
Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease

 , ,
, ,
Abstract
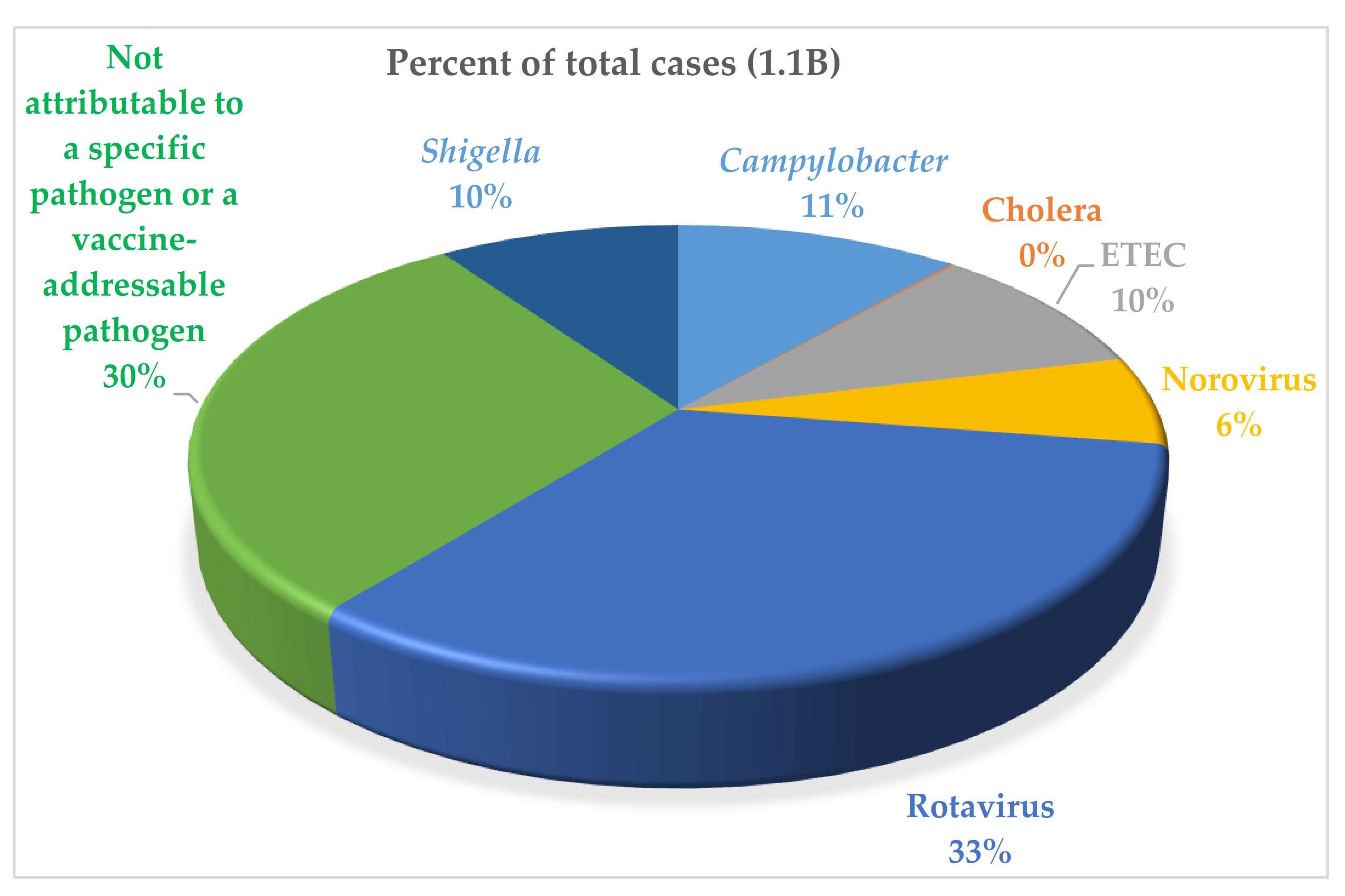
1. Introduction
2. Shigella, ETEC and Campylobacter: Targets for Vaccine Development
3. Shigella Component
3.1. Serotype-Dependent Candidates
3.2. Serotype-Independent Candidates
4. ETEC Component
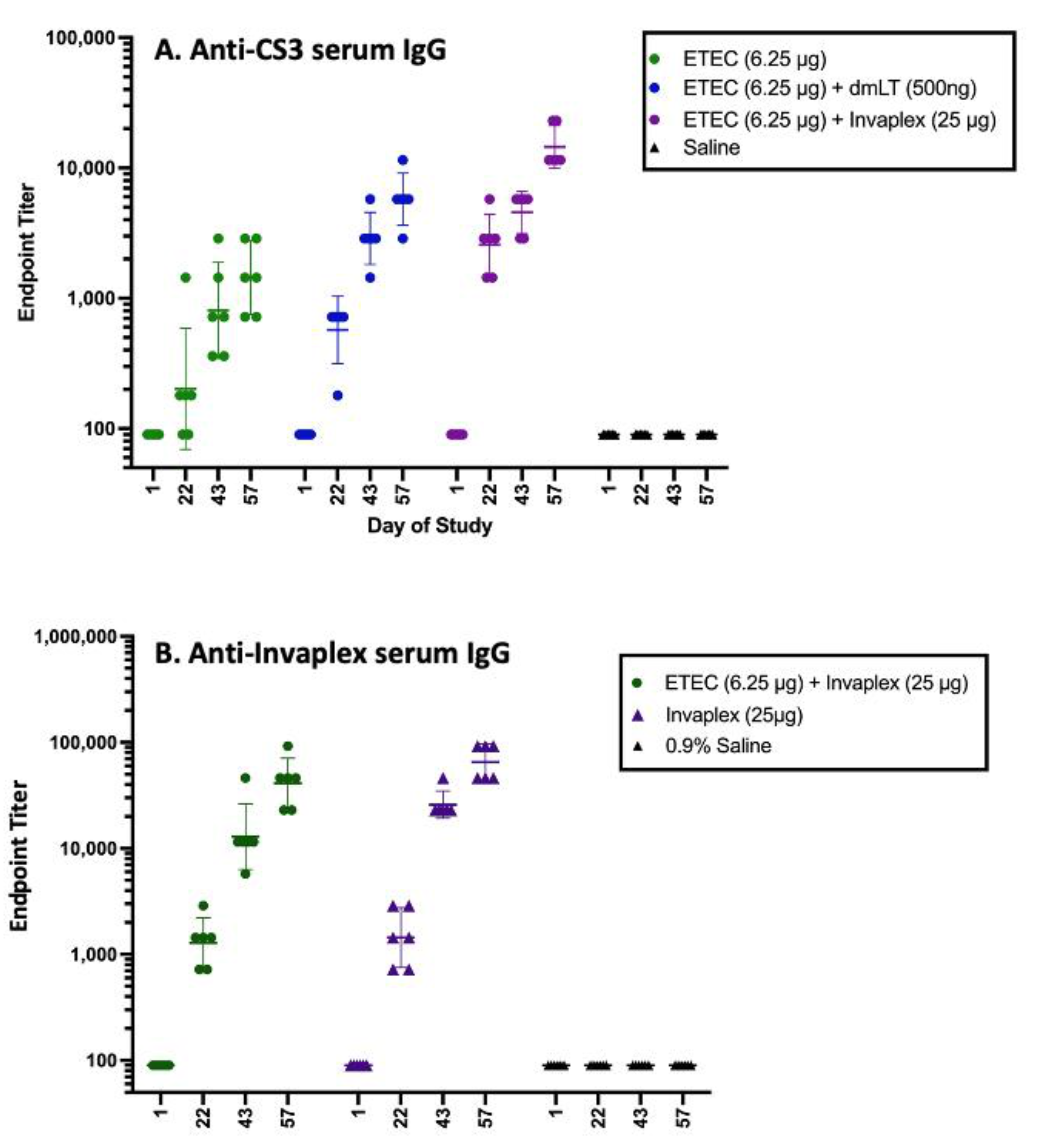
- Immunity against CFA/I-CS antigens could block pathogenesis by interrupting the adhesion of the pathogen to the intestinal epithelium. An innovative, subunit ETEC candidate uses recombinantly produced conserved subunits of some CFA/I-CS proteins. These are the fimbrial tip adhesin (FTA) proteins from ETEC, which can induce strong immune responses at systemic and mucosal sites when co-administered intramuscularly with dmLT [61]. Individual FTA antigens, which would comprise a complete quadrivalent vaccine, have each protected non-human primates [62,63] vaccinated intramuscularly and challenged orally. Two of the FTA antigens have undergone Phase 1 evaluations. The first is CfaE, which was found to safe and immunogenic when given intradermally with mLT, and reduced the incidence and severity of disease following challenge with a CFA/I-expressing ETEC [55]. The inclusion of the mLT adjuvant in the vaccine significantly improved the serum IgA and IgG response to CfaE, as well as the HAI antibody response to CfaE adhesin [55,64]. The safety and immunogenicity of the CssBA (CS6) antigen with or without dmLT given intramuscularly have also recently been evaluated in a Phase 1 trial. The CssBA antigen was found to be safe up to a dose of 45 ug (highest dose tested), and the serum and mucosal antibody responses to the antigen were significantly improved by the addition of dmLT, including increased levels of anti-CS6 α4β7 cells in the peripheral blood and anti-CS6 fecal antibody levels [65,66].
- Multiple epitope fusion antigens (MEFA) utilize CFA/I as a platform to express the dominant epitopes of other CFAs in a single protein, along with non-toxic LTA-LTB and ST [67,68]. MEFA vaccines stimulate neutralizing antibodies against the selected virulence antigens and piglets immunized with the MEFA–K88ac vaccine remained healthy following challenge [69,70]. Quantitative culture of the piglet ileum showed reduced colonization following immunization and K88 challenge. This candidate also protects rabbits against colonization by a human ETEC strain (ETEC strain B7A) [71] and could be cloned into a vector or delivered parenterally as a purified subunit.
- The application of new “Omics” technologies and other gene-based approaches also offer great promise for yielding new vaccine antigens from ETEC that may provide broad protection, as well as to facilitate combined vaccine strategies [72,73,74]. Pangenome analysis of multiple strains, combined with open-aperture ETEC immunoproteome interrogation of samples from both human volunteers [72] and naturally infected hosts, indicate that there are relatively few, highly conserved, strongly immunogenic, and ETEC pathovar specific antigens. These studies have highlighted two antigens, EatA and EtpA. EtpA is an extracellular adhesin of ETEC that promotes bacterial attachment and toxin delivery [73], acting as a bridge between bacteria and human A blood group expressed on the intestinal epithelia [74]. Similar to EatA, vaccination with EtpA effectively reduces the intestinal colonization of mice by ETEC. An additional antigen and virulence factor found in ETEC, as well as in other diarrheagenic and extra-intestinal E. coli, is YghJ (sometimes designated SslE). This protein antigen has shown protection in animal models, suggesting that it may have a vaccine potential [75,76,77,78]. The inclusion of these non-canonical antigens in ETEC vaccine candidates could complement or broaden the protection afforded by CF-based antigens.The majority of ETEC strains express a mucin-degrading serine protease autotransporter protein known as EatA [79], and EatA expression was recently shown to correlate strongly with symptomatic infection among young children in Bangladesh [80]. SepA, discovered as the major secreted protein present in culture supernatants of S. flexneri 5a [81], is an orthologue of EatA with which it shares a ~75% amino acid identity. Surveys of Shigella genomes reveal the presence of either SepA or EatA genes in each Shigella species [82], with SepA predominating in flexneri, and EatA more commonly represented in S. sonnei. The secreted EatA passenger domain is strongly immunogenic following ETEC infection of humans [83], and vaccination with this domain reduces colonization of mice after challenge with ETEC [72]. Whether vaccination with EatA or SepA can afford cross protection against ETEC and Shigella in humans remains to be determined.
- The virulence of ETEC strains is associated with LT and ST enterotoxins expressed in the small intestine. Toxoids based on the LT or ST antigens may be found to contribute to the effectiveness of ETEC vaccines. A transdermal LT patch, no longer in development, was used to show protection against LT-only producing strains in a Phase 3 trial [84]. dmLT has also been shown to induce anti-LT antibody responses in human volunteers and to induce better LT toxin neutralization antibody responses than when B-subunit toxoid preparations have been used [85]. Field and controlled human infection model (CHIM) study data indicate that inducing anti-LT immunity can be an immune marker for reduced risk of ETEC illness, particularly as a result of the ETEC strains expressing only the LT toxin [84,86]. CHIM and field studies also indicate that strong immune responses to LT can help to reduce the severity of ETEC illness when it occurs [84,87]. LT appears to promote ETEC colonization by changing the surface architecture of the intestinal epithelia to favor the pathogen attachment [88], and anti-LT immunity can cooperatively impact effective ETEC small intestinal colonization when combined with anti-adhesin approaches [73]. Moreover, LT has been shown to accentuate the enterotoxic effects of the ST toxin [89], suggesting that anti-LT immunity could be beneficial in mitigating the impact of both toxins. The importance of ST in a vaccine is less well established, but passive immunization studies in piglets have shown protection against disease [70]. Recent work has shown that the ST molecule can be detoxified without losing neutralizing properties, be made immunogenic by linking to a protein, and avoid cross reaction with human guanylin-uroguanylin [90,91,92]. In addition, novel studies of ST toxin secretion and delivery have shown that the STH propeptide is secreted by ETEC, potentially providing additional epitopes for inducing toxin neutralizing antibodies [93]. These studies also showed that EtpA adhesin plays an important role in ST toxin delivery to intestinal epithelial cells, suggesting that antibodies against this potential vaccine antigen could reduce ST toxicity [93].
5. Campylobacter Component
6. Multi-Pathogen Strategies
6.1. Multi-Pathogen Vaccines for Oral Administration
- The guaBA Shigella−ETEC hybrid 1208-122 should provide effective coverage against both Shigella and ETEC, relying on antigens that have been well established to protect [57]. A prototype of this candidate, S. flexneri 2a expressing CFA/I and LT, protected orally challenged mice against ETEC and Shigella [58], and may enter Phase 1 evaluation in 2021 (https://clinicaltrials.gov/ct2/show/NCT04634513, accessed on 1 May 2021). This approach demonstrates the use of expression of heterologous antigens to broaden coverage, which could also be applied to conserved Campylobacter antigens.
- ShigETEC, in contrast with the hybrid, relies entirely on little studied but promising conserved surface proteins minus those of the deleted T3SS to protect against Shigella [49,50]. The strain contains a RfbF deletion that renders it rough, i.e., not expressing the serotype-determining O-PS. ETEC coverage is anticipated from homology existing between Shigella and ETEC [59,60], as well as the LT/ST chimeric toxoid expressed [51]. As stated above, the extent that the homology present in Shigella will contribute to protection against major ETEC strains remains to be determined.
- A third platform at an earlier stage of development than the previous two is the Shigella truncated mutant (STM) comprised of inactivated Shigella mutants with O-polysaccharide chains truncated to one repeating unit in length [48]. This, like the ShigETEC, enhances the immunological accessibility of conserved and protective Shigella outer membrane proteins such as PSSP-1 [46], and may also be a benefit to vectored antigens. STM also express additional conserved proteins not normally masked by O-polysaccharide chains, such as the Ipa proteins (deleted in the ShigETEC approach), which could also contribute to broader serotype-independent immunity [36,37]. STM should also benefit from homology to ETEC, but it remains to be seen whether it may also be necessary to engineer the expression of some additional ETEC antigens such as EtpA or CS6 into the STM for optimal coverage. For instance, data emerging from the analysis of more than 1500 phylogenetically and geographically diverse isolates of ETEC suggest that a combination of CS6 and EtpA, which is more frequently absent in CS6-expressing strains, would afford coverage for more than 80% of ETEC [65,72]. The STM is the only candidate currently planned to be engineered to express the conserved Campylobacter heptasaccharide [104].
- A recombinant live oral ETEC vaccine (Ty21a-ETEC) composed of Ty21a, the oral typhoid vaccine, expressing both heat-labile (LT) and heat stable enterotoxin (STa) and seven adhesins (CFA/I, CFA/II (CS1-CS3), and CFA/IV (CS4-CS6)) that facilitate the colonization of host intestines and binds GM1was constructed [110]. The seven adhesins comprise a multi-epitope fusion antigen (MEFA) that has shown to have a broad spectrum anti-adhesin activity. The intranasal (IN) immunization of BALB/c mice, which mimicked a mucosal/oral route of immunization, induced antibodies against LTB and MEFA that blocked binding to GM1, showing the induction of an anti-toxin activity. Further antibodies induced by IN immunization induced antibodies that blocked the adhesion of ETEC to Caco-2 cells. The typhoid vaccine has also served as a vector for the major Shigella O-PS [20,21].
6.2. Multi-Pathogen Vaccines for Parenteral Administration
7. Considerations to Optimize the Immunological and Practical Impact of Vaccine Candidates to Provide an Effective Multi-Pathogen Vaccine Strategy
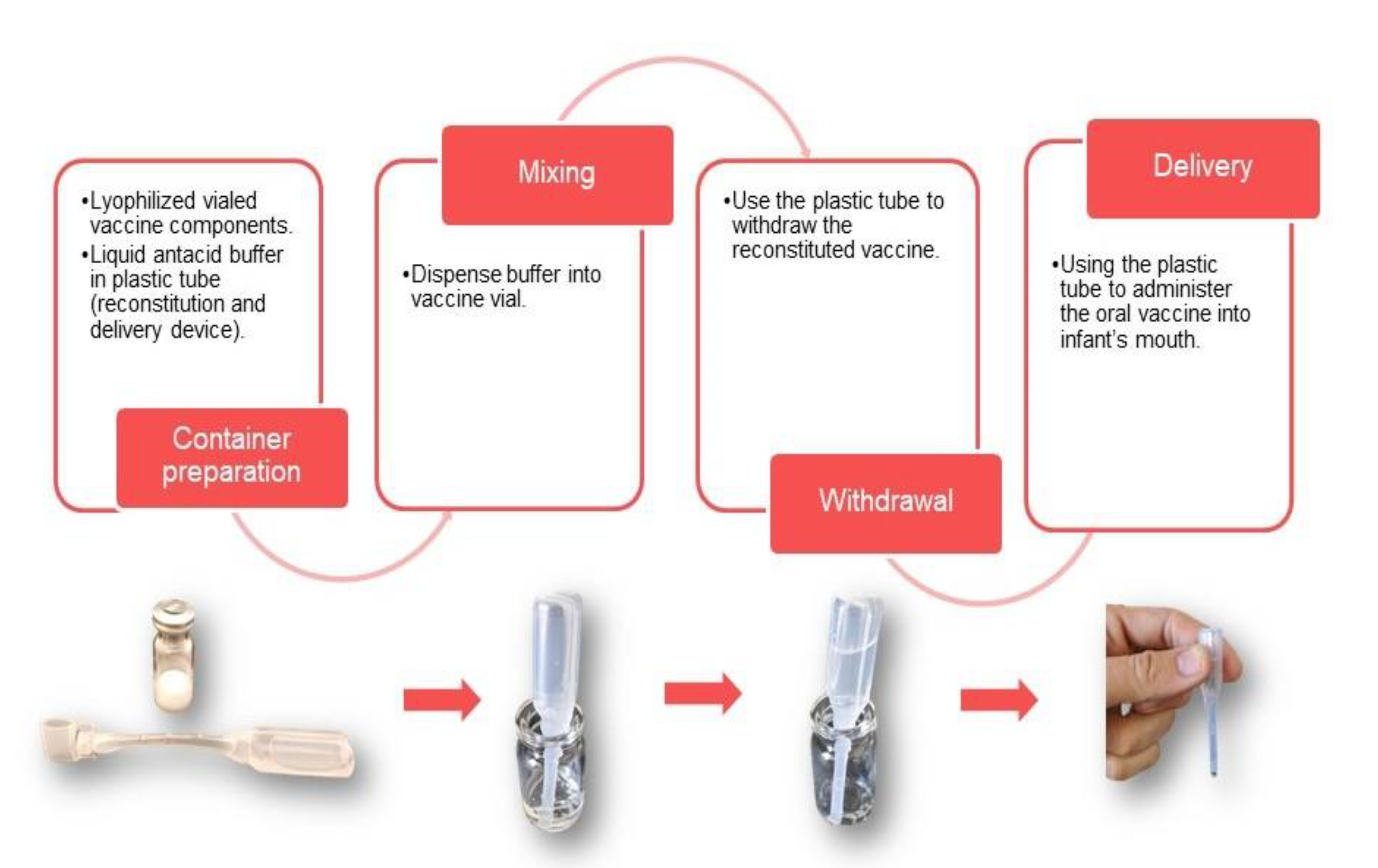
7.1. Pediatric Presentation for Oral Vaccines
7.2. dmLT Adjuvant
7.3. Improved Tolerability and Immunogenicity of Oral Inactivated Whole Cell Vaccines for Infants
7.4. Dosing Schedule
7.5. Animal Models for Protection
7.6. CHIMs for Enteric Vaccine Development
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Disclaimer
Copyright Statement
References
- Formal, S.B.; LaBrec, E.H.; Palmer, A.; Falkow, S. Protection of Monkeys against Experimental Shigellosis with Attenuated Vaccines. J. Bacteriol. 1965, 9, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L.; et al. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef]
- Platts-Mills, J.A.; Liu, J.; Rogawski, E.T.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Khan, S.S.; Praharaj, I.; Murei, A.; Nshama, R.; et al. Use of quantitative molecular diagnostic methods to assess the aetiology, burden, and clinical characteristics of diarrhoea in children in low-resource settings: A reanalysis of the MAL-ED cohort study. Lancet Glob. Health 2018, 6, e1309–e1318. [Google Scholar] [CrossRef]
- GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef]
- Khalil, I.A. (Institute for Health Metrics, Seattle, WA, USA). Personal Communication, 2021.
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef]
- Nataro, J.P.; Guerrant, R.L. Chronic consequences on human health induced by microbial pathogens: Growth faltering among children in developing countries. Vaccine 2017, 14, 6807–6812. [Google Scholar] [CrossRef]
- Rogawski, E.T.; Guerrant, R.L. The Burden of Enteropathy and “Subclinical” Infections. Pediatr. Clin. N. Am. 2017, 64, 815–836. [Google Scholar] [CrossRef]
- Anderson, J.D.; Bagamian, K.H.; Muhib, F.; Amaya, M.P.; Laytner, L.A.; Wierzba, T.; Rheingans, R. Burden of enterotoxigenic Escherichia coli and Shigella non-fatal diarrhoeal infections in 79 low-income and lower middle-income countries: A modelling analysis. Lancet Glob. Health 2019, 7, e321–e330. [Google Scholar] [CrossRef]
- Malarski, M.; Hasso-Agopsowicz, M.; Soble, A.; Mok, W.; Mathewson, S.; Vekemans, J. Vaccine impact on antimicrobial resistance to inform Gavi, the Vaccine Alliance’s 2018 Vaccine Investment Strategy: Report from an expert survey. F1000Research 2019, 8, 1685. [Google Scholar] [CrossRef]
- Antibiotic Resistance Threats in the United States, 2019; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [CrossRef]
- Camacho, A.I.; Irache, J.M.; Gamazo, C. Recent progress towards development of a Shigella vaccine. Expert Rev. Vaccines 2013, 12, 43–55. [Google Scholar] [CrossRef]
- Walker, R.I. An assessment of enterotoxigenic Escherichia coli and Shigella vaccine candidates for infants and children. Vaccine 2015, 33, 954–965. [Google Scholar] [CrossRef]
- Ndungo, E.; Randall, A.; Hazen, T.H.; Kania, D.A.; Trappl-Kimmons, K.; Liang, X.; Barry, E.M.; Kotloff, K.L.; Chakraborty, S.; Mani, S.; et al. A Novel Shigella Proteome Microarray Discriminates Targets of Human Antibody Reactivity following Oral Vaccination and Experimental Challenge. mSphere 2018, 3, e00260–e00318. [Google Scholar] [CrossRef]
- Clarkson, K.A.; Frenck, R.W., Jr.; Dickey, M.; Suvarnapunya, A.E.; Chandrasekaran, L.; Weerts, H.P.; Heaney, C.D.; McNeal, M.; Detizio, K.; Parker, S.; et al. Immune Response Characterization after Controlled Infection with Lyophilized Shigella sonnei 53G. mSphere 2020, 5, e00988–e01019. [Google Scholar] [CrossRef]
- Randall, A. (Antigen Discovery Inc., Irvine, CA, USA). Personal Communication, 2021.
- Venkatesan, M.M.; Ranallo, R.T. Live-attenuated Shigella vaccines. Expert Rev. Vaccines 2006, 5, 669–686. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Simon, J.K.; Pasetti, M.F.; Sztein, M.B.; Wooden, S.L.; Livio, S.; Nataro, J.P.; Blackwelder, W.C.; Barry, E.M.; Picking, W.; et al. Safety and Immunogenicity of CVD 1208S, a Live, OralΔguaBA Δsen Δset Shigella flexneri2a Vaccine Grown on Animal-Free Media. Hum. Vaccines 2007, 3, 268–275. [Google Scholar] [CrossRef]
- Dharmasena, M.N.; Osorio, M.; Takeda, K.; Stibitz, S.; Kopecko, D.J. Stable Chromosomal Expression of Shigella flexneri 2a and 3a O-Antigens in the Live Salmonella Oral Vaccine Vector Ty21a. Clin. Vaccine Immunol. 2017, 24, e00181-17. [Google Scholar] [CrossRef]
- Wu, Y.; Chakraborty, S.; Li, M.; Wai, T.T.; Hoffman, S.L.; Sim, B.K. Development of a live attenuated bivalent oral vaccine against Shigella sonnei shigellosis and typhoid fever. J. Infect. Dis. 2017, 215, 259–268. [Google Scholar] [CrossRef]
- Kaminski, R.W.; Oaks, E.V. Inactivated and subunit vaccines to prevent shigellosis. Expert Rev. Vaccines 2009, 8, 1693–1704. [Google Scholar] [CrossRef]
- McKenzie, R.; Walker, R.I.; Nabors, G.S.; Van De Verg, L.L.; Carpenter, C.; Gomes, G.; Forbes, E.; Tian, J.H.; Yang, H.H.; Pace, J.L.; et al. Safety and immunogenicity of an oral, inactivated, whole-cell vaccine for Shigella sonnei: Preclinical studies and a Phase I trial. Vaccine 2006, 24, 3735–3745. [Google Scholar] [CrossRef]
- Chakraborty, S.; Harro, C.; DeNearing, B.; Bream, J.; Bauers, N.; Dally, L.; Flores, J.; Van De Verg, L.; Sack, D.A.; Walker, R. Evaluation of the Safety, Tolerability, and Immunogenicity of an Oral, Inactivated Whole-Cell Shigella flexneri 2a Vaccine in Healthy Adult Subjects. Clin. Vaccine Immunol. 2016, 23, 315–325. [Google Scholar] [CrossRef]
- Passwell, J.H.; Ashkenazi, S.; Banet-Levi, Y.; Ramon-Saraf, R.; Farzam, N.; Lerner-Geva, L.; Even-Nir, H.; Yerushalmi, B.; Chu, C.; Shiloach, J.; et al. Age-related efficacy of Shigella O-specific polysaccharide conjugates in 1–4-year-old Israeli children. Vaccine 2010, 28, 2231–2235. [Google Scholar] [CrossRef]
- Ravenscroft, N.; Braun, M.; Schneider, J.; Dreyer, A.M.; Wetter, M.; Haeuptle, M.A.; Kemmler, S.; Steffen, M.; Sirena, D.; Herwig, S.; et al. Characterization and immunogenicity of a Shigella flexneri 2a O-antigen bioconjugate vaccine candidate. Glycobiology 2019, 29, 669–680. [Google Scholar] [CrossRef]
- Alaimo, C. Development of a Shigella multivalent bioconjugate vaccine: Toward a phase 1/2 in Kenyan infants. In Proceedings of the 10th International Conference on Vaccine for Enteric Diseases, Lausanne, Switzerland, 16–18 October 2019. [Google Scholar]
- Talaat, K.R.; Alaimo, C.; Bourgeois, A.L.; Kaminski, R.W.; Dreyer, A.; Porter, C.K.; Chakraborty, S.; Clarkson, K.A.; Brubaker, J.; Elwood, D.; et al. Flexyn2a, a candidate bioconjugate vaccine against Shigella flexneri 2a induces protective immune response in a controlled human infection model. In Proceedings of the 9th International Conference on Vaccine for Enteric Diseases, Albufeira, Portugal, 9–11 October 2017. [Google Scholar]
- Barel, L.A.; Mulard, L.A. Classical and novel strategies to develop a Shigella glycoconjugate vaccine: From concept to efficacy in human. Hum. Vaccines Immunother. 2019, 15, 1338–1356. [Google Scholar] [CrossRef]
- Cohen, D.; Meron-Sudai, S.; Bialik, A.; Asato, V.; Goren, S.; Ariel-Cohen, O.; Reizis, A.; Hochberg, A.; Ashkenazi, S. Serum IgG antibodies toShigellalipopolysaccharide antigens—A correlate of protection against shigellosis. Hum. Vaccines Immunother. 2019, 15, 1401–1408. [Google Scholar] [CrossRef]
- Hartman, A.B.; Van de Verg, L.L.; Collins, H.H., Jr.; Tang, D.B.; Bendiuk, N.O.; Taylor, D.N.; Powell, C.J. Local immune response and protection in the guinea pig keratoconjunctivitis model following immunization with Shigella vaccines. Infect. Immun. 1994, 62, 412–420. [Google Scholar] [CrossRef]
- Obiero, C.W.; Ndiaye, A.G.W.; Sciré, A.S.; Kaunyangi, B.M.; Marchetti, E.; Gone, A.M.; Schütte, L.D.; Riccucci, D.; Auerbach, J.; Saul, A. A Phase 2a Randomized Study to Evaluate the Safety and Immunogenicity of the 1790GAHB Generalized Modules for Membrane Antigen Vaccine against Shigella sonnei Administered Intramuscularly to Adults from a Shigellosis-Endemic Country. Front. Immunol. 2017, 8, 1884. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Giorgina Vitali, C.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef]
- Norton, E.B.; Bauer, D.L.; Weldon, W.C.; Oberste, M.S.; Lawson, L.B.; Clements, J.D. The novel adjuvant dmLT promotes dose sparing, mucosal immunity and longevity of antibody responses to the inactivated polio vaccine in a murine model. Vaccine 2015, 33, 1909–1915. [Google Scholar] [CrossRef]
- Svennerholm, A.M.; Lundgren, A.; Akhtar, M.; Walker, R.; Bourgeois, A.L.; Qadri, F. Determination of mucosal immune responses against an oral ETEC vaccine in infants. In Proceedings of the 10th International Conference on Vaccines for Enteric Diseases (VED 2019), Lausanne, Switzerland, 16–18 October 2019. [Google Scholar]
- Martinez-Becerra, F.J.; Kissmann, J.M.; Diaz-McNair, J.; Choudhari, S.P.; Quick, A.M.; Mellado-Sanchez, G.; Clements, J.D.; Pasetti, M.F.; Picking, W.L. Broadly Protective Shigella Vaccine Based on Type III Secretion Apparatus Proteins. Infect. Immun. 2012, 80, 1222–1231. [Google Scholar] [CrossRef]
- Martinez-Becerra, F.J.; Chen, X.; Dickenson, N.E.; Choudhari, S.P.; Harrison, K.; Clements, J.D.; Picking, W.D.; Van De Verg, L.L.; Walker, R.I.; Picking, W.L. Characterization of a Novel Fusion Protein from IpaB and IpaD of Shigella spp. and Its Potential as a Pan-Shigella Vaccine. Infect. Immun. 2013, 81, 4470–4477. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.; Clements, J.D.; Picking, W.; Freytag, L. Protective vaccine against shigellosis composed of a Shigella IpaB-IpaD fusion protein (DBF) in combination with the adjuvant dmLT. In Proceedings of the Vaccines for Enteric Diseases Conference, Albufeira, Portugal, 9–11 October 2017. [Google Scholar]
- Chitradevi, S.T.S.; Kaur, G.; Sivaramakrishna, U.; Singh, D.; Bansal, A. Development of recombinant vaccine candidate molecule against Shigella infection. Vaccine 2016, 34, 5376–5383. [Google Scholar] [CrossRef] [PubMed]
- Oaks, E.V.; Turbyfill, K.R. Development and evaluation of a Shigella flexneri 2a and S. sonnei bivalent invasin complex 7. (Invaplex) vaccine. Vaccine 2006, 24, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Turbyfill, K.R.; Hartman, A.B.; Oaks, E.V. Isolation and Characterization of a Shigella flexneri Invasin Complex Subunit Vaccine. Infect. Immun. 2000, 68, 6624–6632. [Google Scholar] [CrossRef]
- Tribble, D.; Kaminski, R.; Cantrell, J.; Nelson, M.; Porter, C.; Baqar, S.; Williams, C.; Arora, R.; Saunders, J.; Ananthakrishan, M.; et al. Safety and immunogenicity of a Shigella flexneri 2a Invaplex 50 intranasal vaccine in adult volunteers. Vaccine 2010, 28, 6076–6085. [Google Scholar] [CrossRef]
- Riddle, M.S.; Kaminski, R.W.; Williams, C.; Porter, C.; Baqar, S.; Kordis, A.; Gilliland, T.; Lapa, J.; Coughlin, M.; Soltis, C.; et al. Safety and immunogenicity of an intranasal Shigella flexneri 2a Invaplex 50 vaccine. Vaccine 2011, 26, 7009–7019. [Google Scholar] [CrossRef]
- Turbyfill, K.R.; Clarkson, K.A.; Vortherms, A.R.; Oaks, E.V.; Kaminski, R.W. Assembly, Biochemical Characterization, Immunogenicity, Adjuvanticity, and Efficacy of Shigella Artificial Invaplex. mSphere 2018, 3, e00583–e00617. [Google Scholar] [CrossRef]
- Kaminski, R. Protection against Shigellosis with InvaplexAR-Detox administered intramuscularly; Walter Reed Army Institute of Research: Silver Spring, MD, USA, Unpublished; manuscript in preparation.
- Kim, J.O.; Rho, S.; Kim, S.H.; Kim, H.; Song, H.J.; Kim, E.J.; Kim, R.Y.; Kim, E.H.; Sinha, A.; Dey, A.; et al. Shigella Outer Membrane Protein PSSP-1 Is Broadly Protective against Shigella Infection. Clin. Vaccine Immunol. 2015, 22, 381–388. [Google Scholar] [CrossRef]
- Goldberg, M.B.; Bârzu, O.; Parsot, C.; Sansonetti, P.J. Unipolar localization and ATPase activity of IcsA, a Shigella flexneri protein involved in intracellular movement. J. Bacteriol. 1993, 175, 2189–2196. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.H.; Kim, H.; Rho, S.; Shin, Y.K.; Song, M.; Walker, R.; Czerkinsky, C.; Kim, D.W.; Kim, J.O. Cross-Protective Shigella Whole-Cell Vaccine with a Truncated O-Polysaccharide Chain. Front. Microbiol. 2018, 9, 2609. [Google Scholar] [CrossRef]
- Krause, P.; Harutyunyan, S.; Neuhauser, I.; Aichinger, M.; Szijarto, V.; Nagy, G.; Nagy, E.; Henics, T. A Live Attenuated Vaccine against Shigella and ETEC: Characteristics and Potency of the ShigETEC Prototype Strain. In Proceedings of the VASE 2016 Conference, Washington, DC, USA, 28–30 June 2016. [Google Scholar]
- Giraldi, P.; Harutyunyan, S.; Neuhauser, I.; Szijarto, V.; Nagy, G.; Nagy, E.; Henics, T. Immune assays to evaluate ShigETEC, a live, attenuated combination vaccine against shigellosis and ETEC diarrhea. In Proceedings of the 2018 VASE Conference, Mexico City, Mexico, 11–14 June 2018. [Google Scholar]
- Harutyunyan, S.; Neuhauser, I.; Mayer, A.; Aichinger, M.; Szijártó, V.; Nagy, G.; Nagy, E.; Girardi, P.; Malinoski, F.J.; Henics, T. Characterization of ShigETEC, a Novel Live Attenuated Combined Vaccine against Shigellae and ETEC. Vaccines 2020, 8, 689. [Google Scholar] [CrossRef]
- Harro, C.; Bourgeois, A.L.; Sack, D.; Walter, R.; DeNearing, B.; Brubaker, J.; Maier, N.; Fix, A.; Dally, L.; Chakraborty, S.; et al. Live attenuated enterotoxigenic Escherichia coli (ETEC) vaccine with dmLT adjuvant protects human volunteers against virulent experimental ETEC challenge. Vaccine 2019, 37, 1978–1986. [Google Scholar] [CrossRef]
- Akhtar, M.; Chowdhury, M.I.; Bhuiyan, T.R.; Kaim, J.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.A.; Khanam, F.; Begum, Y.A. Evaluation of the safety and immunogenicity of the oral inactivated multivalent enterotoxigenic Escherichia coli vaccine ETVAX in Bangladeshi adults in a double-blind, randomized, placebo-controlled Phase I trial using electrochemiluminescence and ELISA assays for immunogenicity analyses. Vaccine 2019, 37, 5645–5656. [Google Scholar] [CrossRef]
- Qadri, F.; Akhtar, M.; Bhuiyan, T.R.; Chowdhury, M.I.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.A.; Khanam, F.; Lundgren, A.; et al. Safety and immunogenicity of the oral, inactivated, enterotoxigenic Escherichia coli vaccine ETVAX in Bangladeshi children and infants: A double-blind, randomised, placebo-controlled phase 1/2 trial. Lancet Infect. Dis. 2019, 20, 208–219. [Google Scholar] [CrossRef]
- WHO. Product Development for Vaccines Advisory Committee (PDVAC) Meeting: Executive Summary. Geneva, Switzerland, 26–27 June 2018. Available online: https://www.who.int/immunization/research/meetings_workshops/pdvac_june18/en/ (accessed on 22 June 2021).
- Lundgren, A.; Bourgeois, L.; Carlin, N.; Clements, J.; Gustafsson, B.; Hartford, M.; Holmgren, J.; Petzold, M.; Walker, R.; Svennerholm, A.M. Safety and immunogenicity of an improved oral inactivated multivalent enterotoxigenic Escherichia coli (ETEC) vaccine administered alone and together with dmLT adjuvant in a double-blind, randomized, placebo-controlled Phase I study. Vaccine 2014, 32, 7077–7084. [Google Scholar] [CrossRef]
- Barry, E.M.; Levine, M.M. A tale of two bacterial enteropathogens and one multivalent vaccine. Cell. Microbiol. 2019, 21, e13067. [Google Scholar] [CrossRef]
- Medeiros, P.H.Q.S.; Bolick, D.T.; Ledwaba, S.E.; Kolling, G.L.; Costa, D.V.S.; Oriá, R.B.; Lima, A.A.M.; Barry, E.M.; Guerrant, R.L. A bivalent vaccine confers immunogenicity and protection against Shigella flexneri and enterotoxigenic Escherichia coli infections in mice. Vaccines 2020, 5, 30. [Google Scholar] [CrossRef]
- Brenner, D.J.; Fanning, G.R.; Steigerwalt, A.G.; Orskov, I.; Orskov, F. Poly- nucleotide sequence relatedness among three groups of pathogenic Escherichia coli strains. Infect. Immun. 1972, 6, 308–315. [Google Scholar] [CrossRef]
- Devanga, N.K.; Ragupathi, D.P.; Sethuvel, M.; Inbanathan, F.Y.; Veeraraghavan, B. Accurate differentiation of Escherichia coli and Shigella serogroups: Challenges and strategies. New Microbe New Infect. 2018, 21, 58–62. [Google Scholar] [CrossRef]
- Poole, S.T., Jr.; Maciel, M.; Dinadayala, P.; Dori, K.E.; McVeigh, A.L.; Liu, Y.; Barry, E.; Grassel, C.; Prouty, M.G.; Renauld-Mongénie, G.; et al. Biochemical and Immunological Evaluation of Recombinant CS6-Derived Subunit Enterotoxigenic Escherichia coli Vaccine Candidates. Infect. Immun. 2019, 87, e00788–e00818. [Google Scholar] [CrossRef]
- Joseph, S.; Ramakrishnan, A.; Nunez, G.; Royal, J.; Maciel, M., Jr.; Regeimbal, J.; McCoy, A.; Savarino, S.; Renauld-Mongenie, G.; Heinrichs, J.; et al. Evaluation of Class 5a fimbrial adhesin-pilin fusion vaccines in Aotus nancymaae. In Proceedings of the VASE 2018, Mexico City, Mexico, 14 June 2018. [Google Scholar]
- Stoppato, M.; Gaspar, C.; Regeimbal, J.; Nunez, R.G.; Giuntini, S.; Schiller, Z.A.; Gawron, M.A.; Pondish, J.R.; Martin, J.C., 3rd; Schneider, M.I.; et al. Oral administration of an anti-CfaE secretory IgA antibody protects against Enterotoxigenic Escherichia coli diarrheal disease in a nonhuman primate model. Vaccine 2020, 38, 2333–2339. [Google Scholar] [CrossRef]
- Wierzba, T.; Orr, M.; Sturtevant, E.; Lutsch, C.; Bourgis, A.; Kuehn, C.; Giller, N.; Danve-Chery, E.; Prouty, M.; Mark Riddle, M.; et al. Advancing an ETEC vaccine for global health: A fimbrial tip adhesin approach. In Proceedings of the 9th International Conference on Vaccine for Enteric Diseases, Albufeira, Portugal, 9–11 October 2017. [Google Scholar]
- Lee, T.K.; Porter, C.K.; Gutierrez, R.L. A phase 1 dose escalating study of a prototype CS6 subunit vaccine with modified heat-labile enterotoxin from enterotoxigenic Escherichia coli (ETEC). In Proceedings of the E. coli and the Mucosal Immune System (ECMIS) Symposium, Ghent, Belgium, 2–5 June 2019. [Google Scholar]
- Maciel, M.; Trop, S.; Kim, A.; Ward, E.; Villar, Z.; Lee, T.K.; Jaep, K.; Porter, C.; Poole, S.; Prouty, M.G. Serological and α4β7+ antibody-secreting cell responses after intramuscular immunization with CssBA, a CS6-subunit based enterotoxigenic E. coli vaccine candidate and LT(R192G/L211A) as adjuvant. In Proceedings of the 10th International Conference on Vaccines for Enteric Diseases (VED 2019), Lausanne, Switzerland, 16–18 October 2019. [Google Scholar]
- Seo, H.; Lu, T.; Mani, S.; Bourgeois, A.L.; Walker, R.; Sack, D.A.; Zhang, W. Adjuvant effect of enterotoxigenic Escherichia coli (ETEC) double-mutant heat-labile toxin (dmLT) on systemic immunogenicity induced by the CFA/I/II/IV MEFA ETEC vaccine: Dose-related enhancement of antibody responses to seven ETEC adhesins (CFA/I, CS1-CS6). Hum. Vaccines Immunother. 2019, 1–7. [Google Scholar] [CrossRef]
- Seo, H.; Nandre, R.M.; Nietfeld, J.; Chen, Z.; Duan, Q.; Zhang, W. Antibodies induced adherence in vitro by enterotoxigenic Escherichia coli (ETEC) adhesin major structural subunit and minor tip adhesin subunit equivalently inhibit bacteria. PLoS ONE 2019, 14, e0216076. [Google Scholar] [CrossRef]
- Nandre, R.; Ruan, X.; Lu, T.; Duan, Q.; Sack, D.; Zhang, W. Enterotoxigenic Escherichia coli Adhesin-Toxoid Multiepitope Fusion Antigen CFA/I/II/IV-3xSTa N12S -mnLT G192G/L211A -Derived Antibodies Inhibit Adherence of Seven Adhesins, Neutralize Enterotoxicity of LT and STa Toxins, and Protect Piglets against Diarrhea. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef]
- Nandre, R.; Duan, Q.; Wang, Y.; Zhang, W. Passive antibodies derived from intramuscularly immunized toxoid fusion xSTaN12S-dmLT protect against Sta+ enterotoxigenic Escherichia coli (ETEC) diarrhea in a pig model. Vaccine 2017, 35, 552–556. [Google Scholar] [CrossRef]
- Jones, R.M.; Voeglein, J.B.; Connor, S.; Zhang, W.; Sack, D.A. A multi-epitope fusion antigen (MEFA) candidate vaccine for ETEC is protective in rabbit model. In Proceedings of the 52nd Joint Conference, United States–Japan Cooperative Medical Science Program Cholera Panel, Hanoi, Vietnam, 26 February–1 March 2019. [Google Scholar]
- Luo, Q.; Wickers, T.J.; Fleckenstein, J.M. Immunogenicity and Protective Efficacy against Enterotoxigenic Escherichia coli Colonization following Intradermal, Sublingual, or Oral Vaccination with EtpA Adhesin. Clin. Vaccine Immunol. 2016, 23, 628636. [Google Scholar] [CrossRef]
- Roy, K.; Hamilton, D.J.; Fleckenstein, J.M. Cooperative Role of Antibodies against Heat-Labile Toxin and the EtpA Adhesin in Preventing Toxin Delivery and Intestinal Colonization by Enterotoxigenic Escherichia coli. Clin. Vaccine Immunol. 2012, 19, 1603–1608. [Google Scholar] [CrossRef]
- Kumar, P.; Kuhlmann, F.M.; Chakraborty, S.; Bourgeois, A.L.; Foulke-Abel, J.; Tumala, B.; Vickers, T.J.; Sack, D.A.; DeNearing, B.; Harro, C.D.; et al. Enterotoxigenic Escherichia coli–blood group A interactions intensify diarrheal severity. J. Clin. Investig. 2018, 128, 3298–3311. [Google Scholar] [CrossRef]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L.; et al. Interrogation of a live-attenuated enterotoxigenic Escherichia coli vaccine highlights features unique to wild-type infection. NPJ Vaccines 2019, 4, 1–9. [Google Scholar] [CrossRef]
- Nesta, B.; Valeri, M.; Spagnuolo, A.; Rosini, R.; Mora, M.; Donato, P.; Alteri, C.J.; Del Vecchio, M.; Buccato, S.; Pezzicoli, A.; et al. SslE Elicits Functional Antibodies That Impair In Vitro Mucinase Activity and In Vivo Colonization by Both Intestinal and Extraintestinal Escherichia coli Strains. PLoS Pathog. 2014, 10, e1004124. [Google Scholar] [CrossRef]
- Tapader, R.; Bose, D.; Dutta, P.; Das, S.; Pal, A. SslE (YghJ), a Cell-Associated and Secreted Lipoprotein of Neonatal Septicemic Escherichia coli, Induces Toll-Like Receptor 2-Dependent Macrophage Activation and Proinflammation through NF-κB and MAP Kinase Signaling. Infect. Immun. 2018, 86, e00399–e00418. [Google Scholar] [CrossRef] [PubMed]
- Baldi, D.L.; Higginson, E.E.; Hocking, D.M.; Praszkier, J.; Cavaliere, R.; James, C.E.; Bennett-Wood, V.; Azzopardi, K.I.; Turnbull, L.; Lithgow, T.; et al. The Type II Secretion System and Its Ubiquitous Lipoprotein Substrate, SslE, Are Required for Biofilm Formation and Virulence of Enteropathogenic Escherichia coli. Infect. Immun. 2012, 80, 2042–2052. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Dotson, J.; Allen, K.P.; Fleckenstein, J.M. Identification and Molecular Characterization of EatA, an Autotransporter Protein of Enterotoxigenic Escherichia coli. Infect. Immun. 2004, 72, 1786–1794. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, F.M.; Laine, R.O.; Afrin, S.; Nakajima, R.; Akhtar, M.; Vickers, T.; Parker, K.; Nizam, N.N.; Grigura, V.; Goss, C.W.; et al. Contribution of Noncanonical Antigens to Virulence and Adaptive Immunity in Human Infection with Enterotoxigenic E. coli. Infect. Immun. 2021, 89. [Google Scholar] [CrossRef]
- Benjelloun-Touimi, Z.; Sansonetti, P.J.; Parsot, C. SepA, the major extracellular protein of Shigella flexneri: Autonomous secretion and involvement in tissue invasion. Mol. Microbiol. 1995, 17, 123–135. [Google Scholar] [CrossRef]
- Anderson, M.; Sansonetti, P.J.; Marteyn, B.S. Shigella Diversity and Changing Landscape: Insights for the Twenty-First Century. Front. Cell. Infect. Microbiol. 2016, 6, 45. [Google Scholar] [CrossRef]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L.; et al. Human Experimental Challenge with Enterotoxigenic Escherichia coli Elicits Immune Responses to Canonical and Novel Antigens Relevant to Vaccine Development. J. Infect. Dis. 2018, 218, 1436–1446. [Google Scholar] [CrossRef]
- Behrens, R.H.; Cramer, J.P.; Jelinek, T.; Shaw, H.; von Sonnenburg, F.; Wilbraham, D.; Weinke, T.; Bell, D.J.; Asturias, E.; Pauwells, H.L.; et al. Efficacy and safety of a patch vaccine containing heat-labile toxin from Escherichia coli against travellers’ diarrhoea: A phase 3, randomised, double-blind, placebo-controlled field trial in travellers from Europe to Mexico and Guatemala. Lancet Infect. Dis. 2014, 14, 197–204. [Google Scholar] [CrossRef]
- El-Kamary, S.S.; Cohen, M.B.; Bourgeois, A.L.; Van De Verg, L.; Bauers, N.; Reymann, M.; Pasetti, M.F.; Chen, W.H. Safety and Immunogenicity of a Single Oral Dose of Recombinant Double Mutant Heat-Labile Toxin Derived from Enterotoxigenic Escherichia coli. Clin. Vaccine Immunol. 2013, 20, 1764–1770. [Google Scholar] [CrossRef]
- Porter, C.K.; Riddle, M.S.; Tibble, D.R.; Bougeois, A.L.; McKenzie, R.; Isidean, S.D.; Sebeny, P.; Savarino, S.J. A systematic review of experimental infections with enterotoxigenic Escherichia coli (ETEC). Vaccine 2011, 29, 5869–5885. [Google Scholar] [CrossRef]
- McKenzie, R.; Bourgeois, A.L.; Frech, S.A.; Flyer, D.C.; Bloom, A.; Kazempour, K.; Glenn, G.M. Transcutaneous immunization with the heat-labile toxin (LT) of enterotoxigenic Escherichia coli (ETEC): Protective efficacy in a double-blind, placebo-controlled challenge study. Vaccine 2007, 25, 3684–3691. [Google Scholar] [CrossRef]
- Sheikh, A.; Tumula, B.; Vickers, T.J.; Alvarado, D.; Ciorba, M.A.; Bhuiyan, T.R.; Qadri, F.; Singer, B.B.; Fleckenstein, J.M. CEACAMs serve as toxin-stimulated receptors for enterotoxigenicEscherichia coli. Proc. Natl. Acad. Sci. USA 2020, 117, 29055–29062. [Google Scholar] [CrossRef]
- Read, L.T.; Hahn, R.W.; Thompson, C.C.; Bauer, D.L.; Norton, E.B.; Clements, J.D. Simultaneous Exposure to Escherichia coli Heat-Labile and Heat-Stable Enterotoxins Increases Fluid Secretion and Alters Cyclic Nucleotide and Cytokine Production. Infect. Immun. 2014, 82, 5308–5316. [Google Scholar] [CrossRef]
- Zegeye, E.; Govasli, M.; Sommerfelt, H.; Puntervoll, P. Development of an enterotoxigenic Escherichia coli vaccine based on the heat-stable toxin. Hum. Vaccines Immunother. 2018, 15, 1379–1388. [Google Scholar] [CrossRef]
- Duan, Q.; Huang, J.; Xiao, N.; Seo, H.; Zhang, W. Neutralizing Anti-Heat-Stable Toxin (STa) Antibodies Derived from Enterotoxigenic Escherichia coli Toxoid Fusions with STa Proteins Containing N12S, L9A/N12S, or N12S/A14T Mutations Show Little Cross-Reactivity with Guanylin or Uroguanylin. Appl. Environ. Microbiol. 2018, 84, e01737-17. [Google Scholar] [CrossRef]
- Govasli, M.L.; Diaz, Y.; Zegeye, E.D.; Darbakk, C.; Taxt, A.M.; Puntervoll, P. Purification and Characterization of Native and Vaccine Candidate Mutant Enterotoxigenic Escherichia coli Heat-Stable Toxins. Toxins 2018, 10, 274. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Davis, S.M.; Westra, C.; Vickers, T.J.; Fleckenstein, J.M. Molecular Determinants of Enterotoxigenic Escherichia coli Heat-Stable Toxin Secretion and Delivery. Infect. Immun. 2018, 86, e00526–e00618. [Google Scholar] [CrossRef]
- Poly, F.; Noll, A.J.; Riddle, M.S.; Porter, C.K. Update on Campylobacter vaccine development. Hum. Vaccines Immunother. 2019, 15, 1389–1400. [Google Scholar] [CrossRef]
- Maue, A.C.; Poly, F.; Guerry, P. A capsule conjugate vaccine approach to prevent diarrheal disease caused byCampylobacter jejuni. Hum. Vaccines Immunother. 2014, 10, 1499–1504. [Google Scholar] [CrossRef]
- Garlepy, C.L.; Eggleston, H.; Shoemaker, N.H.; Monteiro, M.A.; Beck, Z.; Matyas, G.R.; Poly, F.; Laird, R.M. Vaccination with a Campylobacter jejuni conjugate vaccine CJCV2 administered with liposomes containing monophosphoryl lipid A and QS-21 is protective in a zinc-deficient C. jejuni mouse model. In Proceedings of the 10th International Conference on Vaccine for Enteric Diseases, Lausanne, Switzerland, 16–18 October 2019. [Google Scholar]
- Lee, L.H., 3rd; Burg, E.; Baqar, S.; Bourgeois, A.L.; Burr, D.H.; Ewing, C.P.; Trust, T.J.; Guerry, P. Evaluation of a Truncated Recombinant Flagellin Subunit Vaccine against Campylobacter jejuni. Infect. Immun. 1999, 67, 5799–5805. [Google Scholar] [CrossRef]
- Blaser, M.J.; Duncan, D.J. Human serum antibody response to Campylobacter jejuni infection as measured in an enzyme-linked immunosorbent assay. Infect. Immun. 1984, 44, 292–298. [Google Scholar] [CrossRef]
- Blaser, M.J.; Hopkin, J.A.; Vasil, M.L. Campylobacter jejuni outer membrane proteins are antigenic for humans. Infect. Immun. 1984, 43, 986–993. [Google Scholar] [CrossRef]
- Martin, P.M.; Mathiot, J.; Ipero, J.; Kirimat, M.; Georges, A.J.; Georges-Courbot, M.C. Immune response to Campylobacter jejuni and Campylobacter coli in a cohort of children from birth to 2 years of age. Infect. Immun. 1989, 57, 2542–2546. [Google Scholar] [CrossRef]
- Nachamkin, I.; Hart, A.M. Western blot analysis of the human antibody response to Campylobacter jejuni cellular antigens during gastrointestinal infection. J. Clin. Microbiol. 1985, 21, 33–38. [Google Scholar] [CrossRef]
- Albert, M.J.; Haridas, S.; Steer, D.; Dhaunsi, G.S.; Smith, A.I.; Adler, B. Identification of a Campylobacter jejuni Protein That Cross-Reacts with Cholera Toxin. Infect. Immun. 2007, 75, 3070–3073. [Google Scholar] [CrossRef]
- Albert, M.J.; Mustafa, A.S.; Islam, A.; Haridas, S. Oral Immunization with Cholera Toxin Provides Protection against Campylobacter jejuni in an Adult Mouse Intestinal Colonization Model. mBio 2013, 4, e00246–e00313. [Google Scholar] [CrossRef]
- Nothaft, H.; Davis, B.; Lock, Y.Y.; Perez-Munoz, M.E.; Vinogradov, E.; Walter, J.; Coros, C.; Szymanski, C.M. Engineering the Campylobacter jejuni N-glycan to create an effective chicken vaccine. Sci. Rep. 2016, 6, 26511. [Google Scholar] [CrossRef]
- Walker, R.; Dull, P. Combination vaccine strategies to prevent enteric infections. Vaccine 2017, 35, 6790–6792. [Google Scholar] [CrossRef]
- Barry, E.; Cassels, F.; Riddle, M.; Walker, R.; Wierzba, T. Vaccines against Shigella and Enterotoxigenic Escherichia coli: A summary of the 2018 VASE Conference. Vaccine 2019, 37, 4768–4774. [Google Scholar] [CrossRef]
- Walker, R.I.; Clifford, A. Recommendations regarding the development of combined enterotoxigenic Eschericha coli and Shigella vaccines for infants. Vaccine 2015, 33, 946–953. [Google Scholar] [CrossRef]
- World Health Organization. DRAFT WHO Preferred Product Characteristics for Vaccines against Enterotoxigenic Escherichia coli. Published April 2020. Available online: https://www.who.int/immunization/research/ppc-tpp/PPC_ETEC_April_2020_Public_Consultation.pdf?ua=1 (accessed on 9 March 2021).
- Levine, M.M. Can needle-free administration of vaccines become the norm in global vaccination? Nat. Med. 2003, 9, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wai, T.T.; Bolick, D.T.; Li, M.; Gao, L.; Chakravarty, S.; James, E.R.; Zhang, W.; Sack, D.A.; Guerrant, R.L.; Hoffman, S.L.; et al. Protection of Mice against ETEC-induced Diarrhea and Weight Loss by Immunization with Bi-valent Recombinant Ty21a Typhoid; ASTMH: Arlington, VA, USA, 2020. [Google Scholar]
- Laird, R.M.; Ma, Z.; Dorabawila, N.; Pequegnat, B.; Omari, E.; Liu, Y.; Maue, A.C.; Poole, S.T.; Maciel, M.; Satish, K.; et al. Evaluation of a conjugate vaccine platform against enterotoxigenic Escherichia coli (ETEC), Campylobacter jejuni and Shigella. Vaccine 2018, 36, 6695–6702. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, R. Enhancement of Immune Response to ETEC Antigens Co-administered with Invaplex. Walter Reed Army Institute of Research: Silver Spring, MD, USA, Unpublished data.
- Kaminski, R.W.; Turbyfill, K.R.; Oaks, E.V. Mucosal Adjuvant Properties of the Shigella Invasin Complex. Infect. Immun. 2006, 74, 2856–2866. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kaminski, R.W.; Turbyfill, K.R.; Chao, C.; Ching, W.M.; Oaks, E.V. Mucosal Adjuvanticity of a Shigella Invasin Complex with DNA-Based Vaccines. Clin. Vaccine Immunol. 2009, 16, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, R.W.; Lee, L.F.; Turbyfill, K.R.; Scott, D.; Guerry, P.; Oaks, E.V. Shigella Invaplex enhances cellular and humoral immune responses to Campylobacter FlaA protein: Potential for enteric combination vaccine. In Proceedings of the Sixth Annual Conference on Vaccine Research, Arlington, VA, USA, 5–7 May 2003. [Google Scholar]
- Lal, M.; Jarrahian, C. Presentation matters: Buffers, packaging and delivery devices for new, oral enteric vaccines for infants. Hum. Vaccines Immunother. 2017, 13, 46–49. [Google Scholar] [CrossRef]
- White, J.A.; Haghighi, C.; Brunner, J.; Estrada, M.; Lal, M.; Chen, D. Preformulation studies with Escherichia coli double mutant heat-labile toxin adjuvant for use in an oral vaccine. J. Immunol. Methods 2017, 451, 83–89. [Google Scholar] [CrossRef]
- White, J.A.; Lal, M. Technical product attributes in development of an oral enteric vaccine. Vaccine 2019, 37, 4800–4804. [Google Scholar] [CrossRef]
- Chandrasekaran, L.; Lal, M.; Van De Verg, L.L.; Venkatesan, M. A study of different buffers to maximize viability of an oral Shigella vaccine. Vaccine 2015, 33, 6156–6160. [Google Scholar] [CrossRef][Green Version]
- Walker, R.I.; Clements, J.D. Use of heat-labile toxin of enterotoxigenic Escherichia coli to facilitate mucosal immunization. Vaccine Res. 1993, 2, 1–10. [Google Scholar]
- Dickinson, B.L.; Clements, J.D. Dissociation of Escherichia coli heat-labile enterotoxin adjuvanticity from ADP-ribosyltransferase activity. Infect. Immun. 1995, 63, 1617–1623. [Google Scholar] [CrossRef]
- Clements, J.D.; Norton, E.B. The Mucosal Vaccine Adjuvant LT(R192G/L211A) or dmLT. mSphere 2018, 3, e00215–e00218. [Google Scholar] [CrossRef]
- Lundgren, A.; Akhtar, M.; Kaim, J.; Cardeno, A.; Qadri, F.; Svennerholm, A.M. T cell responses induced by inactivated ETEC vaccine ETVAX given with and without dmLT adjuvant. In Proceedings of the 10th International Holmgren Conference on Vaccines for Enteric Diseases (VED 2019), Lausanne, Switzerland, 16–18 October 2019. [Google Scholar]
- Leach, S.; Clements, J.D.; Kaim, J.; Lundgren, A. The Adjuvant Double Mutant Escherichia coli Heat Labile Toxin Enhances IL-17A Production in Human T Cells Specific for Bacterial Vaccine Antigens. PLoS ONE 2012, 7, e51718. [Google Scholar] [CrossRef]
- Holmgren, J.; Parashar, U.D.; Plotkin, S.; Louis, J.; Ng, S.-P.; Desauziers, E.; Picot, V.; Saadatian-Elahi, M. Correlates of protection for enteric vaccines. Vaccine 2017, 35, 3355–3363. [Google Scholar] [CrossRef]
- Lee, T.; Gutiérrez, R.L.; Maciel, M., Jr.; Poole, S.; Test, K.J.; Trop, S.; Duplessis, C.; Lane, A.; Riddle, M.; Melinda Hamer, M.; et al. Safety and immunogenicity of intramuscularly administered CS6 subunit vaccine with a modified heat-labile enterotoxin from enterotoxigenic Escherichia coli. Unpublished; manuscript in preparation.
- Cholera vaccines: WHO position paper. Wkly. Epidmiol. Record 2017, 92, 477–500.
- Karlsson, S.L.; Ax, E.; Nygren, E.; Källgård, S.; Blomquist, M.; Ekman, A.; Benktander, J.; Holmgren, J.; Lebens, M. Develop-ment of stable Vibrio cholerae O1 Hikojima type vaccine strains co-expressing the Inaba and Ogawa lipopolysaccharide an-tigens. PLoS ONE 2014, 9, e108521. [Google Scholar] [CrossRef]
- Wierzba, T.F. Oral cholera vaccines and their impact on the global burden of disease. Hum. Vaccines Immunother. 2019, 15, 1294–1301. [Google Scholar] [CrossRef]
- Ranallo, R.T.; Kaminski, R.W.; George, T.; Kordis, A.A.; Chen, Q.; Szabo, K.; Venkatesan, M.M. Virulence, Inflammatory Potential, and Adaptive Immunity Induced by Shigella flexneri msbB Mutants. Infect. Immun. 2010, 78, 400–412. [Google Scholar] [CrossRef]
- Bergqvist, P.; Stensson, A.; Hazanov, L.; Holmberg, A.; Mattsson, J.; Mehr, R.; Bemark, M.; Lycke, N.Y. Re-utilization of the germinal centers in multiple Peyer’s patches results in highly synchronized, oligoclonal, and affinity-matured gut IgA responses. Mucosal Immunol. 2013, 6, 122–135. [Google Scholar] [CrossRef]
- Lundgren, A.; Jertborn, M.; Svennerholm, A.M. Induction of long term mucosal immunological memory in humans by an oral inactivated multivalent enterotoxigenic Escherichia coli vaccine. Vaccine 2016, 34, 3132–3140. [Google Scholar] [CrossRef]
- Giallourou, N.; Medlock, G.L.; Bolick, D.T.; Medeiros, P.H.; Ledwaba, S.E.; Kolling, G.L.; Tung, K.; Guerry, P.; Swann, J.R.; Guerrant, R.L. A novel mouse model of Campylobacter jejuni enteropathy and diarrhea. PLoS Pathog. 2018, 14, e1007083. [Google Scholar] [CrossRef]
- Bolick, D.T.; Medeiros, P.H.Q.S.; Ledwaba, S.E.; Lima, A.A.M.; Nataro, J.P.; Barry, E.M.; Guerrant, R.L. Critical Role of Zinc in a New Murine Model of Enterotoxigenic Escherichia coli Diarrhea. Infect. Immun. 2018, 86, e00183–e00218. [Google Scholar] [CrossRef]
- Medeiros, P.H.Q.S.; Ledwaba, S.E.; Bolick, D.T.; Giallourou, N.; Yum, L.K.; Costa, D.V.S.; Oriá, R.B.; Barry, E.M.; Swann, J.R.; Lima, A.Â.M.; et al. A murine model of diarrhea, growth impairment and metabolic disturbances with Shigella flexneri infection and the role of zinc deficiency. Gut Microbes 2019, 1–16. [Google Scholar] [CrossRef]
- Rollenhagen, J.E.; Jones, F.; Hall, E.; Maves, R.; Nunez, G.; Espinoza, N.; O’Dowd, A.; Prouty, M.G.; Savarino, S.J. Establishment, Validation, and Application of a New World Primate Model of Enterotoxigenic Escherichia coli Disease for Vaccine Development. Infect. Immun. 2019, 87, e00634–e00718. [Google Scholar] [CrossRef]
- Gregory, M.; Kaminski, R.W.; Lugo-Roman, L.A.; Galvez Carrillo, H.; Tilley, D.H.; Baldeviano, C.; Simons, M.P.; Reynolds, N.D.; Ranallo, R.T.; Suvarnapunya, A.E.; et al. Development of an Aotus nancymaae Model for Shigella Vaccine Immunogenicity and Efficacy Studies. Infect. Immun. 2014, 82, 2027–2036. [Google Scholar] [CrossRef]
- Islam, D.; Lewis, M.D.; Srijan, A.; Bodhidatta, L.; Aksomboon, A.; Gettayacamin, M.; Baqar, S.; Scott, D.; Mason, C.J. Establishment of a non-human primate Campylobacter disease model for the pre-clinical evaluation of Campylobacter vaccine formulations. Vaccine 2006, 24, 3762–3771. [Google Scholar] [CrossRef]
- Jin, C.; Gibani, M.M.; Moore, M.; Juel, H.B.; Jones, E.; Meiring, J.; Harris, V.; Gardner, J.; Nebykova, A.; Kerridge, S.A.; et al. Efficacy and immunogenicity of a Vi-tetanus toxoid conjugate vaccine in the prevention of typhoid fever using a controlled human infection model of Salmonella Typhi: A randomised controlled, phase 2b trial. Lancet 2017, 390, 2472–2480. [Google Scholar] [CrossRef]
- Meiring, J.E.; Giubilini, A.; Savulescu, J.; Pitzer, V.E.; Pollard, A.J. Generating the Evidence for Typhoid Vaccine Introduction: Considerations for Global Disease Burden Estimates and Vaccine Testing Through Human Challenge. Clin. Infect. Dis. 2019, 69, S402–S407. [Google Scholar] [CrossRef]
- Shakya, M.; Colin-Jones, R.; Theiss-Nyland, K.; Voyzey, M.; Pant, D.; Smith, N.; Liu, X.; Tonks, S.; Mazur, O.; Farooq, Y.G.; et al. Phase 3 Efficacy Analysis of a Typhoid Conjugate Vaccine Trial in Nepal. N. Engl. J. Med. 2019, 381, 2209–2218. [Google Scholar] [CrossRef]
- Chen, W.H.; Cohen, M.B.; Kirkpatrick, B.D.; Brady, R.C.; Galloway, D.; Gurwith, M.; Hall, R.H.; Kessler, R.A.; Lock, M.; Haney, D.; et al. Single-dose Live Oral Cholera Vaccine CVD 103-HgR Protects Against Human Experimental Infection with Vibrio cholerae O1 El Tor. Clin. Infect. Dis. 2016, 62, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, K.A.; Talaat, K.R.; Alaimo, C.; Martin, P.; Bourgeois, A.L.; Dreyer, A.; Porter, C.K.; Chakraborty, S.; Brubaker, J.; Elwood, D.; et al. Immune response characterization in a human challenge study with a Shigella flexneri 2a bioconjugate vaccine. EBioMedicine 2021, 66, 103308. [Google Scholar] [CrossRef] [PubMed]
- A Study to Evaluate the Efficacy, Safety, and Immunogenicity of a Vaccine Designed to Protect against Infection with Shigella sonnei in Healthy Adults. ClinicalTrials.gov Identifier: NCT03527173. Updated 28 July 2020. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03527173 (accessed on 12 April 2021).
- Kirkpatrick, B.D.; Lyon, C.E.; Porter, C.K.; Maue, A.C.; Guerry, P.; Pierce, K.K.; Carmolli, M.P.; Riddle, M.S.; Larsson, C.J.; Hawk, D.; et al. Lack of Homologous Protection Against Campylobacter jejuni CG8421 in a Human Challenge Model. Clin. Infect. Dis. 2013, 57, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.B.; Rylance, J.; Luck, A.; Jambo, K.; Ferreira, D.M.; Manda-Taylor, L.; Bejon, P.; Ngwira, B.; Littler, K.; Seager, Z.; et al. A framework for Controlled Human Infection Model (CHIM) studies in Malawi: Report of a Wellcome Trust workshop on CHIM in Low Income Countries held in Blantyre, Malawi. Wellcome Open Res. 2017, 2, 70. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, M.M.; Tribble, D.R.; Baqar, S.; Scott, D.A.; Ferris, J.A.; Walker, R.I.; Moran, A.P. In Vivo Phase Variation and Serologic Response to Lipooligosaccharide of Campylobacter jejuni in Experimental Human Infection. Infect. Immun. 2004, 72, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Rimmer, J.E.; Harro, C.; Sack, D.A.; Talaat, K.R.; Gutierrez, R.L.; DeNearing, B.; Brubaker, J.; Laird, R.M.; Poly, F.; Maue, A.C.; et al. Rifaximin Fails to Prevent Campylobacteriosis in the Human Challenge Model: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Infect. Dis. 2018, 66, 1435–1441. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Candidate Name [Developer] | Pre Clinical | Ph1 | Ph2 | Ph3 |
|---|---|---|---|---|
| Inactivated cellular candidates | ||||
| Truncated mutant (International Vaccine Institute (IVI), PATH) | X | |||
| Trivalent Shigella whole cell (PATH, Walter Reed Army Institute of Research (WRAIR)) | X | |||
| Inactivated Shigella (Hilleman Laboratories) | X | |||
| Live attenuated cellular candidates | ||||
| GuaBA-based live attenuated (CVD1208S; University of Maryland, Baltimore (UMB), PATH) | X | |||
| VirG-based live attenuated (WRSS1, WRSs2, WRSs3, and WRSf3; WRAIR, PATH) | X | |||
| Ty21a typhoid vaccine expressing Shigella LPS (Protein Potential) | X | |||
| ShigETEC (Eveliqure) | X | |||
| Glycoconjugate candidates | ||||
| Chemically prepared glycoconjugate (National Institute of Child Health and Human Development) | X | |||
| Recombinant glycoconjugate (LimmaTech) | X | |||
| Synthetic glycoconjugate (Institut Pasteur) | X | |||
| Subunit candidates | ||||
| InvaplexAR-DETOX (WRAIR, PATH) | X | |||
| Generalized Module for Membrane Antigens (GMMA; GSK Vaccine Institute for Global Health) | X | |||
| Outer Membrane Vesicles (OMV; University of Navarra) | X | |||
| Ipa DB Fusion (PATH, University of Kansas) | X | |||
| IpaB—GroEL fusion (Defense Inst. of Physiology and Allied Sciences) | X | |||
| 34kDa OMP (National Institute of Cholera and Enteric Diseases) | X | |||
| Candidate Name (Developer) | Pre Clinical | Ph1 | Ph2 | Ph3 |
|---|---|---|---|---|
| Inactivated cellular candidates | ||||
| ETVAX inactivated (Scandinavian Biopharma, University of Gothenburg, PATH) | X | |||
| STM (IVI, University of Georgia (UGA), PATH, Washington University (WASHU), WRAIR) | X | |||
| Live attenuated cellular candidates | ||||
| ACE527 live attenuated (PATH; National Vaccine and Serum Institute, UGA) | X | |||
| ShigETEC (Eveliqure) | X | |||
| CVD GuaBA Shigella−ETEC hybrid (UMB, Emergent) | X | |||
| Ty21a expressing Shigella LPS and MEFA (Protein Potential) | X | |||
| Subunit and toxin candidates | ||||
| FTA (PATH, Naval Medical Research Center (NMRC), Sanofi, IDRI) | X | |||
| MEFA (Kansas State University, John Hopkins University, PATH) | X | |||
| LT/ST Fusion/conjugate (ENTVAC Consortium, PATH) | X | |||
| dmLT mucosal adjuvant and antigen (PATH, Tulane) | X | |||
| Flagellin, EtpA, EatA, EaeH, and YghJ (WASHU) | X | |||
| Candidate Name [Developer] | Pre Clinical | Ph1 | Ph2 | Ph3 |
|---|---|---|---|---|
| Inactivated cellular candidates | ||||
| STM—heptasaccharide (IVI, PATH, UGA, and WRAIR) | X | |||
| ACE527 expressing heptasaccharide (PATH, UGA, University of Alberta (UA)) | X | |||
| Subunit candidates | ||||
| Capsular Polysaccharide (NMRC) | X | |||
| N-glycan Heptasaccharide (UGA, UA, PATH) | X | |||
| Truncated flagellin FlaA-MBP (NMRC) | X | |||
| B-subunit of cholera toxin (Kuwait University) | X | |||
| Route of Administration | Current Description | Status | Possible Added Pathogen Antigens |
|---|---|---|---|
| Oral Route | |||
| Live attenuated [developer] | |||
| CVD GuaBA Shigella-ETEC hybrid (UMB, Emergent) | Major O-PS Shigella serotypes expressing CFA/I and CS antigens as well as LTB and ST toxoid of ETEC | Prototype nearing Phase 1 | Campylobacter: conserved antigen such as Heptasaccharide or FlaA |
| ShigETEC (Eveliqure) | O-PS and T3SS-free S. flexneri 2a expressing ST and LT toxoids to cover ETEC along with Shigella antigens with homology to ETEC | Nearing Phase 1 | Campylobacter: conserved antigen such as Heptasaccharide FlaA |
| Ty21a expressing Shigella LPS and MEFA (Protein Potential) | Ty21a expressing Shigella LPS and ETEC colonization antigens as a MEFA | Preclinical | Campylobacter: conserved antigen such as Heptasaccharide or FlaA; Shigella: T3SS (Ipa) |
| Inactivated cells [developer] | |||
| Truncated Shigella mutant (IVI, PATH) | wzy mutant (O-PS side chain shortened to one repeating unit) of Sf2a + ETEC antigen homology + dmLT | Preclinical | ETEC: ETVAX; non-canonical ETEC antigens (i.e., EtpA, CssBA or YghJ) Campylobacter: Heptasaccharide or FlaA |
| Parenteral Route | |||
| Subunit (developer | |||
| Tri-pathogen conjugate (NMRC) | Shigella O-PS + ETEC adhesin proteins + Campylobacter CPS | Preclinical | Campylobacter: Heptasaccharide or FlaA to replace CPS; more conserved antigens could be included for ETEC and Shigella |
| Invaplex (WRAIR, PATH) | IpaB + IpaC + Shigella LPS with msbB mutation | Prototype Phase 1 completed | ETEC: CFA/I, CS3, CS6 and/or ETEC non canonical proteins Shigella: additional LPS serotypes Campylobacter: Heptasaccharide or FlaA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, R.; Kaminski, R.W.; Porter, C.; Choy, R.K.M.; White, J.A.; Fleckenstein, J.M.; Cassels, F.; Bourgeois, L. Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease. Microorganisms 2021, 9, 1382. https://doi.org/10.3390/microorganisms9071382
Walker R, Kaminski RW, Porter C, Choy RKM, White JA, Fleckenstein JM, Cassels F, Bourgeois L. Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease. Microorganisms. 2021; 9(7):1382. https://doi.org/10.3390/microorganisms9071382
Chicago/Turabian StyleWalker, Richard, Robert W. Kaminski, Chad Porter, Robert K. M. Choy, Jessica A. White, James M. Fleckenstein, Fred Cassels, and Louis Bourgeois. 2021. "Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease" Microorganisms 9, no. 7: 1382. https://doi.org/10.3390/microorganisms9071382
APA StyleWalker, R., Kaminski, R. W., Porter, C., Choy, R. K. M., White, J. A., Fleckenstein, J. M., Cassels, F., & Bourgeois, L. (2021). Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease. Microorganisms, 9(7), 1382. https://doi.org/10.3390/microorganisms9071382