
Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding

 ,
,  ,
,
Abstract
1. Introduction
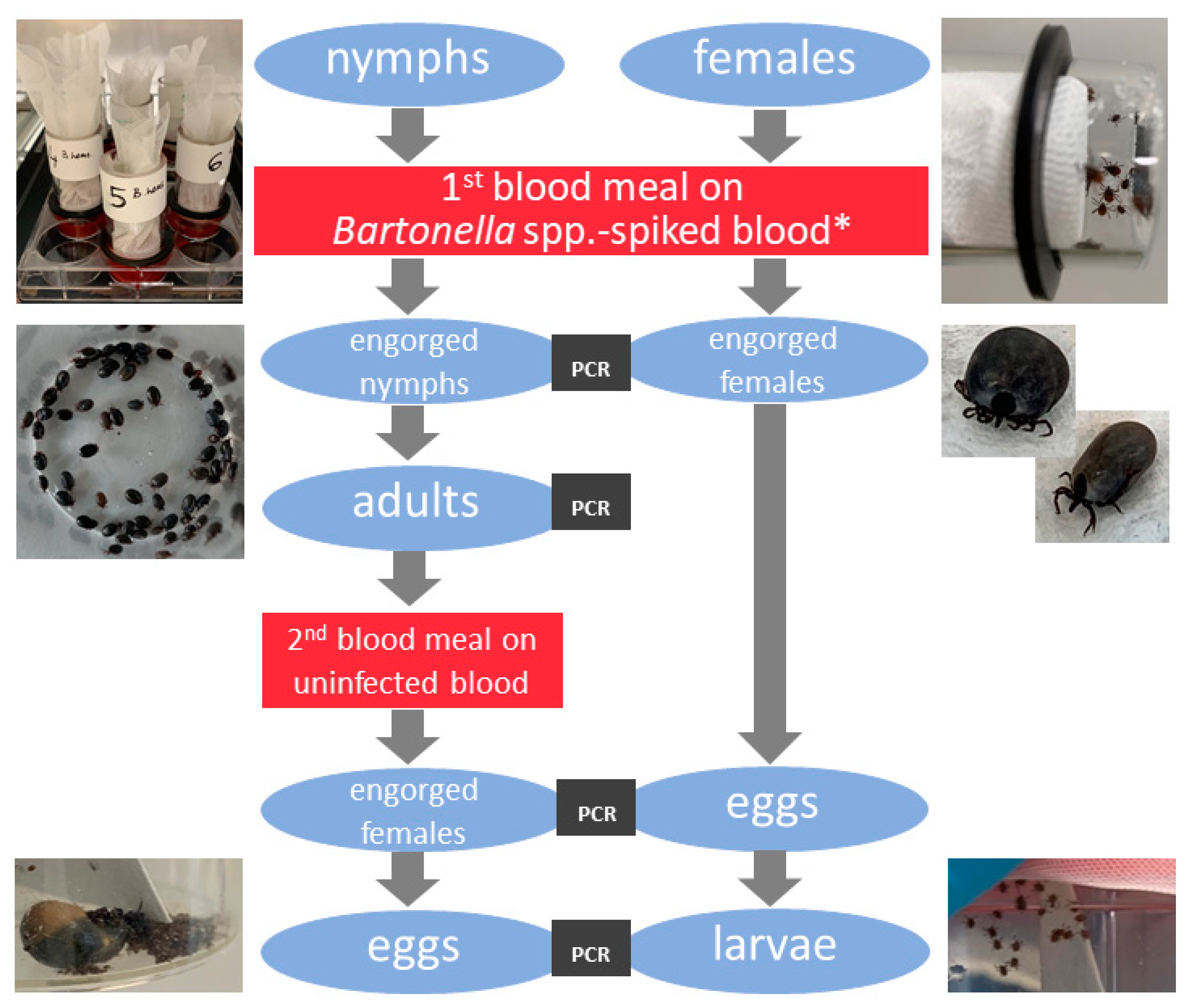
2. Materials and Methods
2.1. Cultivation of Bartonella colonies and Optical Density Measurement
2.2. Preparation of Blood for Experimental Tick Infection
2.3. Ticks
2.4. Preparation of Tick Feeding Units, Tick Feeding and Infection with Bartonella spp. via Artificial Feeding
2.5. Bartonella Detection in Ticks and Blood via PCR
2.6. Recultivation of Bartonella from Tick Gut and Salivary Glands
2.7. Statistical Analyses
3. Results
3.1. Tick Feeding and Ticks Development
3.2. Bartonella spp. Detection in Ticks
3.2.1. Bartonella spp. Acquisition by Ticks from a Blood Meal
3.2.2. Bartonella spp. Transstadial and Transovarial Transmission
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Prutsky, G.; Domecq, J.P.; Mori, L.; Bebko, S.; Matzumura, M.; Sabouni, A.; Shahrour, A.; Erwin, P.J.; Boyce, T.G.; Montori, V.M.; et al. Treatment outcomes of human bartonellosis: A systematic review and meta-analysis. Int. J. Infect. Dis. 2013, 17, e811–e819. [Google Scholar] [CrossRef] [PubMed]
- Rattanavong, S.; Fournier, P.-E.; Chu, V.; Frichitthavong, K.; Kesone, P.; Mayxay, M.; Mirabel, M.; Newton, P.N. Bartonella henselae Endocarditis in Laos—‘The Unsought Will Go Undetected’. PLoS Negl. Trop. Dis. 2014, 8, e3385. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Dehio, C. Role of distinct type-IV-secretion systems and secreted effector sets in host adaptation by pathogenic Bartonella species. Cell. Microbiol. 2019, 21, 1. [Google Scholar] [CrossRef]
- Maurin, M.; Raoult, D. Bartonella infections: Diagnostic and management issues. Curr. Opin. Infect. Dis. 1998, 11, 189–193. [Google Scholar] [CrossRef]
- Harms, A.; Dehio, C. Intruders below the Radar: Molecular Pathogenesis of Bartonella spp. Clin. Microbiol. Rev. 2012, 25, 42–78. [Google Scholar] [CrossRef]
- Gutiérrez, R.; Krasnov, B.; Morick, D.; Gottlieb, Y.; Khokhlova, I.S.; Harrus, S. Bartonella Infection in Rodents and Their Flea Ectoparasites: An Overview. Vector Borne Zoonotic Dis. 2015, 15, 27–39. [Google Scholar] [CrossRef]
- Birtles, R.J.; Hazel, S.M.; Bennett, M.; Bown, K.; Raoult, D.; Begon, M. Longitudinal monitoring of the dynamics of infections due to Bartonella species in UK woodland rodents. Epidemiol. Infect. 2001, 126, 323–329. [Google Scholar] [CrossRef]
- Bajer, A.; Pawelczyk, A.; Behnke, J.M.; Gilbert, F.S.; Sinski, E. Factors affecting the component community structure of haemoparasites in bank voles (Clethrionomys glareolus) from the Mazury Lake District region of Poland. Parasitology 2001, 122, 43–54. [Google Scholar] [CrossRef]
- Cotté, V.; Bonnet, S.; Le Rhun, D.; Le Naour, E.; Chauvin, A.; Boulouis, H.-J.; Lecuelle, B.; Lilin, T.; Vayssier-Taussat, M. Transmission of Bartonella henselae by Ixodes ricinus. Emerg. Infect. Dis. 2008, 14, 1074–1080. [Google Scholar] [CrossRef]
- Ebani, V.V.; Bertelloni, F.; Turchi, B.; Filogari, D.; Cerri, D. Molecular survey of tick-borne pathogens in Ixodid ticks collected from hunted wild animals in Tuscany, Italy. Asian Pac. J. Trop. Med. 2015, 8, 714–717. [Google Scholar] [CrossRef]
- Vayssier-Taussat, M.; Moutailler, S.; Féménia, F.; Raymond, P.; Croce, O.; La Scola, B.; Fournier, P.-E.; Raoult, D. Identification of Novel Zoonotic Activity of Bartonella spp., France. Emerg. Infect. Dis. 2016, 22, 457–462. [Google Scholar] [CrossRef]
- Regier, Y.; Komma, K.; Weigel, M.; Kraiczy, P.; Laisi, A.; Pulliainen, A.T.; Hain, T.; Kempf, V.A.J. Combination of microbiome analysis and serodiagnostics to assess the risk of pathogen transmission by ticks to humans and animals in central Germany. Parasit. Vectors 2019, 12, 699. [Google Scholar] [CrossRef]
- Silaghi, C.; Pfeffer, M.; Kiefer, D.; Kiefer, M.; Obiegala, A. Bartonella, Rodents, Fleas and Ticks: A Molecular Field Study on Host-Vector-Pathogen Associations in Saxony, Eastern Germany. Microb. Ecol. 2016, 72, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; La Fuente, J.; de Cote, M.; Galindo, R.C.; Moutailler, S.; Vayssier-Taussat, M.; Bonnet, S.I. IrSPI, a Tick Serine Protease Inhibitor Involved in Tick Feeding and Bartonella henselae Infection. PLoS Negl. Trop. Dis. 2014, 8, e2993. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D.; Lynn, G.E.; Burkhardt, N.Y.; Price, L.D.; Nelson, C.M.; Kurtti, T.J.; Munderloh, U.G. Infection of Immature Ixodes scapularis (Acari: Ixodidae) by Membrane Feeding. J. Med. Entomol. 2016, 53, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.J.; Xu, G.; Rich, S.M. A silicone membrane for in vitro feeding of Ixodes scapularis (Ixodida: Ixodidae). J. Med. Entomol. 2014, 51, 878–879. [Google Scholar] [CrossRef] [PubMed]
- Fourie, J.J.; Evans, A.; Labuschagne, M.; Crafford, D.; Madder, M.; Pollmeier, M.; Schunack, B. Transmission of Anaplasma phagocytophilum (Foggie, 1949) by Ixodes ricinus (Linnaeus, 1758) ticks feeding on dogs and artificial membranes. Parasit. Vectors 2019, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Koci, J.; Bernard, Q.; Yang, X.; Pal, U. Borrelia burgdorferi surface protein Lmp1 facilitates pathogen dissemination through ticks as studied by an artificial membrane feeding system. Sci. Rep. 2018, 8, 166. [Google Scholar] [CrossRef]
- Böhme, B.; Krull, C.; Clausen, P.-H.; Nijhof, A.M. Evaluation of a semi-automated in vitro feeding system for Dermacentor reticulatus and Ixodes ricinus adults. Parasitol. Res. 2018, 117, 565–570. [Google Scholar] [CrossRef]
- Krull, C.; Böhme, B.; Clausen, P.-H.; Nijhof, A.M. Optimization of an artificial tick feeding assay for Dermacentor reticulatus. Parasit. Vectors. 2017, 10, 453. [Google Scholar] [CrossRef]
- Krull, C. Optimization and Automation of Artificial Tick Feeding; Freie Universität Berlin: Berlin, Germany, 2020. [Google Scholar]
- Kröber, T.; Guerin, P.M. In Vitro feeding assays for hard ticks. Trends Parasitol. 2007, 23, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Kröber, T.; Guerin, P.M. An In Vitro feeding assay to test acaricides for control of hard ticks. Pest. Manag. Sci. 2007, 63, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.H.; Bai, Y.; Malania, L.; Winchell, J.M.; Kosoy, M.Y. Development of a Novel Genus-Specific Real-Time PCR Assay for Detection and Differentiation of Bartonella Species and Genotypes. J. Clin. Microbiol. 2012, 50, 1645–1649. [Google Scholar] [CrossRef]
- Maggi, R.G.; Diniz, P.P.; Cadenas, M.B.; Breitschwerdt, E.B. The use of molecular diagnostic techniques to detect Anaplasma, Bartonella and Ehrlichia species in arthropods or patients. In The International Canine Vector-Borne Disease Symposium; Billesley: Alcester, UK, 2006. [Google Scholar]
- Wechtaisong, W.; Bonnet, S.I.; Lien, Y.-Y.; Chuang, S.-T.; Tsai, Y.-L. Transmission of Bartonella henselae within Rhipicephalus sanguineus: Data on the Potential Vector Role of the Tick. PLoS Negl. Trop. Dis. 2020, 14, e0008664. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, E.; Wijnveld, M.; Bonga, M.; Berger, L.; Manfredi, M.T.; Veronesi, F.; Jongejan, F. Transmission of Rickettsia raoultii and Rickettsia massiliae DNA by Dermacentor reticulatus and Rhipicephalus sanguineus (s.l.) ticks during artificial feeding. Parasit. Vectors 2018, 11, 694. [Google Scholar] [CrossRef]
- Bonnet, S.; Brisseau, N.; Hermouet, A.; Jouglin, M.; Chauvin, A. Experimental in vitro transmission of Babesia sp. (EU1) by Ixodes ricinus. Vet. Res. 2009, 40, 21. [Google Scholar] [CrossRef]
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef]
- Liu, X.Y.; Cote, M.; Paul, R.E.L.; Bonnet, S.I. Impact of feeding system and infection status of the blood meal on Ixodes ricinus feeding. Ticks Tick Borne Dis. 2014, 5, 323–328. [Google Scholar] [CrossRef]
- Militzer, N.; Bartel, A.; Clausen, P.H.; Hoffmann-Köhler, P.; Nijhof, A.M. Artificial Feeding of All Consecutive Life Stages of Ixodes ricinus. Vaccines 2021, 9, 385. [Google Scholar] [CrossRef]
- Knorr, S.; Anguita, J.; Cortazar, J.T.; Hajdusek, O.; Kopáček, P.; Trentelman, J.J.; Kershaw, O.; Hovius, J.W.; Nijhof, A.M. Preliminary Evaluation of Tick Protein Extracts and Recombinant Ferritin 2 as Anti-tick Vaccines Targeting Ixodes ricinus in Cattle. Front. Physiol. 2018, 9, 1696. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.; Morway, C.; Sheff, K.W.; Bai, Y.; Colborn, J.; Chalcraft, L.; Dowell, S.F.; Peruski, L.F.; Maloney, S.A.; Baggett, H.; et al. Bartonella tamiae sp. nov., a Newly Recognized Pathogen Isolated from Three Human Patients from Thailand. J. Clin. Microbiol. 2008, 46, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Halos, L.; Jamal, T.; Maillard, R.; Beugnet, F.; Le Menach, A.; Boulouis, H.-J.; Vayssier-Taussat, M. Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet. Res. 2005, 36, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Skotarczak, B.; Adamska, M. Detection of Bartonella DNA in roe deer (Capreolus capreolus) and in ticks removed from deer. Eur. J. Wildl. Res. 2005, 51, 287–290. [Google Scholar] [CrossRef]
- Tsai, Y.-L.; Chomel, B.B.; Chang, C.-C.; Kass, P.H.; Conrad, P.A.; Chuang, S.-T. Bartonella and Babesia infections in cattle and their ticks in Taiwan. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Billeter, S.A.; Kasten, R.W.; Killmaster, L.F.; Breitschwerdt, E.B.; Levin, M.L.; Levy, M.G.; Kosoy, M.Y.; Chomel, B.B. Experimental infection by capillary tube feeding of Rhipicephalus sanguineus with Bartonella vinsonii subspecies berkhoffii. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.; Cote, M.; Le Rhun, D.; Lecuelle, B.; Levin, M.L.; Vayssier-Taussat, M.; Bonnet, S.I. Vector Competence of the Tick Ixodes ricinus for Transmission of Bartonella birtlesii. PLoS Negl. Trop. Dis. 2011, 5, e1186. [Google Scholar] [CrossRef]
- Florin, T.A.; Zaoutis, T.E.; Zaoutis, L.B. Beyond Cat Scratch Disease: Widening Spectrum of Bartonella henselae Infection. Pediatrics 2008, 121, e1413–e1425. [Google Scholar] [CrossRef]
- Wikswo, M.E.; Hu, R.; Metzger, M.E.; Eremeeva, M.E. Detection of Rickettsia rickettsii and Bartonella henselae in Rhipicephalus sanguineus Ticks from California. J. Med. Entomol. 2007, 44, 158–162. [Google Scholar] [CrossRef]
- Podsiadly, E.; Chmielewski, T.; Sochon, E.; Tylewska-Wierzbanowska, S. Bartonella henselae in Ixodes ricinus Ticks Removed from Dogs. Vector Borne Zoonotic Dis. 2007, 7, 189–192. [Google Scholar] [CrossRef]
- Sanogo, Y.O.; Zeaiter, Z.; Caruso, G.; Merola, F.; Shpynov, S.; Brouqui, P.; Raoult, D. Bartonella henselae in Ixodes ricinus Ticks (Acari: Ixodida) Removed from Humans, Belluno Province, Italy. Emerg. Infect. Dis. 2003, 9, 329–332. [Google Scholar] [CrossRef]
- Angelakis, E.; Pulcini, C.; Waton, J.; Imbert, P.; Socolovschi, C.; Edouard, S.; Dellamonica, P.; Raoult, D. Scalp Eschar and Neck Lymphadenopathy Caused by Bartonella henselae after Tick Bite. Clin. Infect. Dis. 2010, 50, 549–551. [Google Scholar] [CrossRef]
- Daveu, R.; Laurence, C.; Bouju, A.; Sassera, D.; Plantard, O. Symbiont dynamics during the blood meal of Ixodes ricinus nymphs according to their sex. Ticks. Tick. Born. Dis. 2021, 101707. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, A.; van Leeuwen, A.D.; Jahfari, S.; Takken, W.; Földvári, M.; Dremmel, L.; Sprong, H.; Földvári, G. Vertical transmission of Bartonella schoenbuchensis in Lipoptena cervi. Parasit. Vectors 2015, 8, 1489. [Google Scholar] [CrossRef]
- Chomel, B.B.; Boulouis, H.-J.; Maruyama, S.; Breitschwerdt, E.B. Bartonella Spp. in Pets and Effect on Human Health. Emerg. Infect. Dis. 2006, 12, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Oksi, J.; Rantala, S.; Kilpinen, S.; Silvennoinen, R.; Vornanen, M.; Veikkolainen, V.; Eerola, E.; Pulliainen, A.T. Cat Scratch Disease Caused by Bartonella grahamii in an Immunocompromised Patient. J. Clin. Microbiol. 2013, 51, 2781–2784. [Google Scholar] [CrossRef] [PubMed]
- Janecek, E.; Mietze, A.; Goethe, R.; Schnieder, T.; Strube, C. Bartonella spp. Infection Rate and B. grahamii in Ticks. Emerg. Infect. Dis. 2012, 18, 1689–1690. [Google Scholar] [CrossRef]
- Rolain, J.M.; Rousset, E.; La Scola, B.; Duquesnel, R.; Raoult, D. Bartonella schoenbuchensis Isolated from the Blood of a French Cow. Ann. N. Y. Acad. Sci. 2003, 990, 236–238. [Google Scholar] [CrossRef]
- Lei, B.R.; Olival, K.J. Contrasting Patterns in Mammal–Bacteria Coevolution: Bartonella and Leptospira in Bats and Rodents. PLoS Negl. Trop. Dis. 2014, 8, e2738. [Google Scholar] [CrossRef]


{kind=link}
| Life Stage | Blood Meal Spiked with | No. of Feeding Ticks | Engorgement Proportion (%, (no. of Engorged)) | Mean Weight after Feeding (mg, (SD) *) |
|---|---|---|---|---|
| Nymphs | B. grahamii | 300 | 54.6 (n = 164) | 2.71 |
| B. henselae | 291 | 65.3 (n = 190) | 3.01 | |
| B. schoenbuchensis | 291 | 58.1 (n = 169) | 3.21 | |
| PBS | 139 | 46.0 (n = 64) a | 2.82 | |
| total | 1021 | 57.5 (n = 587) | 2.94 | |
| Females | B. grahamii | 52 (+52 males) | 57.7 (n = 30) b | 154.7 (±81.99) |
| B. henselae | 42 (+42 males) | 64.3 (n = 27) | 175.6 (±85.83) | |
| B. schoenbuchensis | 42 (+42 males) | 85.7 (n = 36) | 168.1 (±75.80) | |
| PBS | 20 (+20 males) | 90 (n = 18) | 189.6 (±94.89)) | |
| total | 156 (+156 males) | 71.2 (n = 111) | 170.7 (±82.63) |
| Developmental Stages | Developmental Success for Ticks Feeding on Blood Spiked with | |||
|---|---|---|---|---|
| B. grahamii | B. henselae | B. schoenbuchensis | PBS | |
| Engorged nymphs molting into adults (%; no. molted adults/no. engorged nymphs) | 38.4 (63 (29m:34f)/164) | 38.4 (73 (44m:29f)/190) | 34.9 (59 (24m:35f)/169) | 11.4 a (5 (2m:3f)/44) |
| Engorged females laying eggs (%; no. females laying eggs/no. engorged females) | 50 (15/30) b | 74 (20/27) | 91.7 (33/36) | 66.7 (12/18) |
| Eggs molting into larvae (%; no. females producing larvae/no. females laying eggs) | 33.3 (5/15) | 40 (8/20) | 48.5 (16/33) | 58.3 (7/12) |
| Tick Life Stage | Bartonella spp. Detection (%; (no. Positive/no. Tested)) in Ticks Fed on Blood Spiked with | Total | ||
|---|---|---|---|---|
| B. grahamii | B. henselae | B. schoenbuchensis | ||
| Nymphs | 17.6 (3/17) | 40 (8/20) | 47.6 (10/21) | 36.2 (21/58) |
| Females | 30.8 (4/13) | 20 (4/20) | 41.1 (12/29) | 32.3 (20/62) |
| Total | 23.3 (7/30) | 30 (12/40) | 44 (22/50) | 34.2 (41/120) |
| Developmental Stage | Bartonella spp. Detection (%; (no. Positive/no. Tested)) in Ticks Infected with | ||
|---|---|---|---|
| B. grahamii | B. henselae | B. schoenbuchensis | |
| Adults molted from potentially infected nymphs | 7.7 (3/39) | 1.3 (1/44) | 18.2 (6/33) a |
| Potentially infected females that produced infected eggs and larvae | 18.2 (2/11) | 18.6 (3/16) | 26.1 (6/23) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Król, N.; Militzer, N.; Stöbe, E.; Nijhof, A.M.; Pfeffer, M.; Kempf, V.A.J.; Obiegala, A. Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding. Microorganisms 2021, 9, 901. https://doi.org/10.3390/microorganisms9050901
Król N, Militzer N, Stöbe E, Nijhof AM, Pfeffer M, Kempf VAJ, Obiegala A. Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding. Microorganisms. 2021; 9(5):901. https://doi.org/10.3390/microorganisms9050901
Chicago/Turabian StyleKról, Nina, Nina Militzer, Elisa Stöbe, Ard M. Nijhof, Martin Pfeffer, Volkhard A. J. Kempf, and Anna Obiegala. 2021. "Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding" Microorganisms 9, no. 5: 901. https://doi.org/10.3390/microorganisms9050901
APA StyleKról, N., Militzer, N., Stöbe, E., Nijhof, A. M., Pfeffer, M., Kempf, V. A. J., & Obiegala, A. (2021). Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding. Microorganisms, 9(5), 901. https://doi.org/10.3390/microorganisms9050901