
Molecular Detection and Genetic Identification of Rickettsia Infection in Ixodes granulatus Ticks, an Incriminated Vector for Geographical Transmission in Taiwan



Abstract
1. Introduction
2. Materials and Methods
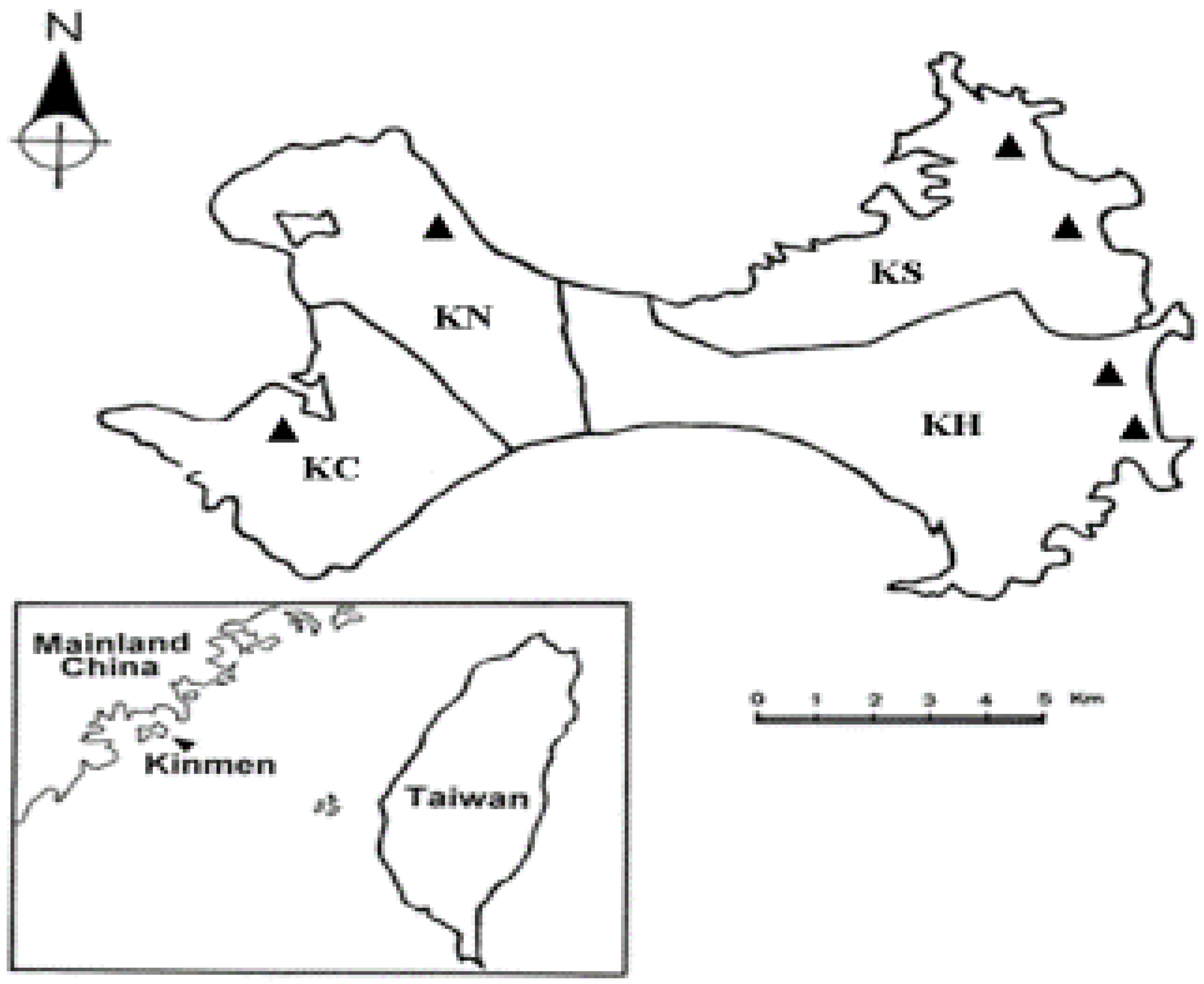
2.1. Tick Collection and Species Identification
2.2. DNA Extraction from Tick Specimens
2.3. DNA Amplification by Nested Polymerase Chain Reaction
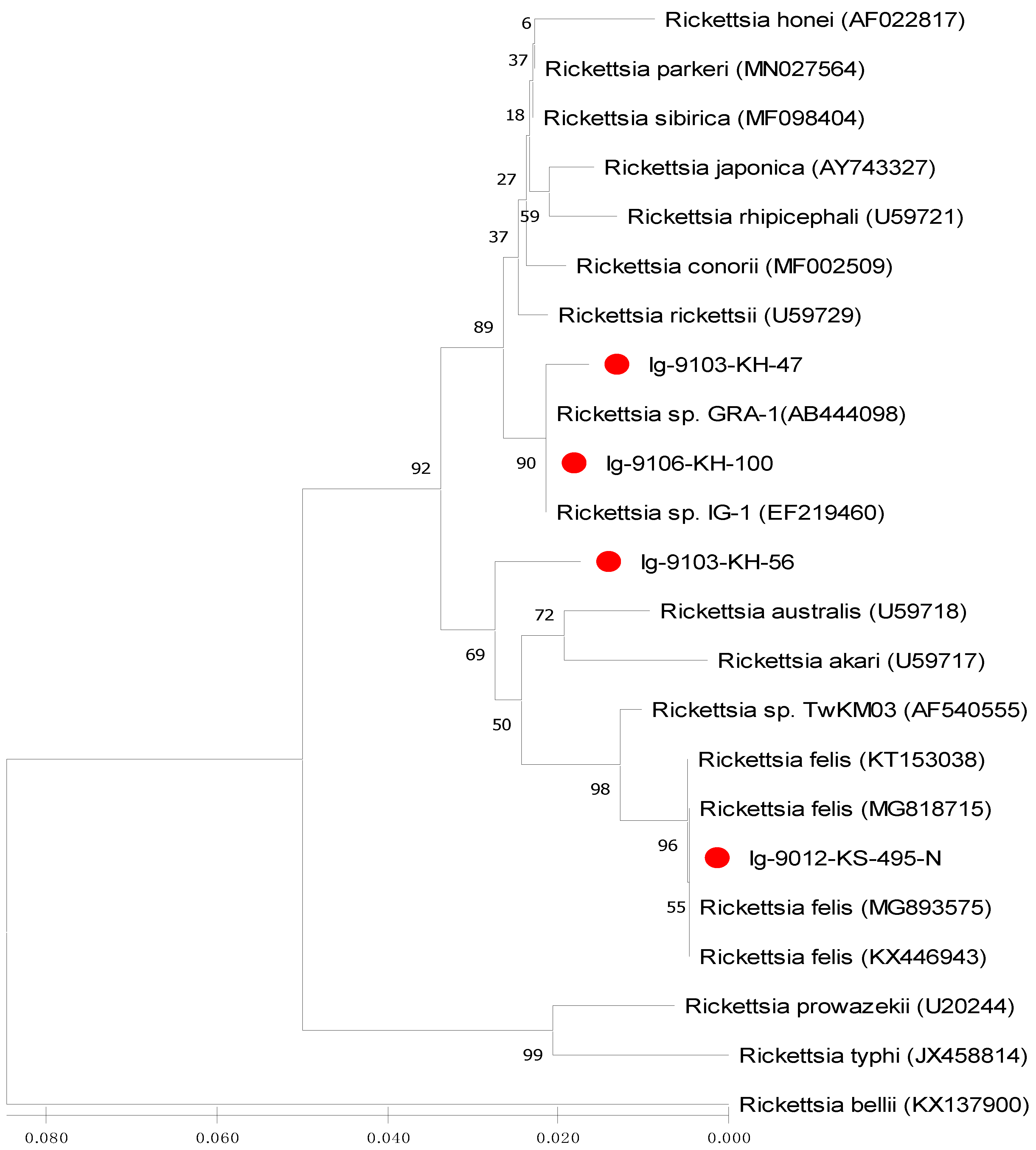
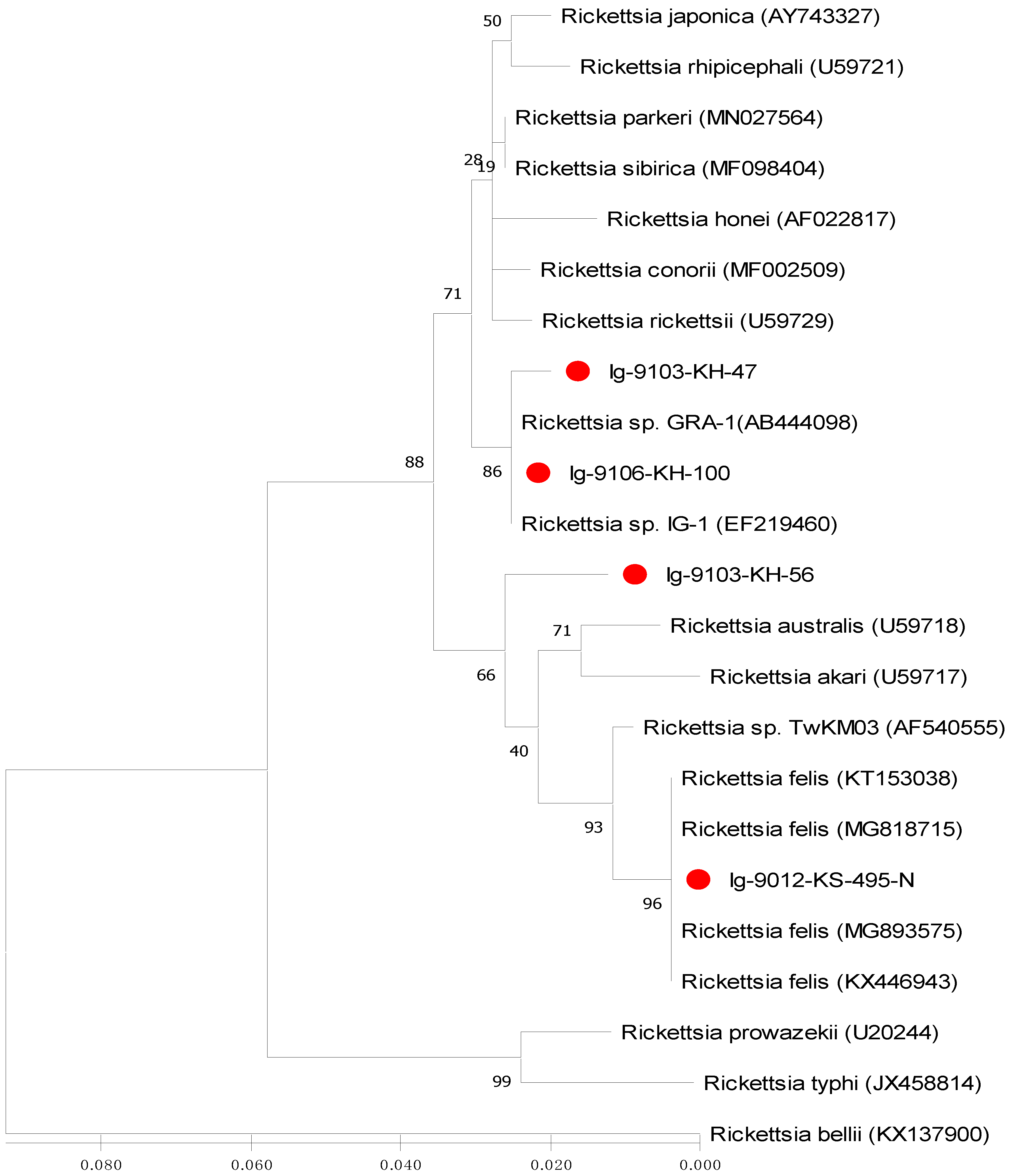
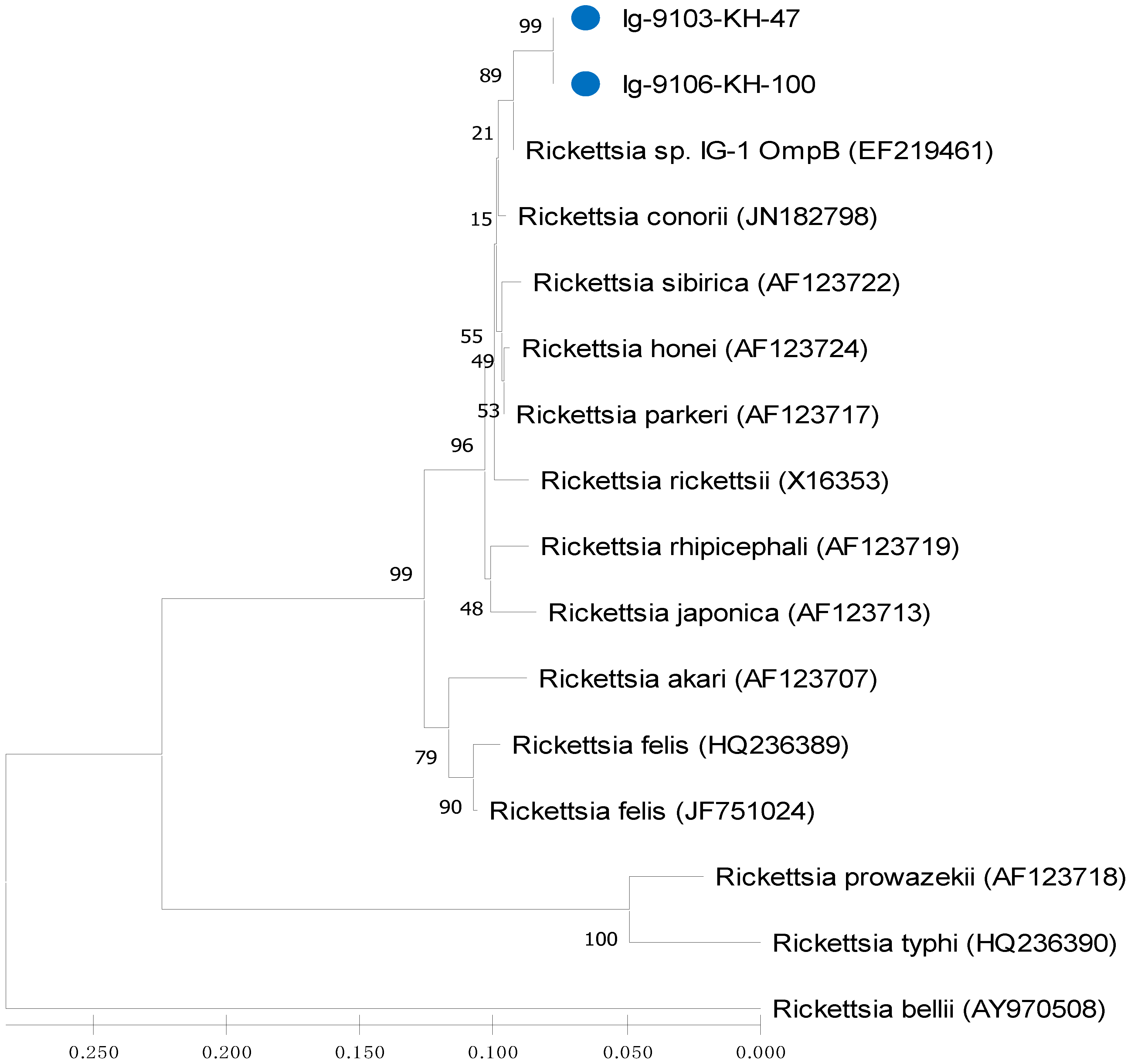
2.4. Sequence Alignments and Phylogenetic Analysis
2.5. Nucleotide Sequence Accession Numbers
3. Results
3.1. Detection of Rickettsia in I. granulatus Ticks from Taiwan
3.2. Sequence Alignment and Genetic Analysis of Rickettsia Detected in Ticks
3.3. Phylogenetic Analysis of Rickettsia Detected in Ticks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Fournier, P.E.; Dumler, J.S.; Greub, G.; Zhang, J.; Wu, Y.; Raoult, D. Gene sequence-based criteria for identification of new Rickettsia isolates and description of Rickettsia heilongjiangensis sp. nov. J. Clin. Microbiol. 2003, 41, 5456–5465. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Raoult, D. Tick-borne rickettsioses around the world: Emerging diseases challenging old concepts. Clin. Microbiol. Rev. 2005, 18, 719–756. [Google Scholar] [CrossRef] [PubMed]
- Gillesoie, J.J.; Williams, K.; Shukla, M.; Snyder, E.E.; Nordberg, E.K.; Ceraul, S.M.; Dharmanolla, C.; Rainey, D.; Soneja, J.; Shallom, J.M.; et al. Rickettsia phylogenimics: Unwinding the intricacies of obligate intracellular life. PLoS ONE 2008, 3, 2018. [Google Scholar]
- Azad, A.F.; Beard, C.B. Rickettsial pathogens and their arthropod vectors. Emerg. Infect. Dis. 1998, 4, 179–186. [Google Scholar] [CrossRef]
- Bremer, W.G.; Schaefer, J.J.; Wagner, E.R.; Ewing, S.A.; Rikihisa, Y.; Needham, G.R.; Jittapalapong, S.; Moore, D.L.; Stich, R.W. Transstadial and intrastadial experimental transmission of Ehrlichia canis by male Rhipicephalus sanguineus. Vet. Parasitol. 2005, 131, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez, S.E.; Troyo, A. A review of the genus Rickettsia in Central America. Res. Rep. Trop. Med. 2018, 9, 103–112. [Google Scholar] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Satjanadumrong, J.; Robinson, M.T.; Hughes, T.; Blacksel, S.D. Distribution and ecological drivers of spotted fever group Rickettsia in Asia. EcoHealth 2019, 16, 611–626. [Google Scholar] [CrossRef]
- Loginov, D.S.; Loginova, Y.F.; Dycka, F.; Böttinger, K.; Vechtova, P.; Sterba, J. Tissue-specific signatures in tick cell line MS profiles. Parasit. Vectors 2019, 12, 212. [Google Scholar] [CrossRef]
- Blazejak, K.; Janecek, E.; Strube, C. A 10-year surveillance of Rickettsiales (Rickettsia spp. and Anaplasma phagocytophilum) in the city of Hanover, Germany, reveals Rickettsia spp. as emerging pathogens in ticks. Parasit. Vectors 2017, 10, 588. [Google Scholar] [CrossRef]
- Choi, Y.J.; Jang, W.J.; Kim, J.H.; Ryu, J.S.; Lee, S.H.; Park, K.H.; Paik, H.S.; Koh, Y.S.; Choi, M.S.; Kim, I.S. Spotted fever group and typhus group rickettsioses in humans, South Korea. Emerg. Infect. Dis. 2005, 11, 237–244. [Google Scholar] [CrossRef]
- Teglas, M.; Matern, E.; Lein, S.; Foley, P.; Mahan, S.M.; Foley, J. Ticks and tick-borne disease in Guatemalan cattle and horses. Vet. Parasitol. 2005, 131, 119–127. [Google Scholar] [CrossRef]
- Unsworth, N.B.; Stenos, J.; Graves, S.R.; Faa, A.G.; Cox, G.E.; Dyer, J.R.; Boutlis, C.S.; Lane, A.M.; Shaw, M.D.; Robson, J.; et al. Flinders Island Spotted Fever rickettsiosis caused by marmionii strain of Rickettsia honei, Eastern Australia. Emerg. Infect. Dis. 2007, 13, 566–573. [Google Scholar] [CrossRef]
- Wood, H.; Drebot, M.A.; Dewailly, E.; Dillon, L.; Dimitrova, K.; Forde, M.; Grolla, A.; Lee, E.; Loftis, A.; Makowski, K.; et al. Short Report: Seroprevalence of seven zoonotic pathogens in pregnant women from the Caribbean. Am. J. Trop. Med. Hyg. 2014, 91, 642–644. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.G.; Junior, J.M.; Foster, R.J.; Harmsen, B.J.; Sanchez, E.; Martins, T.F.; Quigley, H.; Marcili, A.; Labruna, M.B. Ticks and rickettsiae from wildlife in Belize, Central America. Parasit. Vectors 2016, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Blanton, L.S.; Walker, D.H. Rickettsiae as emerging infectious agents. Clin. Lab. Med. 2017, 37, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jin, Y.; Fan, M.; Xu, G.; Liu, Q.; Raoult, D. Genotypic and antigenic identification of two new strains of spotted fever group rickettsiae isolated from China. J. Clin. Microbiol. 1993, 31, 83–88. [Google Scholar] [CrossRef]
- Uchida, T.; Uchiyama, T.; Kumano, K.; Walker, D.H. Rickettsia japonica sp. nov., the etiological agent of spotted fever group rickettsiosis in Japan. Int. J. Syst. Bacteriol. 1992, 42, 303–305. [Google Scholar] [CrossRef]
- Stenos, J.; Roux, V.; Walker, D.H.; Raoult, D. Rickettsia honei sp. nov., the aetiological agent of Flinders Island spotted fever in Australia. Int. J. Syst. Bacteriol. 1998, 48, 1309–1404. [Google Scholar] [CrossRef]
- Kollars, T.M., Jr.; Tippayachai, B.; Bodhidatta, D. Short report: Thai tick typhus, Rickettsia honei, and a unique rickettsia detected in Ixodes granulatus (Ixodidae: Acari) from Thailand. Am. J. Trop. Med. Hyg. 2001, 65, 535–537. [Google Scholar] [CrossRef]
- Fujita, H.; Fournier, P.E.; Takada, N.; Saito, T.; Raoult, D. Rickettsia asiatica sp. nov. isolated in Japan. Int. J. Syst. Evol. Microbiol. 2006, 56, 2365–2368. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fournier, P.E.; Takada, N.; Fujita, H.; Raoult, D. Rickettsia tamurae sp. nov. isolated from Amblyomma testudinarium ticks. Int. J. Syst. Evol. Microbiol. 2006, 56, 1673–1675. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Sun, Y.; Ju, W.; Wang, X.; Cao, W.; Wu, M. Vector competence of the tick Ixodes sinensis (Acari: Ixodidae) for Rickettsia monacensis. Parasit. Vectors 2014, 7, 512. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Montenegro, V.M.; Schicht, S.; Wolfel, S.; Schaper, S.R.; Chitimia-Dobler, L.; Siebert, S.; Strube, C. Detection of Rickettsia monacensis and Rickettsia amblyommatis in ticks collected from dogs in Costa Rica and Nicaragua. Ticks Tick Borne Dis. 2018, 9, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- da Paixao Seva, A.; Martins, T.F.; Munoz-Leal, S.; Rodrigues, A.C.; Pinter, A.; Luz, H.R.; Angerami, R.N.; Labruna, M.B. A human case of spotted fever caused by Rickettsia parkeri strain Atlantic rainforest and its association to the tick Amblyomma ovale. Parasit. Vectors 2019, 12, 471. [Google Scholar] [CrossRef] [PubMed]
- Balashov, Y.S. Bloodsucking ticks (Ixodoidea)-vectors of diseases of man and animals. Misc. Publ. Entomol. Soc. Am. 1972, 8, 268–305. [Google Scholar]
- Wilson, N. New distributional records of ticks from Southeast Asia and the Pacific (Metastigmata: Argasidae, Ixodidae). Orient. Insects 1970, 4, 37–46. [Google Scholar] [CrossRef]
- Chao, L.L.; Wu, W.J.; Shih, C.M. First detection and molecular identification of Borrelia burgdorferi- like spirochetes in Ixodes granulatus ticks collected on Kinmen Island of Taiwan. Am. J. Trop. Med. Hyg. 2009, 80, 389–394. [Google Scholar] [CrossRef]
- Chao, L.L.; Wu, W.J.; Shih, C.M. Molecular detection of Borrelia valaisiana-related spirochetes from Ixodes granulatus ticks in Taiwan. Exp. Appl. Acarol. 2010, 52, 393–407. [Google Scholar] [CrossRef]
- Fujita, H.; Kadosaka, T.; Nitta, Y.; Ando, S.; Takano, A.; Watanabe, H.; Kawabata, H. Rickettsia sp. in Ixodes granulatus ticks, Japan. Emerg. Infect. Dis. 2008, 14, 1963–1965. [Google Scholar] [CrossRef]
- Tsai, K.H.; Wang, H.C.; Chen, C.H.; Huang, J.H.; Lu, H.Y.; Su, C.L.; Shu, P.Y. Isolation and identification of a novel spotted fever group rickettsia, strain IG-1, from Ixodes granulatus ticks collected on Orchid Island (Lanyu), Taiwan. Am. J. Trop. Med. Hyg. 2008, 79, 256–261. [Google Scholar] [CrossRef]
- Robinson, M.T.; Satjannadumrong, J.; Hugbes, T.; Stenos, J.; Blacksell, S.D. Diagnosis of spotted fever group Rickettsia infections: The Asian perspective. Epidemiol. Infect. 2019, 147, e286. [Google Scholar] [CrossRef]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Bio. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 52, 1119–1134. [Google Scholar]
- Tsui, P.Y.; Tsai, K.H.; Weng, M.H.; Hung, Y.W.; Liu, Y.T.; Hu, K.Y.; Lien, J.C.; Lin, P.Y.; Shaio, M.F.; Wang, H.C.; et al. Molecular detection and characterization of spotted fever group Rickettsiae in Taiwan. Am. J. Trop. Med. Hyg. 2007, 77, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.B.; Bechelli, J.; Smalley, C.; Karim, S.; Walker, D.H. Vector tick transmission model of spotted fever rickettsiosis. Am. J. Pathol. 2019, 189, 115–123. [Google Scholar] [CrossRef]
- Blair, P.J.; Jiang, J.; Schoeler, G.B.; Moron, C.; Anaya, E.; Cespedes, M.; Cruz, C.; Felices, V.; Guevara, C.; Mendoza, L.; et al. Characterization of spotted fever group rickettsiae in flea and tick specimens from northern Peru. J. Clin. Microbiol. 2004, 42, 4961–4967. [Google Scholar] [CrossRef]
- Randolph, S.E.; Gern, L.; Nuttall, P.A. Co-feeding ticks epidemiological significance for tick-borne pathogen transmission. Parasitol. Today 1996, 12, 472–479. [Google Scholar] [CrossRef]
- EI-Sayed, A.; Kamel, M. Climate changes and their role in emergence and re-emergence of diseases. Environ. Sci. Pollut. Res. 2020, 27, 22336–22352. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Pena, A. Ticks as vectors: Taxonomy, biology and ecology. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 53–65. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genospecies/Strain | Origin of Rickettsia Strain | Gene Accession Number a | ||
|---|---|---|---|---|
| Biological | Geographic | gltA | ompB | |
| Taiwan strains | ||||
| Ig-9103-KH-56 | Ixodes granulatus | Kinmen, Taiwan | MT847612 | |
| Ig-9103-KH-47 | Ixodes granulatus | Kinmen, Taiwan | MT847613 | MZ198103 |
| Ig-9106-KH-100 | Ixodes granulatus | Kinmen, Taiwan | MT847614 | MZ198104 |
| Ig-9012-KS-495-N | Ixodes granulatus | Kinmen, Taiwan | MT847619 | |
| Rickettsia parkeri | Human skin | Brazil | MN027564 | |
| Rickettsia parkeri | Unknown | France | AF123717 | |
| Rickettsia sibirica | Hyalomma asiaticum | China | MF098404 | |
| Rickettsia sibirica | Unknown | France | AF123722 | |
| Rickettsia honei | Unknown | Australia | AF022817 | |
| Rickettsia honei | Unknown | France | AF123724 | |
| Rickettsia conorii | Rhipicephalus turanicus | China | MF002509 | |
| Rickettsia conorii | Human blood | Italy | JN182798 | |
| Rickettsia rickettsii | Dermacentor andersoni | Montana, USA | U59729 | |
| Rickettsia rickettsii | Dermacentor andersoni | Montana, USA | X16353 | |
| Rickettsia sp. GRA-1 | Ixodes granulatus | Japan | AB444098 | |
| Rickettsia sp. IG-1 | Ixodes granulatus | Lanyu, Taiwan | EF219460 | EF219461 |
| Rickettsia japonica | Unknown | Korea | AY743327 | |
| Rickettsia japonica | Unknown | France | AF123713 | |
| Rickettsia rhipicephali | Rhipicephalus sanguineus | USA | U59721 | |
| Rickettsia rhipicephali | Unknown | France | AF123719 | |
| Rickettsia akari | Human | New York, USA | U59717 | |
| Rickettsia akari | Unknown | France | AF123707 | |
| Rickettsia australis | Human | Australia | U59718 | |
| Rickettsia sp. TwKM03 | Ixodes granulatus | Taiwan | AF540555 | |
| Rickettsia felis | Cat flea | Malta | MG893575 | |
| Rickettsia felis | Lice | China | MG818715 | |
| Rickettsia felis | Cat flea | Brazil | KT153038 | |
| Rickettsia felis | Rat flea | Brazil | KX446943 | |
| Rickettsia felis | Flea | South Korea | HQ236389 | |
| Rickettsia felis | Rhipicephalus sanguineus | Chile | JF751024 | |
| Rickettsia prowazekii | Beetle | France | U20244 | |
| Rickettsia prowazekii | Unknown | France | AF123718 | |
| Rickettsia typhi | Human blood | Mexico | JX458814 | |
| Rickettsia typhi | Flea | Korea | HQ236390 | |
| Rickettsia bellii | Ixodes loricatus | Brazil | KX137900 | |
| Rickettsia bellii | Unknown | France | AY970508 | |
| Site of Collection a | Rickettsia Infection Detected in Life-Stage of Tick | Total | ||
|---|---|---|---|---|
| Nymph | Male | Female | No. Infected /No. Examined (%) | |
| No. Infected /No. Examined (%) | No. Infected /No. Examined (%) | No. Infected /No. Examined (%) | ||
| KH | 1/46 (2.17) | 0/14 (0) | 7/51 (13.73) | 8/111 (7.21) |
| KS | 3/36 (8.33) | 0/9 (0) | 1/57 (1.75) | 4/102 (3.92) |
| KN | 0/5 (0) | 0/0 (0) | 0/3 (0) | 0/8 (0) |
| KC | 0/18 (0) | 0/2 (0) | 0/6 (0) | 0/26 (0) |
| Total | 4/105 (3.81) | 0/25 (0) | 8/117 (6.84) | 12/247 (4.86) |
| Rickettsia Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Ig-9106-KH-100 (Taiwan) | — | ||||||||||||||||
| 2. Ig-9103-KH-47 (Taiwan) | 0.005 | — | |||||||||||||||
| 3. Rickettsia sp. IG-1 (EF219460) | 0.000 | 0.005 | — | ||||||||||||||
| 4. Rickettsia sp. GRA-1 (AB444098) | 0.000 | 0.005 | 0.000 | — | |||||||||||||
| 5. Rickettsia parkeri (MN027564) | 0.008 | 0.013 | 0.008 | 0.008 | — | ||||||||||||
| 6. Rickettsia sibirica (MF098404) | 0.008 | 0.013 | 0.008 | 0.008 | 0.000 | — | |||||||||||
| 7. Rickettsia conorii (MF002509) | 0.013 | 0.019 | 0.013 | 0.013 | 0.005 | 0.005 | — | ||||||||||
| 8. Rickettsia japonica (AY743327) | 0.016 | 0.021 | 0.016 | 0.016 | 0.008 | 0.008 | 0.013 | — | |||||||||
| 9. Rickettsia honei (AF022817) | 0.020 | 0.025 | 0.020 | 0.020 | 0.014 | 0.014 | 0.020 | 0.023 | — | ||||||||
| 10. Ig-9103-KH-56 (Taiwan) | 0.027 | 0.032 | 0.027 | 0.027 | 0.029 | 0.029 | 0.035 | 0.032 | 0.043 | — | |||||||
| 11. Rickettsia australis (U59718) | 0.041 | 0.046 | 0.041 | 0.041 | 0.032 | 0.032 | 0.038 | 0.035 | 0.049 | 0.035 | — | ||||||
| 12. Rickettsia TwKM03 (AF540555) | 0.038 | 0.043 | 0.038 | 0.038 | 0.035 | 0.035 | 0.041 | 0.038 | 0.049 | 0.027 | 0.030 | — | |||||
| 13. Ig-9012-KS-495-N (Taiwan) | 0.043 | 0.049 | 0.043 | 0.043 | 0.040 | 0.040 | 0.046 | 0.043 | 0.055 | 0.032 | 0.035 | 0.011 | — | ||||
| 14. Rickettsia felis (MG893575) | 0.043 | 0.049 | 0.043 | 0.043 | 0.040 | 0.040 | 0.046 | 0.043 | 0.055 | 0.032 | 0.035 | 0.012 | 0.000 | — | |||
| 15. Rickettsia felis (MG818715) | 0.043 | 0.049 | 0.043 | 0.043 | 0.040 | 0.040 | 0.046 | 0.043 | 0.055 | 0.032 | 0.035 | 0.011 | 0.000 | 0.000 | — | ||
| 16. Rickettsia typhi (JX458814) | 0.084 | 0.090 | 0.084 | 0.084 | 0.081 | 0.081 | 0.081 | 0.078 | 0.099 | 0.081 | 0.093 | 0.081 | 0.087 | 0.087 | 0.087 | — | |
| 17. Rickettsia bellii (KX137900) | 0.147 | 0.147 | 0.147 | 0.147 | 0.147 | 0.147 | 0.149 | 0.150 | 0.162 | 0.161 | 0.154 | 0.161 | 0.167 | 0.167 | 0.167 | 0.164 | 0.171 |
| Rickettsia Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Ig-9106-KH-47 (Taiwan) | — | |||||||||||||||
| 2. Ig-9103-KH-100 (Taiwan) | 0.000 | — | ||||||||||||||
| 3. Rickettsia sp. IG-1 (EF219461) | 0.016 | 0.016 | — | |||||||||||||
| 4. Rickettsia parkeri (AF123717) | 0.024 | 0.024 | 0.008 | — | ||||||||||||
| 5. Rickettsia conorii (JN182798) | 0.024 | 0.024 | 0.008 | 0.005 | — | |||||||||||
| 6. Rickettsia honei (AF123724) | 0.026 | 0.026 | 0.010 | 0.003 | 0.008 | — | ||||||||||
| 7. Rickettsia sibirica (AF123722) | 0.032 | 0.032 | 0.016 | 0.008 | 0.013 | 0.010 | — | |||||||||
| 8. Rickettsia rickettsia (X16353) | 0.034 | 0.034 | 0.018 | 0.016 | 0.016 | 0.018 | 0.024 | — | ||||||||
| 9. Rickettsia rhipicephali (AF123719) | 0.043 | 0.043 | 0.029 | 0.026 | 0.021 | 0.029 | 0.029 | 0.037 | — | |||||||
| 10. Rickettsia japonica (AF123713) | 0.046 | 0.046 | 0.030 | 0.027 | 0.027 | 0.030 | 0.030 | 0.038 | 0.032 | — | ||||||
| 11. Rickettsia felis (JF751024) | 0.068 | 0.068 | 0.051 | 0.048 | 0.049 | 0.051 | 0.057 | 0.059 | 0.062 | 0.066 | — | |||||
| 12. Rickettsia felis (HQ236389) | 0.070 | 0.070 | 0.064 | 0.061 | 0.059 | 0.064 | 0.070 | 0.073 | 0.067 | 0.080 | 0.011 | — | ||||
| 13. Rickettsia akari (AF123707) | 0.087 | 0.087 | 0.070 | 0.067 | 0.069 | 0.070 | 0.076 | 0.078 | 0.079 | 0.086 | 0.040 | 0.047 | — | |||
| 14. Rickettsia prowazekii (AF123718) | 0.356 | 0.356 | 0.341 | 0.333 | 0.343 | 0.333 | 0.332 | 0.342 | 0.333 | 0.351 | 0.313 | 0.321 | 0.321 | — | ||
| 15. Rickettsia typhi (HQ236390) | 0.375 | 0.375 | 0.366 | 0.357 | 0.365 | 0.357 | 0.356 | 0.349 | 0.357 | 0.377 | 0.332 | 0.330 | 0.362 | 0.077 | — | |
| 16. Rickettsia bellii (AY970508) | 0.484 | 0.484 | 0.478 | 0.468 | 0.468 | 0.474 | 0.472 | 0.470 | 0.475 | 0.468 | 0.471 | 0.477 | 0.487 | 0.546 | 0.563 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shih, C.-M.; Yang, P.-W.; Chao, L.-L. Molecular Detection and Genetic Identification of Rickettsia Infection in Ixodes granulatus Ticks, an Incriminated Vector for Geographical Transmission in Taiwan. Microorganisms 2021, 9, 1309. https://doi.org/10.3390/microorganisms9061309
Shih C-M, Yang P-W, Chao L-L. Molecular Detection and Genetic Identification of Rickettsia Infection in Ixodes granulatus Ticks, an Incriminated Vector for Geographical Transmission in Taiwan. Microorganisms. 2021; 9(6):1309. https://doi.org/10.3390/microorganisms9061309
Chicago/Turabian StyleShih, Chien-Ming, Pei-Wen Yang, and Li-Lian Chao. 2021. "Molecular Detection and Genetic Identification of Rickettsia Infection in Ixodes granulatus Ticks, an Incriminated Vector for Geographical Transmission in Taiwan" Microorganisms 9, no. 6: 1309. https://doi.org/10.3390/microorganisms9061309
APA StyleShih, C.-M., Yang, P.-W., & Chao, L.-L. (2021). Molecular Detection and Genetic Identification of Rickettsia Infection in Ixodes granulatus Ticks, an Incriminated Vector for Geographical Transmission in Taiwan. Microorganisms, 9(6), 1309. https://doi.org/10.3390/microorganisms9061309