
The Role of the Respiratory Microbiome and Viral Presence in Lower Respiratory Tract Infection Severity in the First Five Years of Life

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population and Study Design
2.2. Outcomes
2.3. Data and Sample Collection
2.4. Microbiome Sample Processing
2.5. Virology Procedures
2.6. Analysis
2.6.1. Microbiome Data Analysis
2.6.2. Statistical Analysis
3. Results
3.1. Study Population
3.2. Virology
3.3. Association between Viral Infection and Clinical Outcome Measures
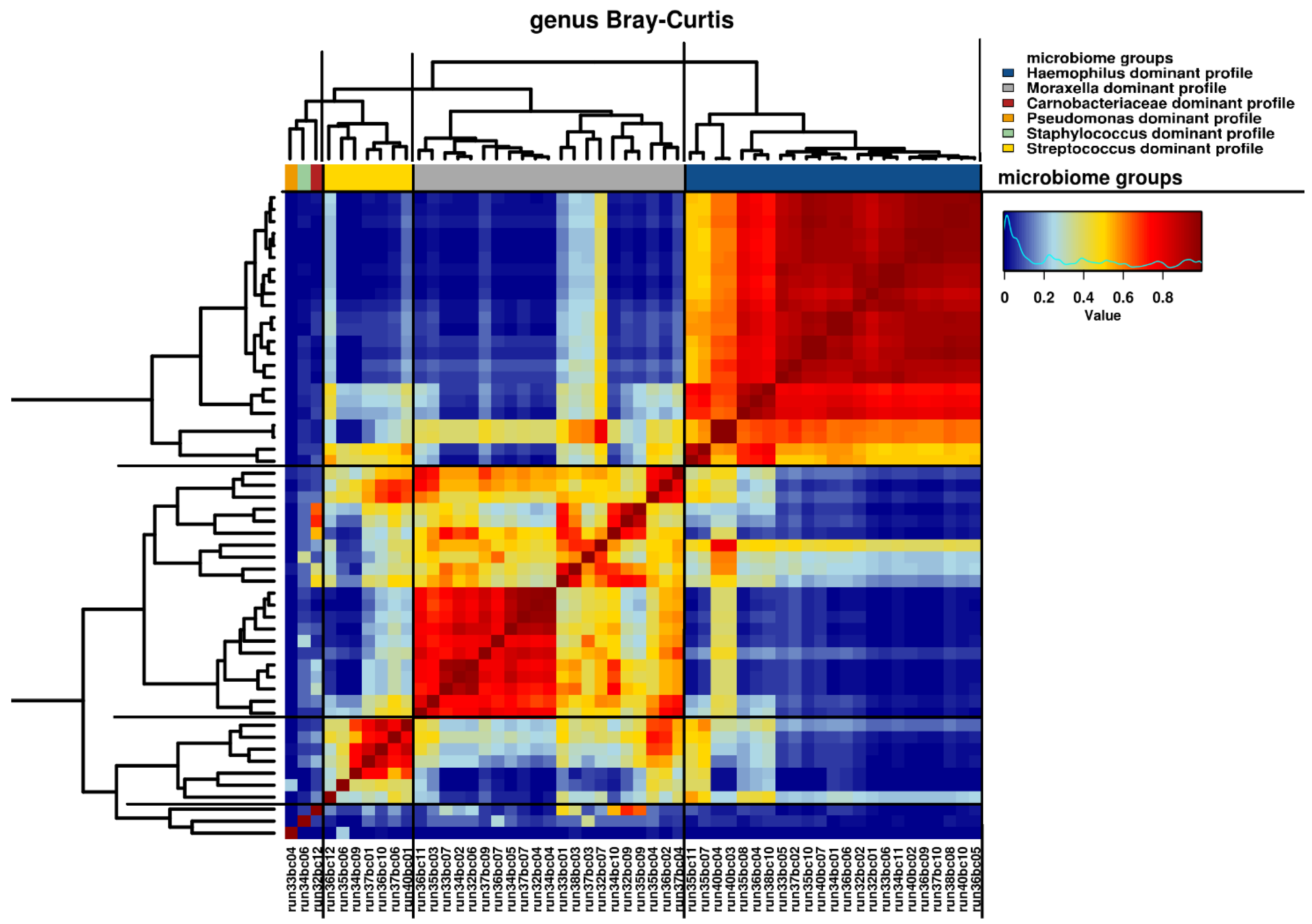
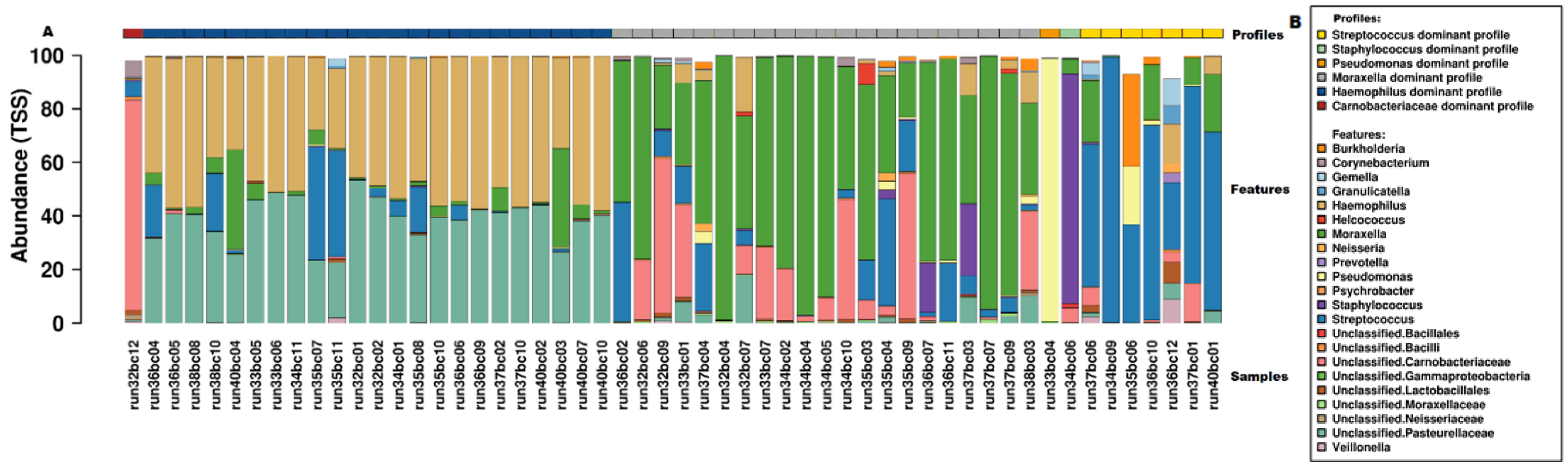
3.4. Microbiome
3.5. Integrating Viral, Microbiome, and Clinical Data
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaborators GBDCoD. Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1151–1210. [Google Scholar] [CrossRef] [Green Version]
- Collaborators GBDLRI. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Tsugawa, Y.; Mansbach, J.M.; Camargo, C.A., Jr.; Hasegawa, K. Trends in Infectious Disease Hospitalizations in US Children, 2000 to 2012. Pediatr. Infect. Dis. J. 2016, 35, e158–e163. [Google Scholar] [CrossRef] [Green Version]
- Tramper-Stranders, G.A. Childhood community-acquired pneumonia: A review of etiology- and antimicrobial treatment studies. Paediatr. Respir. Rev. 2018, 26, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Rudan, I.; O’Brien, K.L.; Nair, H.; Liu, L.; Theodoratou, E.; Qazi, S.; Luksic, I.; Fischer Walker, C.L.; Black, R.E.; Campbell, H.; et al. Epidemiology and etiology of childhood pneumonia in 2010: Estimates of incidence, severe morbidity, mortality, underlying risk factors and causative pathogens for 192 countries. J. Glob. Health 2013, 3, 010401. [Google Scholar]
- Ortiz-Hernández, A.A.; Nishimura, K.K.; Noyola, D.E.; Moreno-Espinosa, S.; Gamiño, A.; Galindo-Fraga, A.; Valdéz Vázquez, R.; Magaña Aquino, M.; Ramirez-Venegas, A.; Valdés Salgado, R.; et al. Differential risk of hospitalization among single virus infections causing influenza-like illnesses. Influenza Other Respir. Viruses 2019, 13, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Neu, U.; Mainou, B.A. Virus interactions with bacteria: Partners in the infectious dance. PLoS Pathog. 2020, 16, e1008234. [Google Scholar] [CrossRef] [PubMed]
- McCauley, K.; Durack, J.; Valladares, R.; Fadrosh, D.W.; Lin, D.L.; Calatroni, A.; LeBeau, P.K.; Tran, H.T.; Fujimura, K.E.; LaMere, B.; et al. Distinct nasal airway bacterial microbiotas differentially relate to exacerbation in pediatric patients with asthma. J. Allergy Clin. Immunol. 2019, 144, 1187–1197. [Google Scholar] [CrossRef]
- Hasegawa, K.; Linnemann, R.W.; Mansbach, J.M.; Ajami, N.J.; Espinola, J.A.; Petrosino, J.F.; Piedra, P.A.; Stevenson, M.D.; Sullivan, A.F.; Thompson, A.D.; et al. Nasal Airway Microbiota Profile and Severe Bronchiolitis in Infants: A Case-control Study. Pediatr. Infect. Dis. J. 2017, 36, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; van Houten, M.A.; Mérelle, M.E.; Vlieger, A.M.; Chu, M.; Jansen, N.J.G.; Sanders, E.A.M.; Bogaert, D. Bacterial and viral respiratory tract microbiota and host characteristics in children with lower respiratory tract infections: A matched case-control study. Lancet Respir. Med. 2019, 7, 417–426. [Google Scholar] [CrossRef]
- Hasegawa, K.; Mansbach, J.M.; Ajami, N.J.; Espinola, J.A.; Henke, D.M.; Petrosino, J.F.; Piedra, P.A.; Shaw, C.A.; Sullivan, A.F.; Camargo, C.A., Jr.; et al. Association of nasopharyngeal microbiota profiles with bronchiolitis severity in infants hospitalised for bronchiolitis. Eur. Respir. J. 2016, 48, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- de Steenhuijsen Piters, W.A.; Heinonen, S.; Hasrat, R.; Bunsow, E.; Smith, B.; Suarez-Arrabal, M.C.; Chaussabel, D.; Cohen, D.M.; Sanders, E.A.; Ramilo, O.; et al. Nasopharyngeal Microbiota, Host Transcriptome, and Disease Severity in Children with Respiratory Syncytial Virus Infection. Am. J. Respir. Crit. Care Med. 2016, 194, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- van de Maat, J.S.; Peeters, D.; Nieboer, D.; van Wermeskerken, A.M.; Smit, F.J.; Noordzij, J.G.; Tramper-Stranders, G.; Driessen, G.J.A.; Obihara, C.C.; Punt, J.; et al. Evaluation of a clinical decision rule to guide antibiotic prescription in children with suspected lower respiratory tract infection in The Netherlands: A stepped-wedge cluster randomised trial. PLoS Med. 2020, 17, e1003034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijman, R.G.; Vergouwe, Y.; Moll, H.A.; Smit, F.J.; Weerkamp, F.; Steyerberg, E.W.; van der Lei, J.; de Rijke, Y.B.; Oostenbrink, R. Validation of the Feverkidstool and procalcitonin for detecting serious bacterial infections in febrile children. Pediatr. Res. 2018, 83, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Nijman, R.G.; Vergouwe, Y.; Thompson, M.; van Veen, M.; van Meurs, A.H.; van der Lei, J.; Steyerberg, E.W.; Moll, H.A.; Oostenbrink, R. Clinical prediction model to aid emergency doctors managing febrile children at risk of serious bacterial infections: Diagnostic study. BMJ 2013, 346, f1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikema, A.P.; Horst-Kreft, D.; Boers, S.A.; Jansen, R.; Hiltemann, S.D.; de Koning, W.; Kraaij, R.; de Ridder, M.A.J.; van Houten, C.B.; Bont, L.J.; et al. Comparison of Illumina versus Nanopore 16S rRNA Gene Sequencing of the Human Nasal Microbiota. Genes 2020, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, I.; Wittouck, S.; Wuyts, S.; Oerlemans, E.F.M.; van den Broek, M.F.L.; Vandenheuvel, D.; Vanderveken, O.; Lebeer, S. Comparing the Healthy Nose and Nasopharynx Microbiota Reveals Continuity as Well as Niche-Specificity. Front. Microbiol. 2017, 8, 2372. [Google Scholar] [CrossRef] [Green Version]
- Gan, W.; Yang, F.; Tang, Y.; Zhou, D.; Qing, D.; Hu, J.; Liu, S.; Liu, F.; Meng, J. The difference in nasal bacterial microbiome diversity between chronic rhinosinusitis patients with polyps and a control population. Int. Forum Allergy Rhinol. 2019, 9, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Depner, M.; Ege, M.J.; Cox, M.J.; Dwyer, S.; Walker, A.W.; Birzele, L.T.; Genuneit, J.; Horak, E.; Braun-Fahrlander, C.; Danielewicz, H.; et al. Bacterial microbiota of the upper respiratory tract and childhood asthma. J. Allergy Clin. Immunol. 2017, 139, 826–834.e13. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoek, R.A.; Paats, M.S.; Pas, S.D.; Bakker, M.; Hoogsteden, H.C.; Boucher, C.A.; van der Eerden, M.M. Incidence of viral respiratory pathogens causing exacerbations in adult cystic fibrosis patients. Scand. J. Infect. Dis. 2013, 45, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Moesker, F.M.; van Kampen, J.J.A.; Aron, G.; Schutten, M.; van de Vijver, D.; Koopmans, M.P.G.; Osterhaus, A.D.M.E.; Fraaij, P.L.A. Diagnostic performance of influenza viruses and RSV rapid antigen detection tests in children in tertiary care. J. Clin. Virol. 2016, 79, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Spuesens, E.B.; Fraaij, P.L.; Visser, E.G.; Hoogenboezem, T.; Hop, W.C.; van Adrichem, L.N.; Weber, F.; Moll, H.A.; Broekman, B.; Berger, M.Y.; et al. Carriage of Mycoplasma pneumoniae in the upper respiratory tract of symptomatic and asymptomatic children: An observational study. PLoS Med. 2013, 10, e1001444. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.R.; Wieringa, J.; Koekkoek, S.M.; Visser, C.E.; Pajkrt, D.; Molenkamp, R.; de Jong, M.D.; Schinkel, J. Frequent detection of respiratory viruses without symptoms: Toward defining clinically relevant cutoff values. J. Clin. Microbiol. 2011, 49, 2631–2636. [Google Scholar] [CrossRef] [Green Version]
- Chonmaitree, T.; Alvarez-Fernandez, P.; Jennings, K.; Trujillo, R.; Marom, T.; Loeffelholz, M.J.; Miller, A.L.; McCormick, D.P.; Patel, J.A.; Pyles, R.B. Symptomatic and asymptomatic respiratory viral infections in the first year of life: Association with acute otitis media development. Clin. Infect. Dis. 2015, 60, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Self, W.H.; Williams, D.J.; Zhu, Y.; Ampofo, K.; Pavia, A.T.; Chappell, J.D.; Hymas, W.C.; Stockmann, C.; Bramley, A.M.; Schneider, E.; et al. Respiratory Viral Detection in Children and Adults: Comparing Asymptomatic Controls and Patients with Community-Acquired Pneumonia. J. Infect. Dis. 2016, 213, 584–591. [Google Scholar] [CrossRef] [Green Version]
- Spichak, T.V.; Yatsyshina, S.; Katosova, L.K.; Kim, S.S.; Korppi, M.O. Is the role of rhinoviruses as causative agents of pediatric community-acquired pneumonia over-estimated? Eur. J. Pediatr. 2016, 175, 1951–1958. [Google Scholar] [CrossRef]
- Singleton, R.J.; Bulkow, L.R.; Miernyk, K.; DeByle, C.; Pruitt, L.; Hummel, K.B.; Bruden, D.; Englund, J.A.; Anderson, L.J.; Lucher, L.; et al. Viral respiratory infections in hospitalized and community control children in Alaska. J. Med. Virol. 2010, 82, 1282–1290. [Google Scholar] [CrossRef]
- Sarna, M.; Lambert, S.B.; Sloots, T.P.; Whiley, D.M.; Alsaleh, A.; Mhango, L.; Bialasiewicz, S.; Wang, D.; Nissen, M.D.; Grimwood, K.; et al. Viruses causing lower respiratory symptoms in young children: Findings from the ORChID birth cohort. Thorax 2018, 73, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; McLean, K.; Campbell, H.; Nair, H. Aetiological role of common respiratory viruses in acute lower respiratory infections in children under five years: A systematic review and meta-analysis. J. Glob. Health 2015, 5, 010408. [Google Scholar] [CrossRef]
- Brand, H.K.; de Groot, R.; Galama, J.M.; Brouwer, M.L.; Teuwen, K.; Hermans, P.W.; Melchers, W.J.; Warris, A. Infection with multiple viruses is not associated with increased disease severity in children with bronchiolitis. Pediatr. Pulmonol. 2012, 47, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Toizumi, M.; Suzuki, M.; Nguyen, H.A.T.; Le, M.N.; Ariyoshi, K.; Moriuchi, H.; Hashizume, M.; Dang, D.A.; Yoshida, L.M. Viral Acute Respiratory Illnesses in Young Infants Increase the Risk of Respiratory Readmission. Pediatr. Infect. Dis. J. 2018, 37, 1217–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigl, J.A.; Puppe, W.; Grondahl, B.; Schmitt, H.J. Epidemiological investigation of nine respiratory pathogens in hospitalized children in Germany using multiplex reverse-transcriptase polymerase chain reaction. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Zhao, D.; Liu, J.; Wang, X.; Yang, K.; Xicheng, H.; Yang, L.; Wang, F. Multipathogen infections in hospitalized children with acute respiratory infections. Virol. J. 2009, 6, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishaupt, J.O.; van der Ploeg, T.; de Groot, R.; Versteegh, F.G.; Hartwig, N.G. Single- and multiple viral respiratory infections in children: Disease and management cannot be related to a specific pathogen. BMC Infect. Dis. 2017, 17, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotzen-Ostlund, M.; Eriksson, M.; Tiveljung Lindell, A.; Allander, T.; Zweygberg Wirgart, B.; Grillner, L. Children with multiple viral respiratory infections are older than those with single viruses. Acta Paediatr. 2014, 103, 100–104. [Google Scholar] [CrossRef]
- Jennings, L.C.; Anderson, T.P.; Werno, A.M.; Beynon, K.A.; Murdoch, D.R. Viral etiology of acute respiratory tract infections in children presenting to hospital: Role of polymerase chain reaction and demonstration of multiple infections. Pediatr. Infect. Dis. J. 2004, 23, 1003–1007. [Google Scholar] [CrossRef]
- Moesker, F.M.; van Kampen, J.J.; van Rossum, A.M.; de Hoog, M.; Koopmans, M.P.; Osterhaus, A.D.; Fraaij, P.L. Viruses as Sole Causative Agents of Severe Acute Respiratory Tract Infections in Children. PLoS ONE 2016, 11, e0150776. [Google Scholar] [CrossRef] [Green Version]
- Rhedin, S.; Lindstrand, A.; Hjelmgren, A.; Ryd-Rinder, M.; Ohrmalm, L.; Tolfvenstam, T.; Ortqvist, A.; Rotzen-Ostlund, M.; Zweygberg-Wirgart, B.; Henriques-Normark, B.; et al. Respiratory viruses associated with community-acquired pneumonia in children: Matched case-control study. Thorax 2015, 70, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.T.; Kuypers, J.; Wald, A.; Englund, J.A. Multiple versus single virus respiratory infections: Viral load and clinical disease severity in hospitalized children. Influenza Other Respir. Viruses 2012, 6, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.J.; Mansbach, J.M.; Wong, M.C.; Ajami, N.J.; Petrosino, J.F.; Camargo, C.A., Jr.; Hasegawa, K. Associations of Nasopharyngeal Metabolome and Microbiome with Severity among Infants with Bronchiolitis. A Multiomic Analysis. Am. J. Respir. Crit. Care Med. 2017, 196, 882–891. [Google Scholar] [CrossRef]
- Biesbroek, G.; Tsivtsivadze, E.; Sanders, E.A.; Montijn, R.; Veenhoven, R.H.; Keijser, B.J.; Bogaert, D. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. 2014, 190, 1283–1292. [Google Scholar] [CrossRef]
- Laufer, A.S.; Metlay, J.P.; Gent, J.F.; Fennie, K.P.; Kong, Y.; Pettigrew, M.M. Microbial communities of the upper respiratory tract and otitis media in children. mBio 2011, 2, e00245-10. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.L.; de Koff, E.M.; Bogaert, D. Characterising the respiratory microbiome. Eur. Respir. J. 2019, 53, 1801711. [Google Scholar] [CrossRef] [PubMed]
- Fodha, I.; Vabret, A.; Ghedira, L.; Seboui, H.; Chouchane, S.; Dewar, J.; Gueddiche, N.; Trabelsi, A.; Boujaafar, N.; Freymuth, F. Respiratory syncytial virus infections in hospitalized infants: Association between viral load, virus subgroup, and disease severity. J. Med. Virol. 2007, 79, 1951–1958. [Google Scholar] [CrossRef] [PubMed]
- Laham, F.R.; Mansbach, J.M.; Piedra, P.A.; Hasegawa, K.; Sullivan, A.F.; Espinola, J.A.; Camargo, C.A., Jr. Clinical Profiles of Respiratory Syncytial Virus Subtypes A and B Among Children Hospitalized with Bronchiolitis. Pediatr. Infect. Dis. J. 2017, 36, 808–810. [Google Scholar] [CrossRef]
- Shin, J.; Lee, S.; Go, M.J.; Lee, S.Y.; Kim, S.C.; Lee, C.H.; Cho, B.K. Analysis of the mouse gut microbiome using full-length 16S rRNA amplicon sequencing. Sci. Rep. 2016, 6, 29681. [Google Scholar] [CrossRef] [Green Version]
- Rang, F.J.; Kloosterman, W.P.; de Ridder, J. From squiggle to basepair: Computational approaches for improving nanopore sequencing read accuracy. Genome Biol. 2018, 19, 90. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, L.; Hasegawa, K.; Waris, M.; Ajami, N.J.; Petrosino, J.F.; Camargo, C.A., Jr.; Peltola, V. Early nasal microbiota and acute respiratory infections during the first years of life. Thorax 2019, 74, 592–599. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| General Characteristics | N or Median | % or IQR |
|---|---|---|
| Age in months * | 17 | 9–34 |
| Gender: male | 104 | 62.3 |
| Season of ED visit | ||
| Spring | 57 | 34.1 |
| Summer | 25 | 15 |
| Autumn | 41 | 24.6 |
| Winter | 44 | 26.3 |
| Clinical presentation | ||
| Duration of ED stay in minutes * (n = 141) | 147 | 112–193 |
| Temperature at ED visit | 38.8 | 1.0 |
| Duration of fever in days* (n = 166) | 2 | 1-3 |
| Tachypnea according to APLS (n = 165) | 145 | 87.9 |
| Tachycardia according to APLS | 127 | 76 |
| Saturation < 94% (n = 166) | 39 | 23.5 |
| Capillary refill time prolonged (n = 166) | 17 | 10.2 |
| Retractions (n = 166) | 105 | 63.3 |
| Ill appearance (n = 165) | 70 | 42.4 |
| Diagnosis | ||
| Pneumonia | 88 | 52.7 |
| Bronchiolitis | 22 | 13.2 |
| Upper RTI | 36 | 21.6 |
| Subglottic laryngitis | 2 | 1.2 |
| Viral induced wheeze | 18 | 10.8 |
| Other | 1 | 0.6 |
| Investigations | ||
| CRP in mg/L * (n = 152) | 35.5 | 15–76 |
| Other lab tests performed (n = 161) | 41 | 25.5 |
| PCR for viral diagnostics performed (n = 161) | 45 | 28 |
| Chest X-ray performed | 35 | 21 |
| Management and outcome | ||
| Hospitalization at first visit | 107 | 64.1 |
| Hospitalized at any time | 111 | 66.5 |
| Duration of hospitalization in days (n = 164) * | 1 | 0-3 |
| Strategy failure (n = 161) | 36 | 21.6 |
| Antibiotics at first visit | 85 | 50.9 |
| Antibiotics at any time (n = 162) | 100 | 61.7 |
| Risk groups based on Feverkidstool a (n = 146) | ||
| Low | 30 | 20.5 |
| Medium | 33 | 22.6 |
| High | 83 | 56.8 |
| Predicted risk of bacterial pneumonia by feverkidstool (n = 146) * | 12.3 | 5.6–29.0 |
| Oxygen therapy received at any time | 87 | 52.1 |
| N positive | ADV | boca | corona | entero | hMPV | infl A | infl B | Myco | parainfl | rhino | RSV | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N (%) | Count | Count | Count | Count | Count | Count | Count | Count | Count | Count | Count | |
| Adenovirus | 21 (13.3) | (2) | ||||||||||
| Bocavirus | 11 (7.0) | 4 | (1) | |||||||||
| Coronavirus | 10 (6.3) | 2 | 0 | (2) | ||||||||
| Enterovirus | 14 (8.9) | 2 | 0 | 1 | (6) | |||||||
| hMPV | 20 (12.7) | 1 | 1 | 1 | 1 | (15) | ||||||
| Influenza A | 5 (3.2) | 0 | 0 | 0 | 0 | 0 | (5) | |||||
| Influenza B | 2 (1.3) | 0 | 0 | 0 | 1 | 0 | 0 | (1) | ||||
| Mycoplasma | 2 (1.3) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | (0) | |||
| Parainfluenza | 14 (8.9) | 2 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | (8) | ||
| Rhinovirus | 63 (39.9) | 12 | 6 | 4 | 6 | 2 | 0 | 1 | 1 | 5 | (33) | |
| RSV | 39 (24.7) | 6 | 3 | 4 | 2 | 1 | 0 | 0 | 1 | 1 | 8 | (21) |
| Pathogen | Unadjusted OR | 95% CI | sig | Adjusted OR | 95% CI | sig | |
|---|---|---|---|---|---|---|---|
| Adenovirus | 0.46 | 0.182–1.161 | 0.1 | 0.388 | 0.137–1.095 | 0.074 | |
| Bocavirus | 2.69 | 0.561–12.907 | 0.216 | ||||
| Enterovirus | 0.076 | 0.016–0.353 | 0.001 | 0.078 | 0.015–0.395 | 0.002 | |
| Coronavirus | 1.37 | 0.340–5.519 | 0.685 | ||||
| Rhinovirus | 1.087 | 0.559–2.114 | 0.805 | ||||
| hMPV | 1.055 | 0.395–2.818 | 0.915 | ||||
| Influenza A | 0.842 | 0.136–5.193 | 0.853 | ||||
| Parainfluenza | 0.532 | 0.177–1.602 | 0.262 | ||||
| RSV | 1.368 | 0.631–2.968 | 0.427 | ||||
| Virusgroups | |||||||
| High virulence | 1.167 | 0.411–3.315 | 0.772 | ||||
| Low virulence | 0.899 | 0.310–2.607 | 0.845 | ||||
| Negative | ref | ||||||
| PCR positive for virus | 1.038 | 0.384–2.807 | 0.941 | ||||
| Microbiome profiles | |||||||
| Haemophilus | ref | ref | |||||
| Moraxella | 1.129 | 0.287–4.441 | 0.862 | 1.303 | 0.268–6.343 | 0.743 | |
| Streptococcus | 2.118 | 0.210–21.389 | 0.525 | 2.877 | 0.220–37.641 | 0.420 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoefnagels, I.; van de Maat, J.; van Kampen, J.J.A.; van Rossum, A.; Obihara, C.; Tramper-Stranders, G.A.; Heikema, A.P.; de Koning, W.; van Wermerskerken, A.-M.; Horst-Kreft, D.; et al. The Role of the Respiratory Microbiome and Viral Presence in Lower Respiratory Tract Infection Severity in the First Five Years of Life. Microorganisms 2021, 9, 1446. https://doi.org/10.3390/microorganisms9071446
Hoefnagels I, van de Maat J, van Kampen JJA, van Rossum A, Obihara C, Tramper-Stranders GA, Heikema AP, de Koning W, van Wermerskerken A-M, Horst-Kreft D, et al. The Role of the Respiratory Microbiome and Viral Presence in Lower Respiratory Tract Infection Severity in the First Five Years of Life. Microorganisms. 2021; 9(7):1446. https://doi.org/10.3390/microorganisms9071446
Chicago/Turabian StyleHoefnagels, Ivo, Josephine van de Maat, Jeroen J.A. van Kampen, Annemarie van Rossum, Charlie Obihara, Gerdien A. Tramper-Stranders, Astrid P. Heikema, Willem de Koning, Anne-Marie van Wermerskerken, Deborah Horst-Kreft, and et al. 2021. "The Role of the Respiratory Microbiome and Viral Presence in Lower Respiratory Tract Infection Severity in the First Five Years of Life" Microorganisms 9, no. 7: 1446. https://doi.org/10.3390/microorganisms9071446
APA StyleHoefnagels, I., van de Maat, J., van Kampen, J. J. A., van Rossum, A., Obihara, C., Tramper-Stranders, G. A., Heikema, A. P., de Koning, W., van Wermerskerken, A.-M., Horst-Kreft, D., Driessen, G. J. A., Punt, J., Smit, F. J., Stubbs, A., Noordzij, J. G., Hays, J. P., & Oostenbrink, R. (2021). The Role of the Respiratory Microbiome and Viral Presence in Lower Respiratory Tract Infection Severity in the First Five Years of Life. Microorganisms, 9(7), 1446. https://doi.org/10.3390/microorganisms9071446