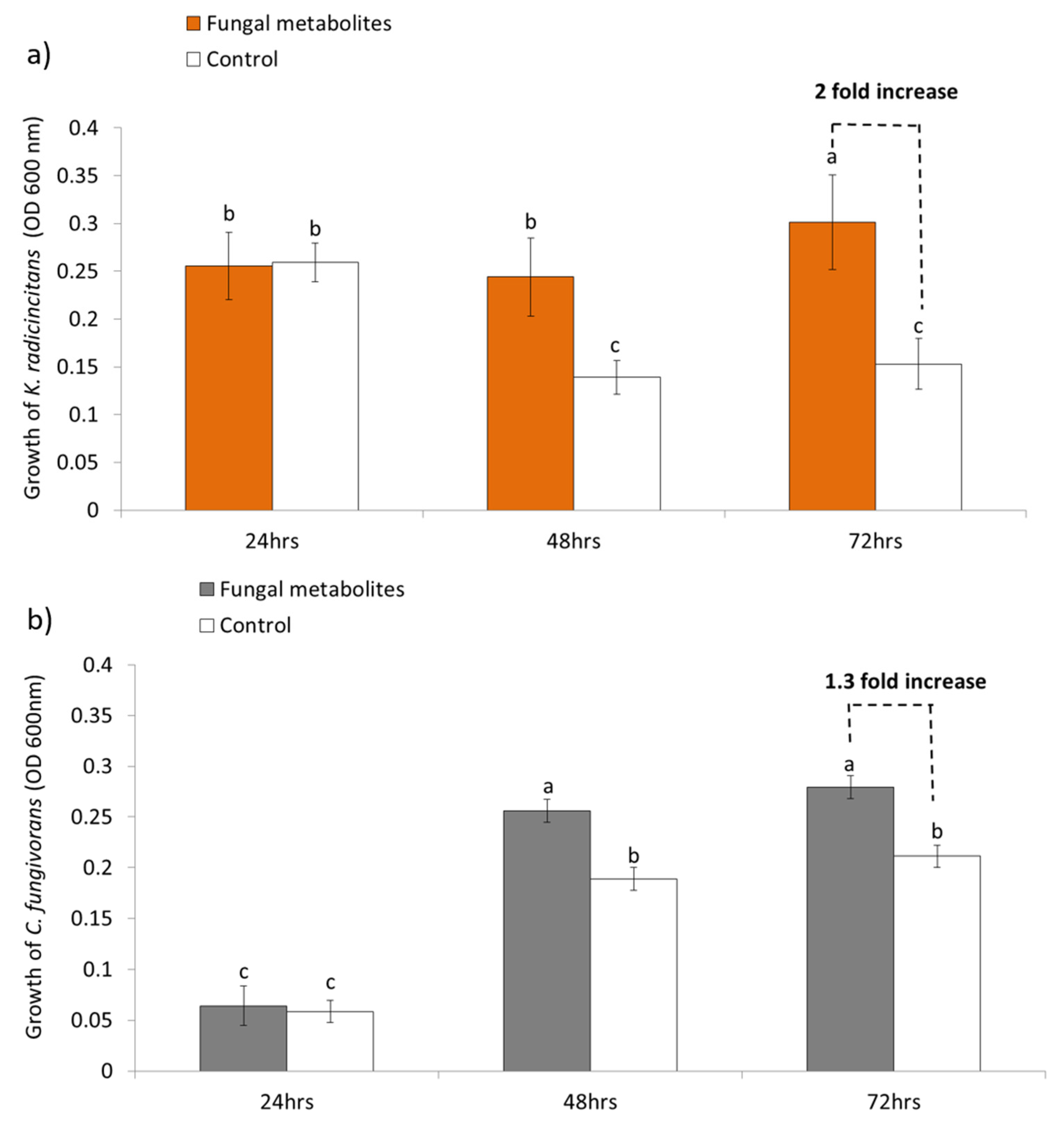
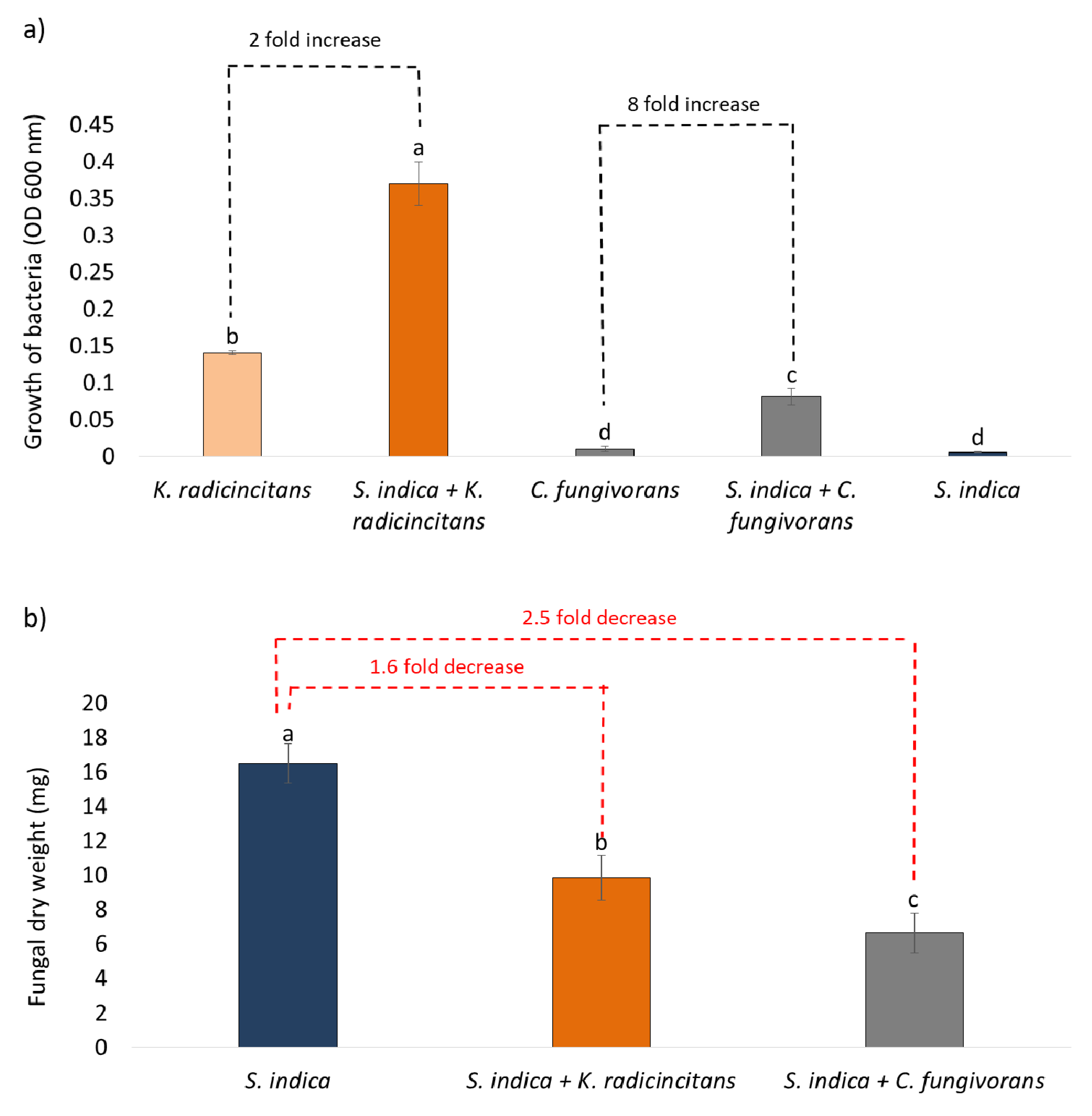
3.3. Growth of the Bacteria on Fungal Metabolites
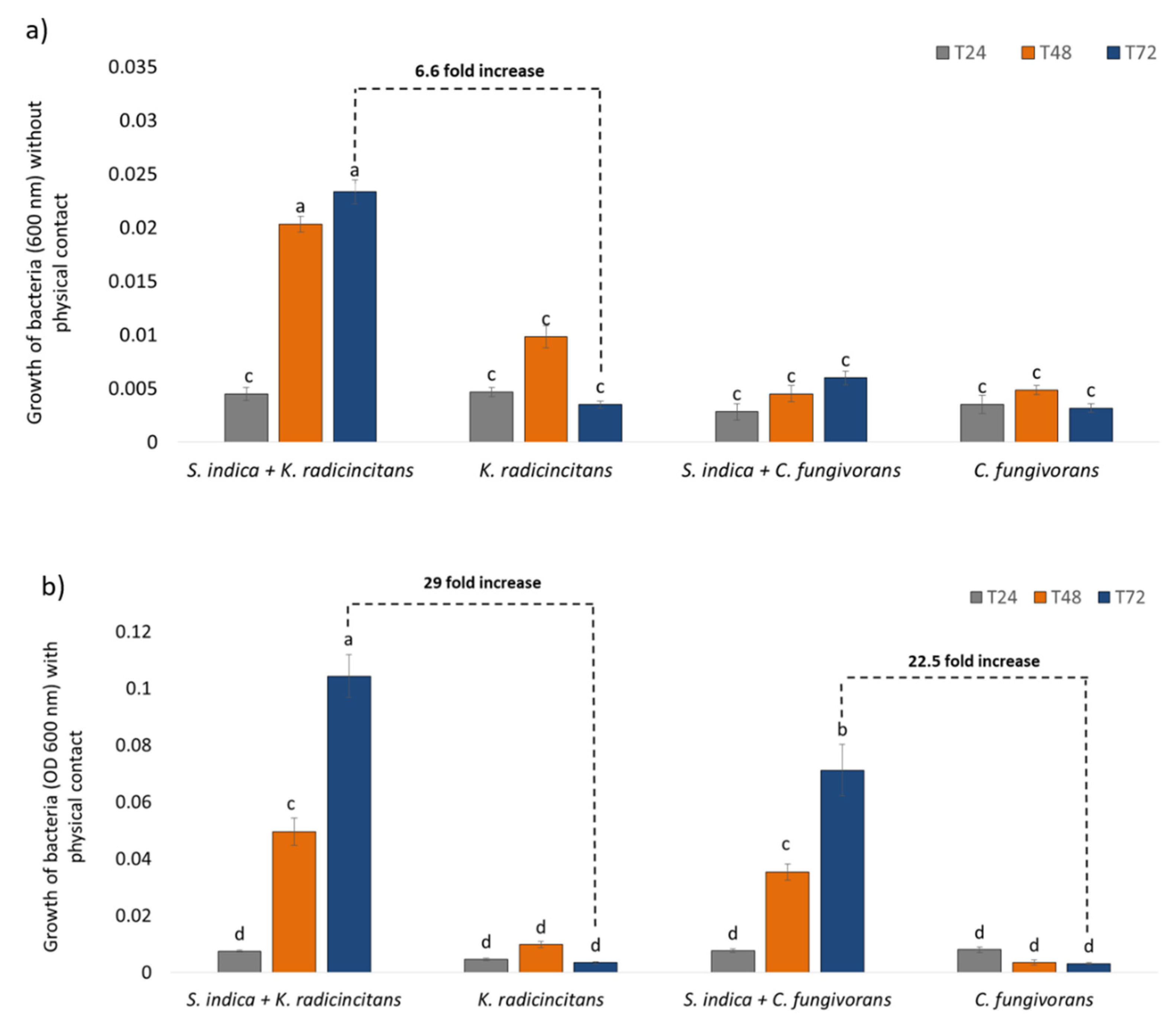
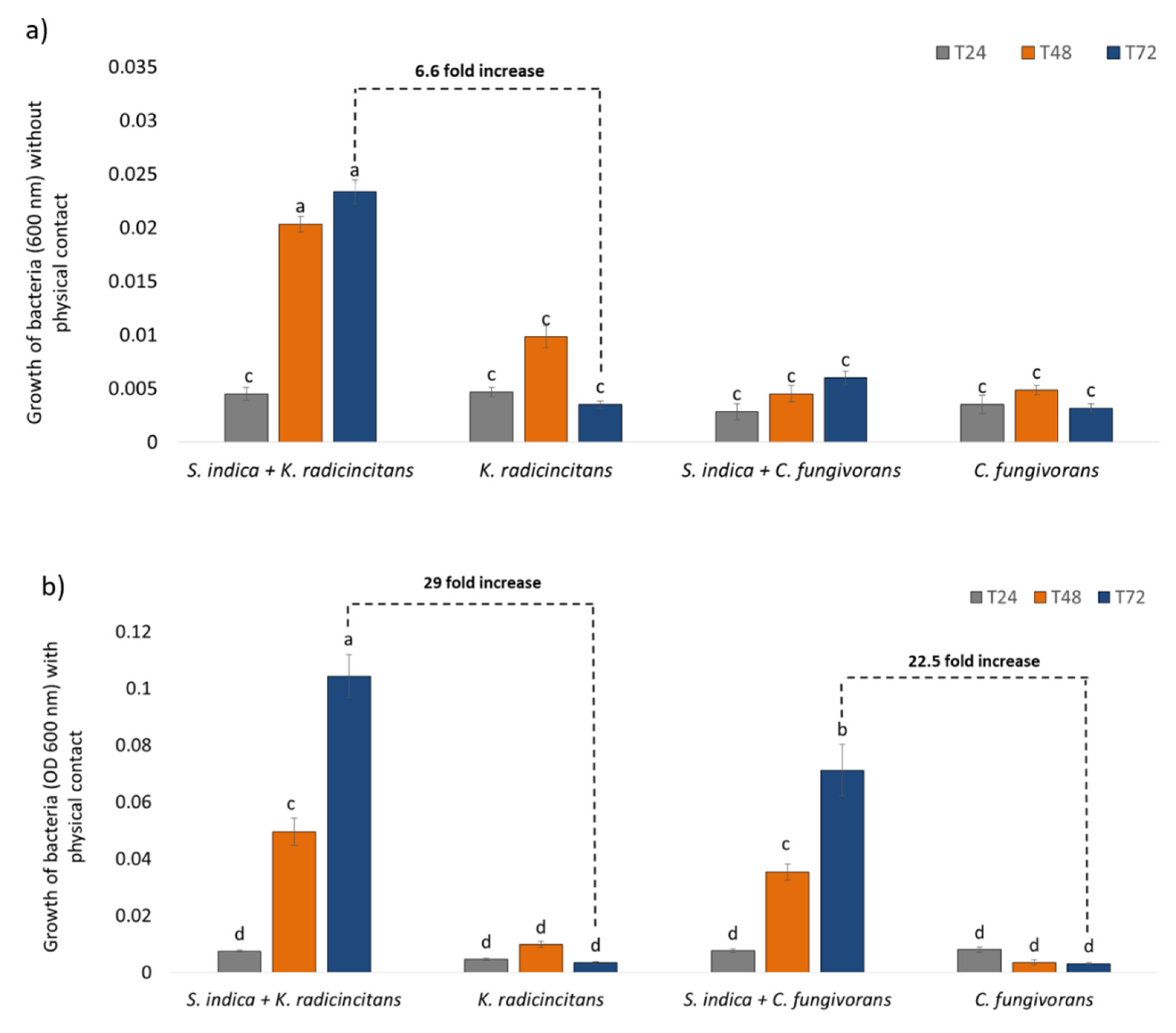
3.4. Co-Cultivation of Bacteria and Fungus (with or without Physical Contact)
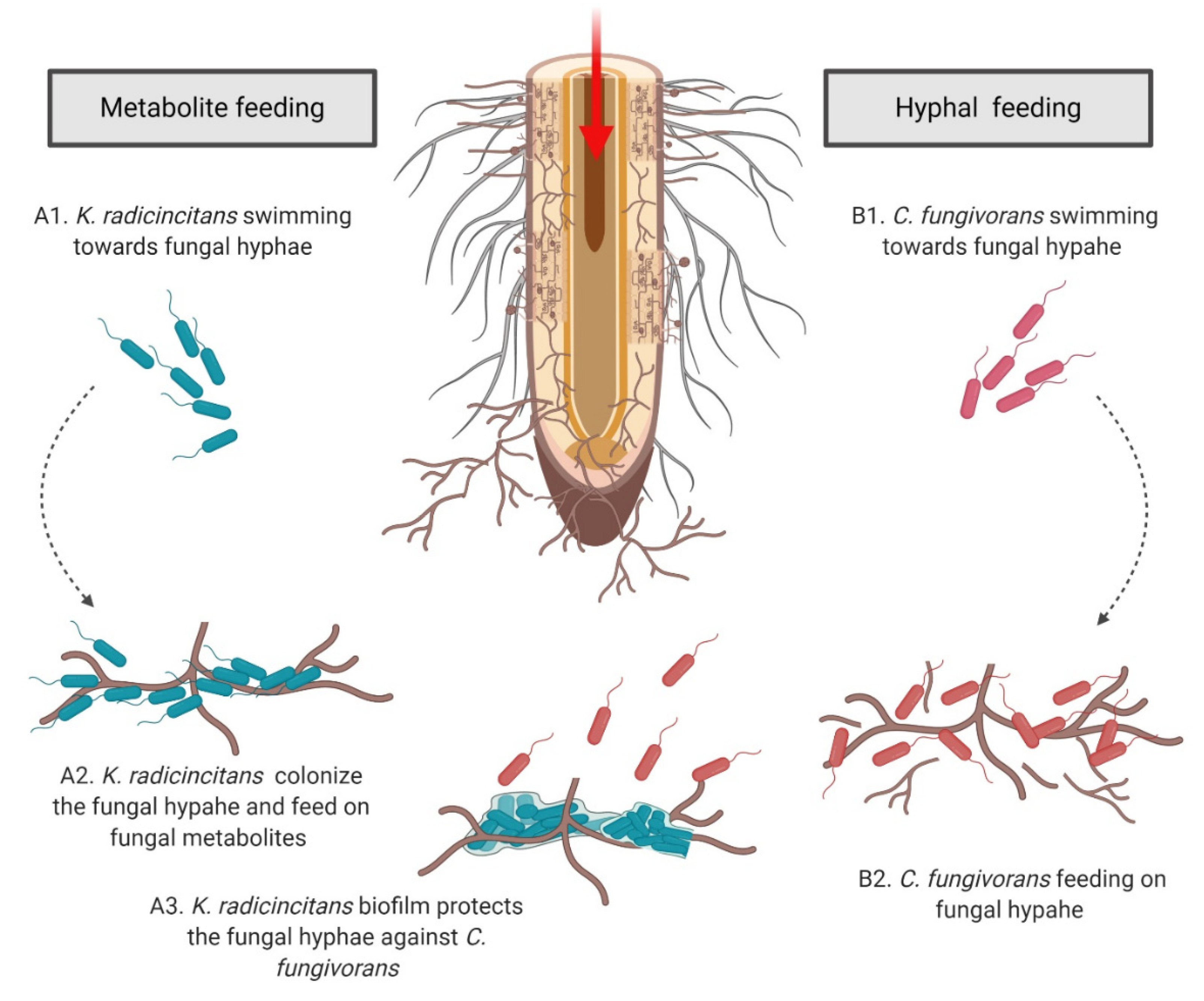
3.5. Tripartite Interaction: K. radicincitans, S. indica, and C. fungivorans
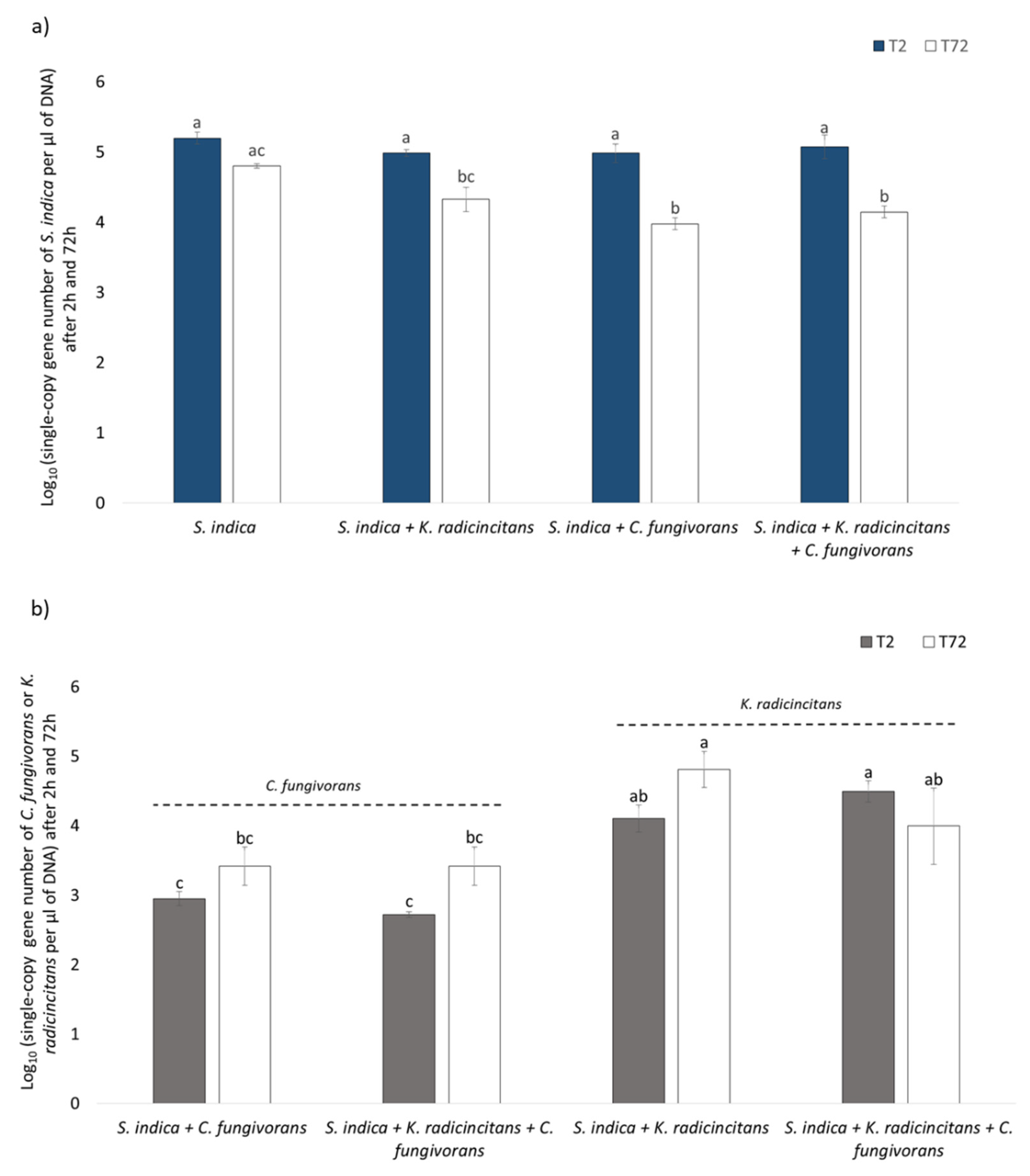
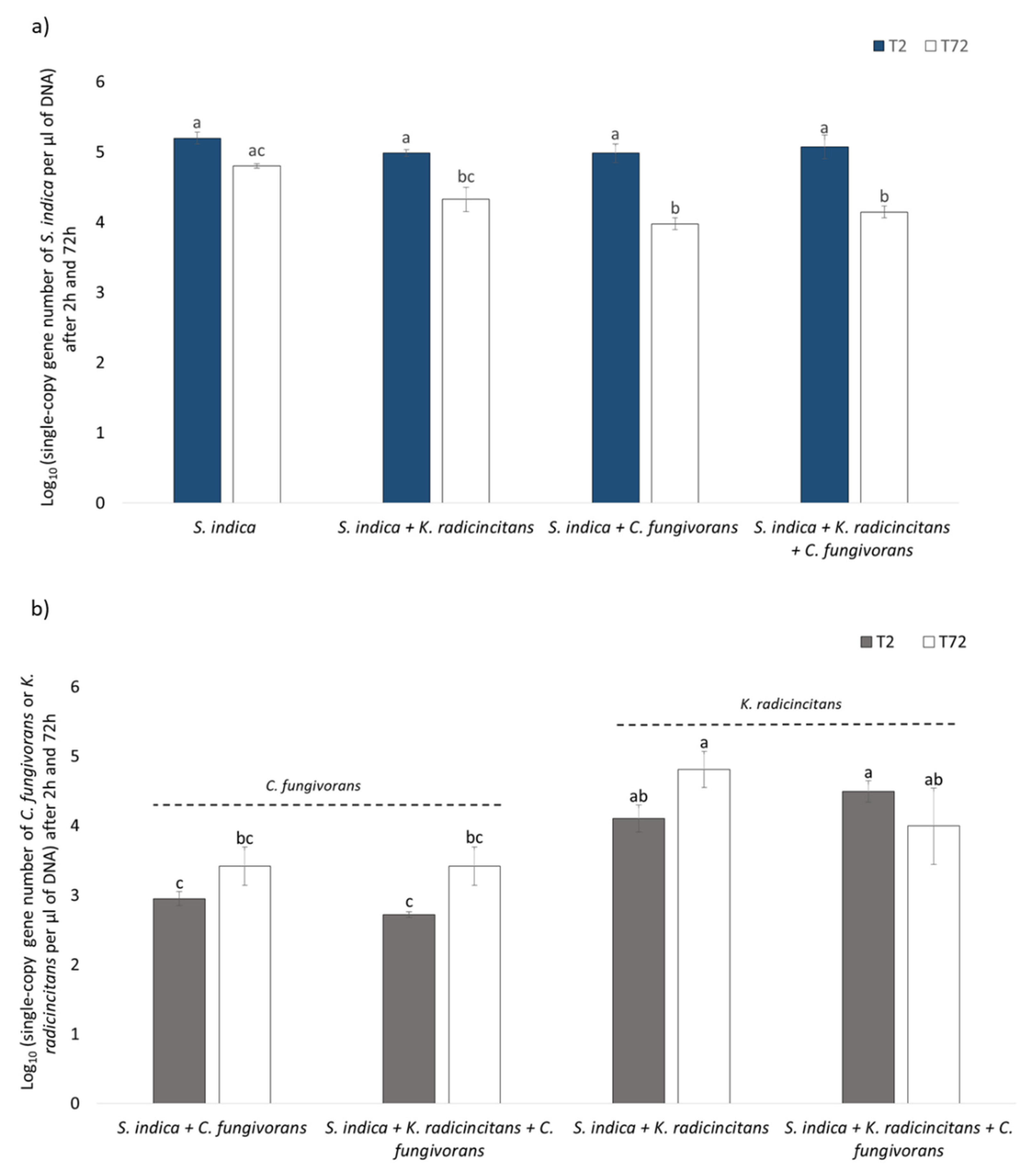
3.6. Quantification of Bacterial and Fungal Copy Numbers via qPCR
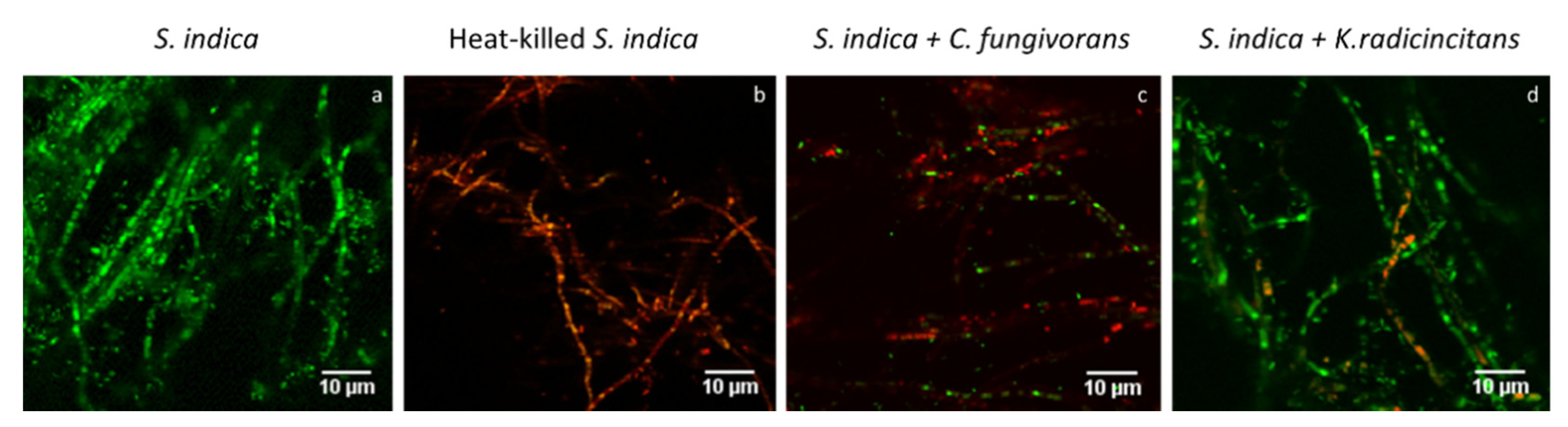
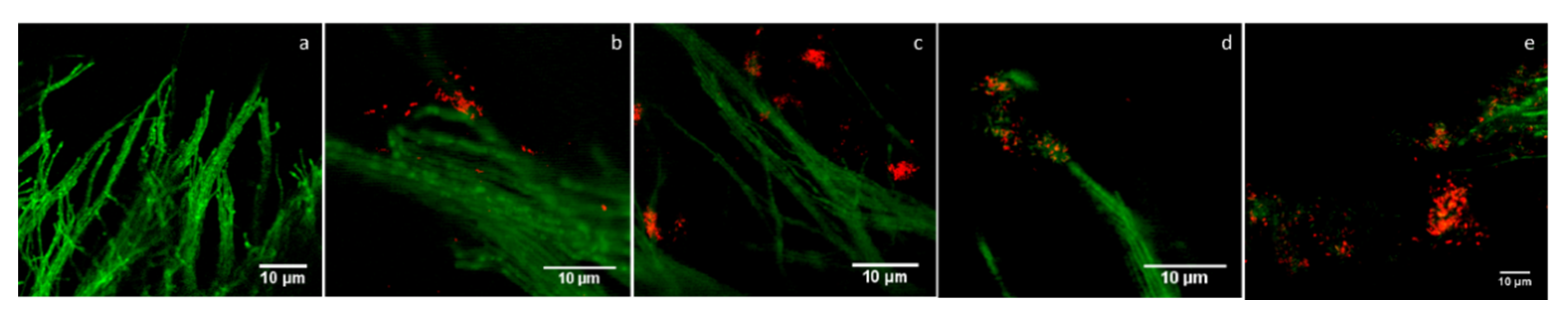
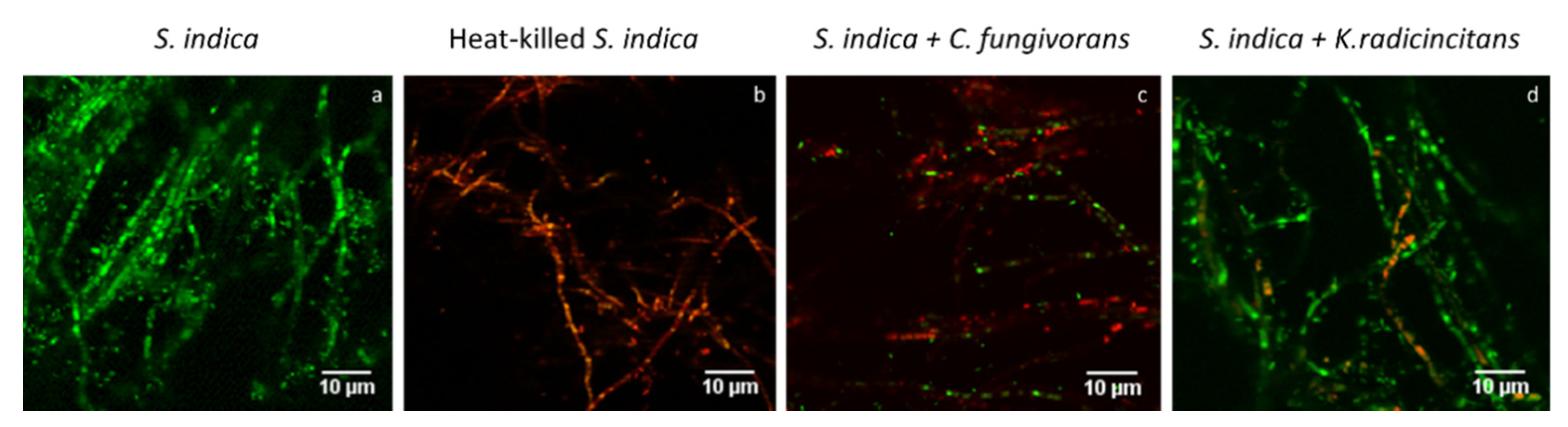
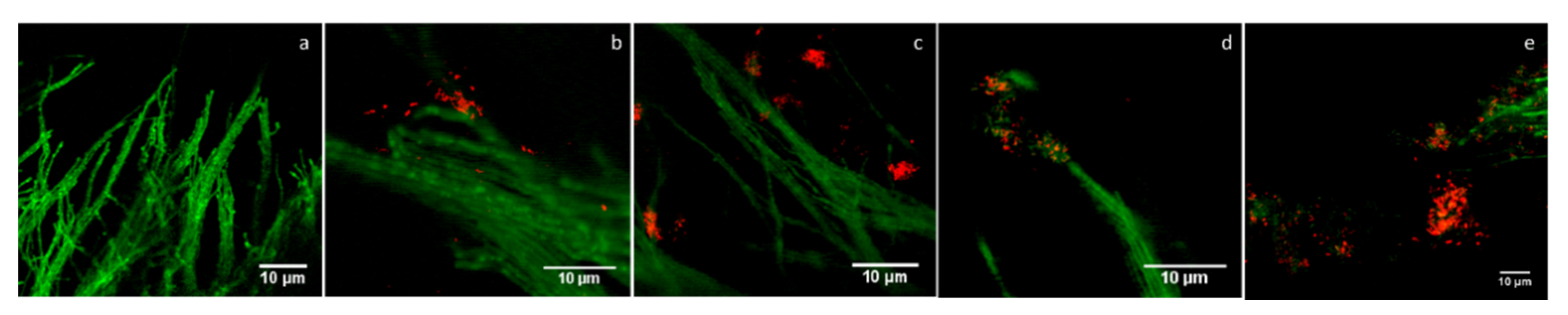
3.7. Confocal Laser Scanning Microscopy (CLSM) Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Barea, J.-M.; Pozo, M.J.; Azcón, R.; Azcón-Aguilar, C. Microbial Interactions in the Rhizosphere. In Molecular Microbial Ecol-ogy of the Rhizosphere; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 29–44. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Pratama, A.A.; De Araújo, W.L.; Trevors, J.T. Microbial Interactions in Soil. In Modern Soil Microbiology; El-sas, J.D., van Trevors, J.T., Rosado, A.S., Nannipieri, P., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 141–161. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, R. Carbon flow in the rhizosphere: Carbon trading at the soil–root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Jones, D.L.; Hodge, A.; Kuzyakov, Y. Plant and mycorrhizal regulation of rhizodeposition. New Phytol. 2004, 163, 459–480. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.; Lemanceau, P.; van der Putten, W. Going back to the roots: The microbial ecology of the rhizo-sphere. Nat. Rev. Genet. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Shamoot, S.; McDonald, L.; Bartholomew, W.V. Rhizo-Deposition of Organic Debris in Soil1. Soil Sci. Soc. Am. J. 1968, 32, 817–820. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2008, 32, 666–681. [Google Scholar] [CrossRef]
- Weixin, C.; Coleman, D.; Carroll, C.; Hoffman, C.A. In situ measurement of root respiration and soluble C concentrations in the rhizosphere. Soil Biol. Biochem. 1993, 25, 1189–1196. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Barka, E.A. Endophytic Colonization of Vitis vinifera L. by Plant Growth-Promoting Bacterium Burkholderia sp. Strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.-J.; Wang, L.-L.; Li, Q.; Shang, Q.-M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of to-mato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Francioli, D.; van Rijssel, S.Q.; van Ruijven, J.; Termorshuizen, A.J.; Cotton, T.E.A.; Dumbrell, A.J.; Raaijmakers, J.M.; Wei-gelt, A.; Mommer, L. Plant functional group drives the community structure of saprophytic fungi in a grassland biodiversity experiment. Plant Soil 2020, 461, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, I.; Rosendahl, L. Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytol. 1990, 115, 77–83. [Google Scholar] [CrossRef]
- Kaiser, C.; Kilburn, M.; Clode, P.; Fuchslueger, L.; Koranda, M.; Cliff, J.B.; Solaiman, Z.M.; Murphy, D. Exploring the transfer of recent plant photosynthates to soil microbes: Mycorrhizal pathway vs direct root exudation. New Phytol. 2015, 205, 1537–1551. [Google Scholar] [CrossRef]
- Toljander, J.F.; Lindahl, B.D.; Paul, L.R.; Elfstrand, M.; Finlay, R.D. Influence of arbuscular mycorrhizal mycelial exudates on soil bacterial growth and community structure: Effects of AM mycelial exudates on soil bacteria. FEMS Microbiol. Ecol. 2007, 61, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Akyol, T.Y.; Niwa, R.; Hirakawa, H.; Maruyama, H.; Sato, T.; Suzuki, T.; Fukunaga, A.; Sato, T.; Yoshida, S.; Tawaraya, K.; et al. Impact of Introduction of Arbuscular Mycorrhizal Fungi on the Root Microbial Community in Agricultural Fields. Microbes Environ. 2019, 34, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Ballhausen, M.-B.; de Boer, W. The sapro-rhizosphere: Carbon flow from saprotrophic fungi into fungus-feeding bacteria. Soil Biol. Biochem. 2016, 102, 14–17. [Google Scholar] [CrossRef]
- Emmett, B.D.; Lévesque-Tremblay, V.; Harrison, M.J. Conserved and reproducible bacterial communities associate with ex-traradical hyphae of arbuscular mycorrhizal fungi. ISME J. 2021, 1–13. [Google Scholar] [CrossRef]
- Zhou, J.; Chai, X.; Zhang, L.; George, T.S.; Wang, F.; Feng, G. Different Arbuscular Mycorrhizal Fungi Cocolonizing on a Sin-gle Plant Root System Recruit Distinct Microbiomes. mSystems 2020, 5, e00929-20. [Google Scholar] [CrossRef]
- Drigo, B.; Pijl, A.S.; Duyts, H.; Kielak, A.M.; Gamper, H.; Houtekamer, M.J.; Boschker, H.T.S.; Bodelier, P.; Whiteley, A.; van Veen, J.A.; et al. Shifting carbon flow from roots into associated microbial communities in response to elevated atmospheric CO2. Proc. Natl. Acad. Sci. USA 2010, 107, 10938–10942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xu, M.; Liu, Y.; Zhang, F.; Hodge, A.; Feng, G. Carbon and phosphorus exchange may enable cooperation be-tween an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 2016, 210, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Guennoc, C.M.; Rose, C.; Labbé, J.; Deveau, A. Bacterial biofilm formation on the hyphae of ectomycorrhizal fungi: A wide-spread ability under controls? FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Sharma, S.; Compant, S.; Ballhausen, M.-B.; Ruppel, S.; Franken, P. The interaction between Rhizoglomus irregulare and hyphae attached phosphate solubilizing bacteria increases plant biomass of Solanum lycopersicum. Microbiol. Res. 2020, 240, 126556. [Google Scholar] [CrossRef] [PubMed]
- Taktek, S.; St-Arnaud, M.; Piché, Y.; Fortin, J.A.; Antoun, H. Igneous phosphate rock solubilization by biofilm-forming my-corrhizobacteria and hyphobacteria associated with Rhizoglomus irregulare DAOM 197198. Mycorrhiza 2017, 27, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warmink, J.A.; van Elsas, J.D. Migratory Response of Soil Bacteria to Lyophyllum sp. Strain Karsten in Soil Microcosms. AEM 2009, 75, 2820–2830. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wingender, H.-C.F.J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Genet. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Nazir, R.; Warmink, J.A.; Boersma, H.; Van Elsas, J.D. Mechanisms that promote bacterial fitness in fungal-affected soil mi-crohabitats. FEMS Microbiol. Ecol. 2009, 71, 169–185. [Google Scholar] [CrossRef] [Green Version]
- Warmink, J.; Nazir, R.; Corten, B.; van Elsas, J. Hitchhikers on the fungal highway: The helper effect for bacterial migration via fungal hyphae. Soil Biol. Biochem. 2011, 43, 760–765. [Google Scholar] [CrossRef]
- Rossi, F. Beneficial biofilms for land rehabilitation and fertilization. FEMS Microbiol. Lett. 2020, 367. [Google Scholar] [CrossRef]
- Kuramitsu, H.K.; He, X.; Lux, R.; Anderson, M.H.; Shi, W. Interspecies Interactions within Oral Microbial Communities. Microbiol. Mol. Biol. Rev. 2007, 71, 653–670. [Google Scholar] [CrossRef] [Green Version]
- Nazir, R.; Tazetdinova, D.I.; Van Elsas, J.D. Burkholderia terrae BS001 migrates proficiently with diverse fungal hosts through soil and provides protection from antifungal agents. Front. Microbiol. 2014, 5, 598. [Google Scholar] [CrossRef] [PubMed]
- Essig, A.; Hofmann, D.; Münch, D.; Gayathri, S.; Künzler, M.; Kallio, P.T.; Sahl, H.-G.; Wider, G.; Schneider, T.; Aebi, M. Copsin, a Novel Peptide-based Fungal Antibiotic Interfering with the Peptidoglycan Synthesis. J. Biol. Chem. 2014, 289, 34953–34964. [Google Scholar] [CrossRef] [Green Version]
- Stöckli, M.; Lin, C.-W.; Sieber, R.; Plaza, D.F.; Ohm, R.A.; Künzler, M. Coprinopsis cinerea intracellular lactonases hydrolyze quorum sensing molecules of Gram-negative bacteria. Fungal Genet. Biol. 2017, 102, 49–62. [Google Scholar] [CrossRef]
- De Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche de-velopment. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Brookes, P.C.; Xu, J.; Feng, Y. Bacterial degradation of Aroclor 1242 in the mycorrhizosphere soils of zucchini (Cu-curbita pepo L.) inoculated with arbuscular mycorrhizal fungi. Environ. Sci. Pollut. Res. 2014, 21, 12790–12799. [Google Scholar] [CrossRef] [PubMed]
- Toljander, J.F.; Artursson, V.; Paul, L.R.; Jansson, J.; Finlay, R. Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae is determined by hyphal vitality and fungal species. FEMS Microbiol. Lett. 2006, 254, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, W.; Gunnewiek, P.J.K.; Lafeber, P.; Janse, J.D.; Spit, B.E.; Woldendorp, J.W. Anti-fungal properties of chitinolytic dune soil bacteria. Soil Biol. Biochem. 1998, 30, 193–203. [Google Scholar] [CrossRef]
- Leveau, J.H.J.; Preston, G. Bacterial mycophagy: Definition and diagnosis of a unique bacterial–fungal interaction. New Phytol. 2008, 177, 859–876. [Google Scholar] [CrossRef]
- Deveau, A.; Palin, B.; Delaruelle, C.; Peter, M.; Kohler, A.; Pierrat, J.C.; Sarniguet, A.; Garbaye, J.; Martin, F.; Frey-Klett, P. The mycorrhiza helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytol. 2007, 175, 743–755. [Google Scholar] [CrossRef]
- Mela, F.; Fritsche, K.; de Boer, W.; van Veen, J.A.; De Graaff, L.H.; Berg, M.V.D.; Leveau, J.H.J. Dual transcriptional profiling of a bacterial/fungal confrontation: Collimonas fungivorans versus Aspergillus niger. ISME J. 2011, 5, 1494–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, W. Collimonas fungivorans gen. nov.; sp. nov.; a chitinolytic soil bacterium with the ability to grow on living fungal hyphae. Int. J. Syst. Evol. Microbiol. 2004, 54, 857–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, W.; Gunnewiek, P.J.A.K.; Kowalchuk, G.A.; Van Veen, J.A. Growth of Chitinolytic Dune Soil β-SubclassProteobacteria in Response to Invading Fungal Hyphae. Appl. Environ. Microbiol. 2001, 67, 3358–3362. [Google Scholar] [CrossRef] [Green Version]
- Doan, H.K.; Maharaj, N.N.; Kelly, K.N.; Miyao, E.M.; Davis, R.M.; Leveau, J.H.J. Antimycotal Activity of Collimonas Isolates and Synergy-Based Biological Control of Fusarium Wilt of Tomato. Phytobiomes J. 2020, 4, 64–74. [Google Scholar] [CrossRef]
- Fritsche, K.; van den Berg, M.; de Boer, W.; van Beek, T.A.; Raaijmakers, J.M.; van Veen, J.A.; Leveau, J.H.J. Biosynthetic genes and activity spectrum of antifungal polyynes from Collimonas fungivorans Ter331: Characterization of Collimonas antifungal polyynes. Environ. Microbiol. 2014, 16, 1334–1345. [Google Scholar] [CrossRef] [Green Version]
- Hoppener-Ogawa, S.; De Boer, W.; Leveau, J.H.J.; Van Veen, J.A.; De Brandt, E.; Vanlaere, E.; Sutton, H.; Dare, D.J.; Vandamme, P. Collimonas arenae sp. nov. and Collimonas pratensis sp. nov., isolated from (semi-)natural grassland soils. Int. J. Syst. Evol. Microbiol. 2008, 58, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Höppener-Ogawa, S.; Leveau, J.H.J.; Smant, W.; van Veen, J.A.; de Boer, W. Specific Detection and Real-Time PCR Quanti-fication of Potentially Mycophagous Bacteria Belonging to the Genus Collimonas in Different Soil Ecosystems. Appl. Environ. Microbiol. 2007, 73, 4191–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, K.; Sogame, M.; Sakurai, F.; Nasu, N.; Fujita, M. Collimonins A–D, Unstable Polyynes with Antifungal or Pigmentation Activities from the Fungus-Feeding BacteriumCollimonas fungivoransTer331. Org. Lett. 2018, 20, 3536–3540. [Google Scholar] [CrossRef]
- Song, C.; Schmidt, R.; De Jager, V.; Krzyzanowska, D.; Jongedijk, E.; Cankar, K.; Beekwilder, J.; Van Veen, A.; De Boer, W.; Van Veen, J.A.; et al. Exploring the genomic traits of fungus-feeding bacterial genus Collimonas. BMC Genom. 2015, 16, 1103. [Google Scholar] [CrossRef] [PubMed]
- Ballhausen, M.-B.; van Veen, J.A.; Hundscheid, M.P.J.; de Boer, W. Methods for Baiting and Enriching Fungus-Feeding (My-cophagous) Rhizosphere Bacteria. Front. Microbiol. 2015, 6, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivato, B.; Offre, P.; Marchelli, S.; Barbonaglia, B.; Mougel, C.; Lemanceau, P.; Berta, G. Bacterial effects on arbuscular my-corrhizal fungi and mycorrhiza development as influenced by the bacteria, fungi, and host plant. Mycorrhiza 2008, 19, 81–90. [Google Scholar] [CrossRef]
- Höppener-Ogawa, S.; Leveau, J.H.; van Veen, J.A.; De Boer, W. Mycophagous growth of Collimonas bacteria in natural soils, impact on fungal biomass turnover and interactions with mycophagous Trichoderma fungi. ISME J. 2008, 3, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Höppener-Ogawa, S.; Leveau, J.H.J.; Hundscheid, M.P.J.; van Veen, J.A.; de Boer, W. Impact of Collimonas bacteria on community composition of soil fungi. Environ. Microbiol. 2009, 11, 1444–1452. [Google Scholar] [CrossRef]
- Haq, I.U.; Zhang, M.; Yang, P.; van Elsas, J.D. The Interactions of Bacteria with Fungi in Soil. In Advances in Clinical Chemis-try; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 89, pp. 185–215. [Google Scholar]
- Stopnišek, N.; Zühlke, D.; Carlier, A.; Barberán, A.; Fierer, N.; Becher, D.; Riedel, K.; Eberl, L.; Weisskopf, L. Molecular mechanisms underlying the close association between soil Burkholderia and fungi. ISME J. 2015, 10, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Jan-Roblero, J.; Cruz-Maya, J.A.; Barajas, C.G. Kosakonia. In Beneficial Microbes in Agro-Ecology; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 213–231. [Google Scholar]
- Romano, I.; Ventorino, V.; Ambrosino, P.; Testa, A.; Chouyia, F.E.; Pepe, O. Development and Application of Low-Cost and Eco-Sustainable Bio-Stimulant Containing a New Plant Growth-Promoting Strain Kosakonia pseudosacchari TL13. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Taulé, C.; Luizzi, H.; Beracochea, M.; Mareque, C.; Platero, R.; Battistoni, F. The Mo- and Fe-nitrogenases of the endophyte Kosakonia sp. UYSO10 are necessary for growth promotion of sugarcane. Ann. Microbiol. 2019, 69, 741–750. [Google Scholar] [CrossRef]
- Ruppel, S.; Merbach, W. Effects of different nitrogen sources on nitrogen fixation and bacterial growth of Pantoea agglomer-ans and Azospirillum sp. in bacterial pure culture: An investigation using 15N2 incorporation and acetylene reduction measures. Microbiol. Res. 1995, 150, 409–418. [Google Scholar] [CrossRef]
- Berger, B.; Brock, A.K.; Ruppel, S. Nitrogen supply influences plant growth and transcriptional responses induced by Enter-obacter radicincitans in Solanum lycopersicum. Plant Soil 2013, 370, 641–652. [Google Scholar] [CrossRef]
- Brock, A.K.; Berger, B.; Mewis, I.; Ruppel, S. Impact of the PGPB Enterobacter radicincitans DSM 16656 on Growth, Glucos-inolate Profile, and Immune Responses of Arabidopsis thaliana. Microb. Ecol. 2013, 65, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Krey, T.; Caus, M.; Baum, C.; Ruppel, S.; Eichler-Löbermann, B. Interactive effects of plant growth-promoting rhizobacteria and organic fertilization on P nutrition of Zea mays L. and Brassica napus L. J. Plant Nutr. Soil Sci. USA 2011, 174, 602–613. [Google Scholar] [CrossRef]
- Remus, R.; Rüppel, S.; Jacob, H.-J.; Hecht-Buchholz, C.; Merbach, W. Colonization behaviour of two enterobacterial strains on cereals. Biol. Fertil. Soils 2000, 30, 550–557. [Google Scholar] [CrossRef]
- Becker, M.; Patz, S.; Becker, Y.; Berger, B.; Drungowski, M.; Bunk, B.; Overmann, J.; Sprör, C.; Reetz, J.; Tchuisseu, GV.; et al. Comparative Genomics Reveal a Flagellar System, a Type VI Secretion System and Plant Growth-Promoting Gene Clusters Unique to the Endophytic Bacterium Kosakonia radicincitans. Front. Microbiol. 2018, 9, 1997. [Google Scholar] [CrossRef] [Green Version]
- Varma, A.; Sherameti, I.; Tripathi, S.; Prasad, R.; Das, A.; Sharma, M.; Bakshi, M.; Johnson, J.M.; Bhardwaj, S.; Arora, M.; et al. 13 The Symbiotic Fungus Piriformospora indica: Review. In Fungal Associations; Springer Science and Business Media LLC: Berlin, Germany, 2012; pp. 231–254. [Google Scholar]
- Selosse, M.-A.; Roy, M. Green plants that feed on fungi: Facts and questions about mixotrophy. Trends Plant Sci. 2009, 14, 64–70. [Google Scholar] [CrossRef]
- Weiß, M.; Sýkorová, Z.; Garnica, S.; Riess, K.; Martos, F.; Krause, C.; Oberwinkler, F.; Bauer, R.; Redecker, D. Sebac-inales Everywhere: Previously Overlooked Ubiquitous Fungal Endophytes. PLoS ONE 2011, 6, e16793. [Google Scholar] [CrossRef]
- Oelmüller, R.; Sherameti, I.; Tripathi, S.; Varma, A. Piriformospora indica, a cultivable root endophyte with multiple bio-technological applications. Symbiosis 2009, 49, 1–17. [Google Scholar] [CrossRef]
- Qiang, X.; Weiss, M.; Kogel, K.-H.; Schäfer, P. Piriformospora indica—A mutualistic basidiomycete with an exceptionally large plant host range: Mutualistic root symbiosis. Mol. Plant Pathol. 2012, 13, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Franken, P. The plant strengthening root endophyte Piriformospora indica: Potential application and the biology behind. Appl. Microbiol. Biotechnol. 2012, 96, 1455–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varma, A.; Bakshi, M.; Lou, B.; Hartmann, A.; Oelmueller, R. Piriformospora indica: A Novel Plant Growth-Promoting My-corrhizal Fungus. Agric. Res. 2012, 1, 117–131. [Google Scholar] [CrossRef]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; von Wettstein, D.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [Green Version]
- Bhuyan, S.K.; Bandyopadhyay, P.; Kumar, P.; Mishra, D.K.; Prasad, R.R.; Kumari, A.; Upadhyaya, K.C.; Varma, A.; Yadava, P.K. Interaction of Piriformospora indica with Azotobacter chroococcum. Sci. Rep. 2015, 5, 13911. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatiles produced by the mycophagous soil bacteriumCollimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Kamilova, F.; Leveau, J.H.J.; Lugtenberg, B. Collimonas fungivorans, an unpredicted in vitro but efficient in vivo biocontrol agent for the suppression of tomato foot and root rot. Environ. Microbiol. 2007, 9, 1597–1603. [Google Scholar] [CrossRef]
- Bending, G.D.; Poole, E.J.; Whipps, J.M.; Read, D.J. Characterisation of bacteria from Pinus sylvestris–Suillus luteus mycor-rhizas and their effects on root–fungus interactions and plant growth. FEMS Microbiol. Ecol. 2002, 39, 219–227. [Google Scholar] [PubMed]
- Levy, A.; Chang, B.J.; Abbott, L.; Kuo, J.; Harnett, G.; Inglis, T.J.J. Invasion of Spores of the Arbuscular Mycorrhizal Fungus Gigaspora decipiens by Burkholderia spp. Appl. Environ. Microbiol. 2003, 69, 6250–6256. [Google Scholar] [CrossRef] [Green Version]
- Mansfeld-Giese, K.; Larsen, J.; Bødker, L. Bacterial populations associated with mycelium of the arbuscular mycorrhizal fungus Glomus intraradices. FEMS Microbiol. Ecol. 2002, 41, 133–140. [Google Scholar] [CrossRef]
- Herrera, H.; Sanhueza, T.; Novotná, A.; Charles, T.C.; Arriagada, C. Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile. Diversity 2020, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Wright, S.F.; Nichols, K.A. Using glomalin as an indicator for arbuscular mycorrhizal hyphal growth: An example from a tropical rain forest soil. Soil Biol. Biochem. 2004, 36, 1009–1012. [Google Scholar] [CrossRef]
- Olsson, P.A.; Larsson, L.; Bago, B.; Wallander, H.; Van Aarle, I.M. Ergosterol and fatty acids for biomass estimation of my-corrhizal fungi. New Phytol. 2003, 159, 7–10. [Google Scholar] [CrossRef] [Green Version]
- Wallander, H.; Ekblad, A.; Godbold, D.; Johnson, D.; Bahr, A.; Baldrian, P.; Björk, R.; Kieliszewska-Rokicka, B.; Kjøller, R.; Kraigher, H.; et al. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in for-ests soils—A review. Soil Biol. Biochem. 2013, 57, 1034–1047. [Google Scholar] [CrossRef]
- Verma, S.; Varma, A.; Rexer, K.-H.; Hassel, A.; Kost, G.; Sarbhoy, A.; Franken, P. Piriformospora indica, gen. et sp. nov.; a New Root-Colonizing Fungus. Mycologia 1998, 90, 896. [Google Scholar] [CrossRef]
- Pham, G.H.; Kumari, R.; Singh, A.; Malla, R.; Prasad, R.; Sachdev, M.; Kaldorf, M.; Buscot, F.; Oelmüller, R.; Hampp, R.; et al. Axenic Culture of Symbiotic Fungus Piriformospora indica. In Plant Surface Microbiology; Springer Science and Business Media LLC: Berlin, Germany, 2008; pp. 593–613. [Google Scholar]
- De Boer, W.; Wagenaar, A.-M.; Klein Gunnewiek, P.J.A.; Van Veen, J.A. In vitro suppression of fungi caused by combinations of apparently non-antagonistic soil bacteria: Suppression of fungi by non-antagonistic soil bacteria. FEMS Microbiol. Ecol. 2007, 59, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, L.; Sutherland, I.W. Polysaccharide lyases from gellan-producing Sphingomonas spp. Microbiology 1996, 142, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranenbrouck, S.; Voets, L.; Bivort, C.; Renard, L.; Strullu, D.-G.; Declerck, S. Methodologies for in Vitro Cultivation of Ar-buscular Mycorrhizal Fungi with Root Organs. In Soil Biology; Springer Science and Business Media LLC: Berlin, Germany, 2005; Volume 4, pp. 341–375. [Google Scholar]
- Egli, T. Microbial growth and physiology: A call for better craftsmanship. Front. Microbiol. 2015, 6, 287. [Google Scholar] [CrossRef]
- Guennoc, C.M.; Rose, C.; Guinnet, F.; Miquel, I.; Labbé, J.; Deveau, A. A New Method for Qualitative Multi-scale Analysis of Bacterial Biofilms on Filamentous Fungal Colonies Using Confocal and Electron Microscopy. J. Vis. Exp. 2017, e54771. [Google Scholar] [CrossRef] [Green Version]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Rasmussen, R. Quantification on the LightCycler. In Rapid Cycle Real-Time PCR; Springer Science and Business Media LLC: Berlin, Germany, 2001; pp. 21–34. [Google Scholar]
- Bütehorn, B.; Rhody, D.; Franken, P. Isolation and Characterisation of Pitef1 Encoding the Translation Elongation Factor EF-1α of the Root Endophyte Piriformospora indica. Plant Biol. 2000, 2, 687–692. [Google Scholar] [CrossRef]
- Ruppel, S.; Rühlmann, J.; Merbach, W. Quantification and Localization of Bacteria in Plant Tissues Using Quantitative Real-Time PCR and Online Emission Fingerprinting. Plant Soil 2006, 286, 21–35. [Google Scholar] [CrossRef]
- Boulos, L.; Prévost, M.; Barbeau, B.; Coallier, J.; Desjardins, R. LIVE/DEAD® BacLightTM: Application of a new rapid stain-ing method for direct enumeration of viable and total bacteria in drinking water. J. Microbiol. Methods 1999, 37, 77–86. [Google Scholar] [CrossRef]
- Witzel, K.; Gwinn-Giglio, M.; Nadendla, S.; Shefchek, K.; Ruppel, S. Genome Sequence of Enterobacter radicincitans DSM16656 T, a Plant Growth-Promoting Endophyte. J. Bacteriol. 2012, 194, 5469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbontín, R.; Vlamakis, H.; Kolter, R. Mutualistic interaction between Salmonella enterica and Aspergillus niger and its effects on Zea mays colonization. Microb. Biotechnol. 2014, 7, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveau, A.; Bonito, G.; Uehling, J.; Paoletti, M.; Becker, M.; Bindschedler, S.; Hacquard, S.; Hervé, V.; Labbé, J.; Lastovetsky, O.; et al. Bacterial–fungal interactions: Ecology, mechanisms and challenges. FEMS Microbiol. Rev. 2018, 42, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-Fungal Interactions: Hyphens between Agricultural, Clinical, Environmental, and Food Microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [Green Version]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.-W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [Green Version]
- Haq, I.U.; Zwahlen, R.D.; Yang, P.; Van Elsas, J.D. The Response of Paraburkholderia terrae Strains to Two Soil Fungi and the Potential Role of Oxalate. Front. Microbiol. 2018, 9, 989. [Google Scholar] [CrossRef]
- Nazir, R.; Semenov, A.V.; Sarigül, N.; Van Elsas, J.D. Bacterial community establishment in native and non-native soils and the effect of fungal colonization. Microbiol. Discov. 2013, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Berger, B.; Patz, S.; Ruppel, S.; Dietel, K.; Faetke, S.; Junge, H.; Becker, M. Successful Formulation and Application of Plant Growth-Promoting Kosakonia radicincitans in Maize Cultivation. BioMed Res. Int. 2018, 2018, 6439481. [Google Scholar] [CrossRef] [Green Version]
- Berger, B.; Wiesner, M.; Brock, A.K.; Schreiner, M.; Ruppel, S.K. radicincitans, a beneficial bacteria that promotes radish growth under field conditions. Agron. Sustain. Dev. 2015, 35, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Schluter, J.; Nadell, C.D.; Bassler, B.; Foster, K.R. Adhesion as a weapon in microbial competition. ISME J. 2015, 9, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, L.; Zhou, J.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi enhance mineralisation of organic phos-phorus by carrying bacteria along their extraradical hyphae. New Phytol. 2021, 230, 304–315. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Smits, T.H.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the Fungal Highway: Mobilization of Pollutant-Degrading Bacteria by Fungi. Environ. Sci. Technol. 2005, 39, 4640–4646. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Bindschedler, S.; Job, D.; Wick, L.Y.; Filippidou, S.; Kooli, W.M.; Verrecchia, E.P.; Junier, P. Exploiting the fungal highway: Development of a novel tool for the in situ isolation of bacteria migrating along fungal mycelium. FEMS Microbiol. Ecol. 2015, 91, fiv116. [Google Scholar] [CrossRef] [Green Version]
- Raghupathi, P.K.; Liu, W.; Sabbe, K.; Houf, K.; Burmølle, M.; Sørensen, S.J.; Raghupathi, P.K.; Liu, W.; Sabbe, K.; Houf, K.; et al. Synergistic Interactions within a Multispecies Biofilm Enhance Individual Species Protection against Grazing by a Pelagic Protozoan. Front. Microbiol. 2018, 8, 2649. [Google Scholar] [CrossRef] [Green Version]
- Seiler, C.; Van Velzen, E.; Neu, T.; Gaedke, U.; Berendonk, T.U.; Weitere, M. Grazing resistance of bacterial biofilms: A matter of predators’ feeding trait. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shi, N.; Fan, J.; Wang, F.; George, T.; Feng, G. Arbuscular mycorrhizal fungi stimulate organic phosphate mobili-zation associated with changing bacterial community structure under field conditions. Environ. Microbiol. 2018, 20, 2639–2651. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, G.; Declerck, S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phyt-ate mineralization by a phosphate solubilizing bacterium. ISME J. 2018, 12, 2339–2351. [Google Scholar] [CrossRef] [Green Version]
- Zuccaro, A.; Lahrmann, U.; Güldener, U.; Langen, G.; Pfiffi, S.; Biedenkopf, D.; Wong, P.; Samans, B.; Grimm, C.; Basiewicz, M.; et al. Endophytic Life Strategies Decoded by Genome and Transcriptome Analyses of the Mutualistic Root Sym-biont Piriformospora indica. PLoS Pathog. 2011, 7, e1002290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorka, S.; Dietrich, M.; Mayerhofer, W.; Gabriel, R.; Wiesenbauer, J.; Martin, V.; Zheng, Q.; Imai, B.; Prommer, J.; Weidinger, M.; et al. Rapid Transfer of Plant Photosynthates to Soil Bacteria via Ectomycorrhizal Hyphae and Its Interaction with Nitrogen Availability. Front. Microbiol. 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.A.; Pieterse, C.M.J. The rhizosphere revisited: Root mi-crobiomics. Front. Plant Sci. 2013, 4, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Veen, J.A.; van Overbeek, L.S.; van Elsas, J.D. Fate and activity of microorganisms introduced into soil. Microbiol. Mol. Biol Rev. 1997, 61, 121–135. [Google Scholar] [CrossRef]
- Acea, M.J.; Moore, C.R.; Alexander, M. Survival and growth of bacteria introduced into soil. Soil Biol. Biochem. 1988, 20, 509–515. [Google Scholar] [CrossRef]
- Bidyarani, N.; Prasanna, R.; Babu, S.; Hossain, F.; Saxena, A.K. Enhancement of plant growth and yields in Chickpea (Cicer arietinum L.) through novel cyanobacterial and biofilmed inoculants. Microbiol. Res. 2016, 188–189, 97–105. [Google Scholar] [CrossRef]
- Sudadi; Suryono; Triharyanto, E. The application of biofilm biofertilizer-based organic fertilizer to increase available soil nu-trients and spinach yield on dry land (a study case in Lithosol soil type). In Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2018; Volume 200, p. 012006. [Google Scholar] [CrossRef]
- Jayasinghearachchi, H.S.; Seneviratne, G. A bradyrhizobial-Penicillium spp. biofilm with nitrogenase activity improves N2 fixing symbiosis of soybean. Biol. Fertil. Soils 2004, 40, 432–434. [Google Scholar] [CrossRef]
- Seneviratne, G.; Jayasinghearachchi, H. A rhizobial biofilm with nitrogenase activity alters nutrient availability in a soil. Soil Biol. Biochem. 2005, 37, 1975–1978. [Google Scholar] [CrossRef]
- Seneviratne, G.; Zavahir, J.S.; Bandara, W.M.M.S.; Weerasekara, M.L.M.A.W. Fungal-bacterial biofilms: Their development for novel biotechnological applications. World J. Microbiol. Biotechnol. 2008, 24, 739–743. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, S.; Compant, S.; Franken, P.; Ruppel, S.; Ballhausen, M.-B. It Takes Two to Tango: A Bacterial Biofilm Provides Protection against a Fungus-Feeding Bacterial Predator. Microorganisms 2021, 9, 1566. https://doi.org/10.3390/microorganisms9081566
Sharma S, Compant S, Franken P, Ruppel S, Ballhausen M-B. It Takes Two to Tango: A Bacterial Biofilm Provides Protection against a Fungus-Feeding Bacterial Predator. Microorganisms. 2021; 9(8):1566. https://doi.org/10.3390/microorganisms9081566
Chicago/Turabian StyleSharma, Shubhangi, Stéphane Compant, Philipp Franken, Silke Ruppel, and Max-Bernhard Ballhausen. 2021. "It Takes Two to Tango: A Bacterial Biofilm Provides Protection against a Fungus-Feeding Bacterial Predator" Microorganisms 9, no. 8: 1566. https://doi.org/10.3390/microorganisms9081566