
Regulation of tert-Butyl Hydroperoxide Resistance by Chromosomal OhrR in A. baumannii ATCC 19606



Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Culturing Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Description | Antibiotic Resistance (µg/mL) | Reference/Source |
|---|---|---|---|
| pK18mobsecB | Suicide vector for homologous recombination | Km50 | [23] |
| pK18dohrR | pK18mobsecB contains the upstream and downstream regions of ohrR-C | Km50 | This study |
| pK18dohr | pK18mobsecB contains the upstream and downstream regions of ohr-C | Km50 | This study |
| pK18dohrRohr | pK18mobsecB contains the upstream and downstream regions of ohrR-C-ohr-C | Km50 | This study |
| pQE80L | Expression vector with colE1 origin for His-tag fusion protein purification | Amp50 | Qiagen |
| pOhrR-C | ohrR-C gene with N-terminal-fused His-tag in pQE80L | Amp50 | This study |
| pOhrR-C-Pohr-ohr-C | ohr-C and ohrR-C gene promoter was cloned downstream of pOhrR-C | Amp50 | This study |
| Strain | Description | Reference/Source | |
| E. coli DH5α | F−, supE44, hsdR17, recA1, gyrA96, endA1, thi-1, relA1, deoR, λ− | ATCC 53868 | |
| E. coli S17-1λpir | thi-, thr , leu , tonA , lacY , supE , recA::RP4-2-Tc::Mu, Smr, lpir | [23] | |
| Acinetobacter baumannii ATCC 19606 | Primary strain used in this study | [19] | |
| Acinetobacter baumannii ATCC 17978 | Most studied strain to date | [14] | |
| ∆ohrR | Marker-less ohrR-c deletion mutant | This study | |
| ∆ohr | Marker-less ohr-c deletion mutant | This study | |
| ∆ohrRohr | Marker-less ohrR-c-ohr-c deletion mutant | This study | |
2.2. Markerless Mutant Generation
2.3. Minimum Inhibition Concentration
2.4. Disk Diffusion Assay
2.5. RNA Extraction and qRT-PCR
2.6. OhrR Overexpression and Purification
2.7. EMSA
2.8. G. Mellonella Experiments
3. Results
3.1. Identification of Two Paralogous ohr-ohrR Genes in A. baumannii ATCC 19606
3.2. Role of ohr-p and ohrR-p in tBHP Resistance Is Limited
3.3. Chromosomal ohr-C Plays an Important Role in Bacterial Proliferation
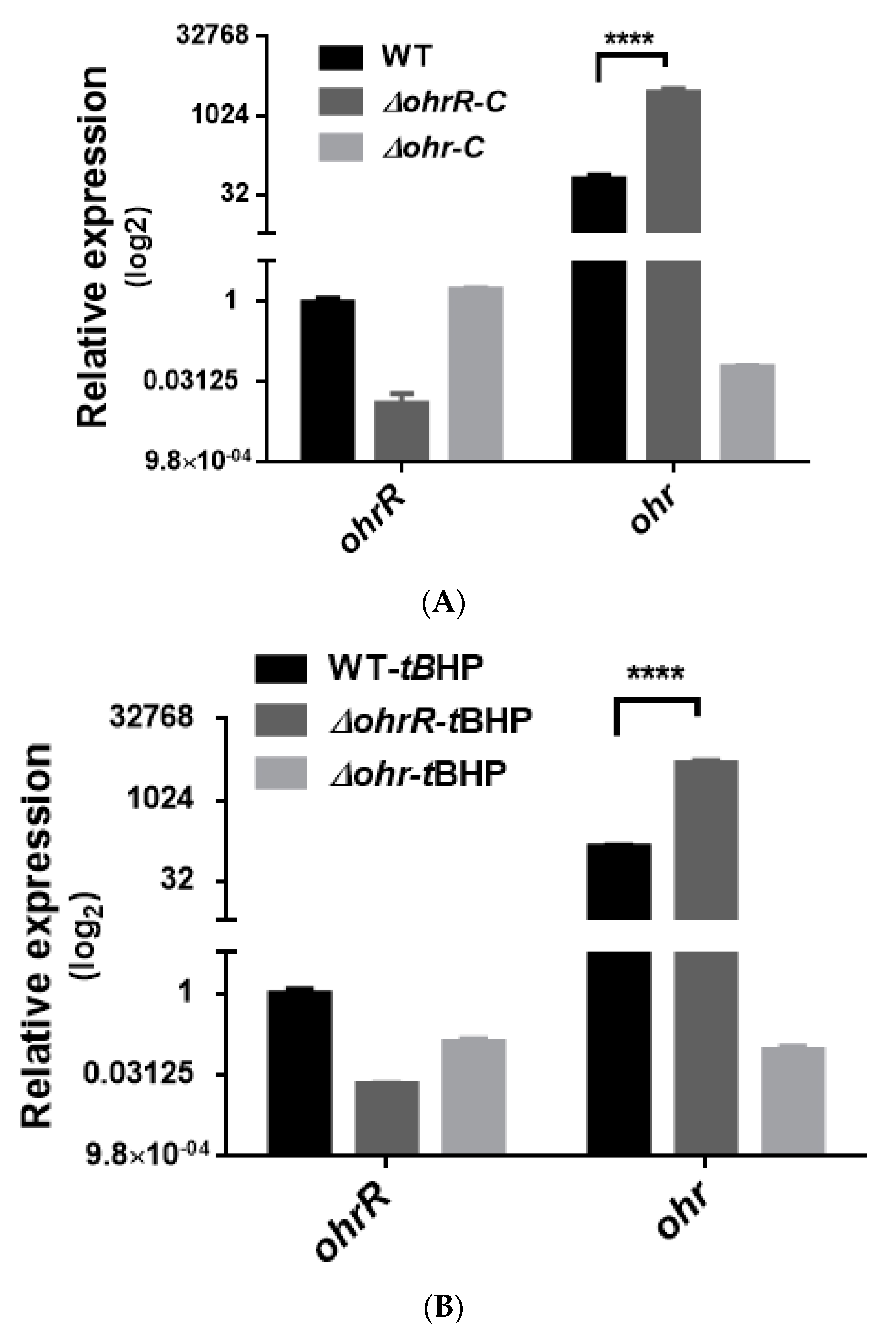
3.4. Gene Expression of Chromosomal ohr-C Can Be Induced by tBHP
3.5. Binding of OhrR-C to the Promoter of the ohr-C and ohr-p Genes
3.6. OhrR-C Suppressed Ohr-C Expression in E. coli
3.7. The ohrR-C Mutant Demonstrated Decreased Virulence in Galleria mellonella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hurst, J.K. What really happens in the neutrophil phagosome? Free Radic. Biol. Med. 2012, 53, 508–520. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C. Antimicrobial actions of reactive oxygen species. mBio 2011, 2, e00141-11. [Google Scholar] [CrossRef]
- Cussiol, J.R.; Alegria, T.G.; Szweda, L.I.; Netto, L.E. Ohr (organic hydroperoxide resistance protein) possesses a previously undescribed activity, lipoyl-dependent peroxidase. J. Biol. Chem. 2010, 285, 21943–21950. [Google Scholar] [CrossRef]
- Si, Y.; Guo, D.; Deng, S.; Lu, X.; Zhu, J.; Rao, B.; Cao, Y.; Jiang, G.; Yu, D.; Zhong, Z.; et al. Ohr and OhrR Are Critical for Organic Peroxide Resistance and Symbiosis in Azorhizobium caulinodans ORS571. Genes 2020, 11, 335. [Google Scholar] [CrossRef]
- Cussiol, J.R.; Alves, S.V.; de Oliveira, M.A.; Netto, L.E. Organic hydroperoxide resistance gene encodes a thiol-dependent peroxidase. J. Biol. Chem. 2003, 278, 11570–11578. [Google Scholar] [CrossRef]
- Fuangthong, M.; Atichartpongkul, S.; Mongkolsuk, S.; Helmann, J.D. OhrR is a repressor of ohrA, a key organic hydroperoxide resistance determinant in Bacillus subtilis. J. Bacteriol. 2001, 183, 4134–4141. [Google Scholar] [CrossRef]
- Das, K.; Garnica, O.; Dhandayuthapani, S. Utility of OhrR-Ohr system for the expression of recombinant proteins in mycobacteria and for the delivery of M. tuberculosis antigens to the phagosomal compartment. Tuberculosis 2019, 116S, S19–S27. [Google Scholar] [CrossRef]
- Will, W.R.; Fang, F.C. The evolution of MarR family transcription factors as counter-silencers in regulatory networks. Curr. Opin. Microbiol. 2020, 55, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fuangthong, M.; Helmann, J.D. The OhrR repressor senses organic hydroperoxides by reversible formation of a cysteine-sulfenic acid derivative. Proc. Natl. Acad. Sci. USA 2002, 99, 6690–6695. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Soonsanga, S.; Helmann, J.D. A complex thiolate switch regulates the Bacillus subtilis organic peroxide sensor OhrR. Proc. Natl. Acad. Sci. USA 2007, 104, 8743–8748. [Google Scholar] [CrossRef]
- Warmbold, B.; Ronzheimer, S.; Freibert, S.A.; Seubert, A.; Hoffmann, T.; Bremer, E. Two MarR-Type Repressors Balance Precursor Uptake and Glycine Betaine Synthesis in Bacillus subtilis to Provide Cytoprotection Against Sustained Osmotic Stress. Front. Microbiol. 2020, 11, 1700. [Google Scholar] [CrossRef]
- Pande, A.; Veale, T.C.; Grove, A. Gene regulation by redox-sensitive Burkholderia thailandensis OhrR and its role in bacterial killing of Caenorhabditis elegans. Infect. Immun. 2018. [CrossRef] [PubMed]
- Baumann, P.; Doudoroff, M.; Stanier, R.Y. A study of the Moraxella group. II. Oxidative-negative species (genus Acinetobacter). J. Bacteriol. 1968, 95, 1520–1541. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Bergogne-Berezin, E.; Decre, D.; Joly-Guillou, M.L. Opportunistic nosocomial multiply resistant bacterial infections—their treatment and prevention. J. Antimicrob. Chemother. 1993, 32 (Suppl. A), 39–47. [Google Scholar] [CrossRef]
- Bergogne-Berezin, E.; Towner, K.J. Acinetobacter spp. as nosocomial pathogens: Microbiological, clinical, and epidemiological features. Clin. Microbiol. Rev. 1996, 9, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Villegas, M.V.; Hartstein, A.I. Acinetobacter outbreaks, 1977–2000. Infect. Control Hosp. Epidemiol. 2003, 24, 284–295. [Google Scholar] [CrossRef]
- Bouvet, P.J.; Grimont, P.A. Taxonomy of the genus Acinetobacter with the recognition of Acinetobacter baumannii sp. nov., Acinetobacter haemolyticus sp. nov., Acinetobacter johnsonii sp. nov., and Acinetobacter junii sp. nov. and emended descriptions of Acinetobacter calcoaceticus and Acinetobacter lwoffii. Int. J. Syst. Bacteriol. 1986, 36, 228–240. [Google Scholar]
- Dorsey, C.W.; Tomaras, A.P.; Actis, L.A. Sequence and organization of pMAC, an Acinetobacter baumannii plasmid harboring genes involved in organic peroxide resistance. Plasmid 2006, 56, 112–123. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods. 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Schäfer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Beeton, M.L.; Chalker, V.J.; Maxwell, N.; Kotecha, S.; Spiller, O.B. Concurrent titration and determination of antibiotic resistance in ureaplasma species with identification of novel point mutations in genes associated with resistance. Antimicrob. Agents. Chemother. 2009, 53, 2020–2027. [Google Scholar] [CrossRef]
- Lin, H.R.; Shu, H.Y.; Lin, G.H. Biological roles of indole-3-acetic acid in Acinetobacter baumannii. Microbiol. Res. 2018, 216, 30–39. [Google Scholar] [CrossRef]
- Shu, H.Y.; Lin, L.C.; Lin, T.K.; Chen, H.P.; Yang, H.H.; Peng, K.C.; Lin, G.H. Transcriptional regulation of the iac locus from Acinetobacter baumannii by the phytohormone indole-3-acetic acid. Antonie Van Leeuwenhoek. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang-Kan, X.; Blair, J.M.A.; Chirullo, B.; Betts, J.; La Ragione, R.M.; Ivens, A.; Ricci, V.; Opperman, T.J.; Piddock, L.J.V. Lack of AcrB Efflux Function Confers Loss of Virulence on Salmonella enterica Serovar Typhimurium. mBio 2017, 8, e00968-17. [Google Scholar] [CrossRef]
- Tsai, C.J.; Loh, J.M.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Chuchue, T.; Tanboon, W.; Prapagdee, B.; Dubbs, J.M.; Vattanaviboon, P.; Mongkolsuk, S. ohrR and ohr are the primary sensor/regulator and protective genes against organic hydroperoxide stress in Agrobacterium tumefaciens. J. Bacteriol. 2006, 188, 842–851. [Google Scholar] [CrossRef]
- Oh, S.Y.; Shin, J.H.; Roe, J.H. Dual role of OhrR as a repressor and an activator in response to organic hydroperoxides in Streptomyces coelicolor. J. Bacteriol. 2007, 189, 6284–6292. [Google Scholar] [CrossRef]
- Saikolappan, S.; Das, K.; Dhandayuthapani, S. Inactivation of the organic hydroperoxide stress resistance regulator OhrR enhances resistance to oxidative stress and isoniazid in Mycobacterium smegmatis. J. Bacteriol. 2015, 197, 51–62. [Google Scholar] [CrossRef]
- Atichartpongkul, S.; Vattanaviboon, P.; Wisitkamol, R.; Jaroensuk, J.; Mongkolsuk, S.; Fuangthong, M. Regulation of Organic Hydroperoxide Stress Response by Two OhrR Homologs in Pseudomonas aeruginosa. PLoS ONE 2016, 11, e0161982. [Google Scholar] [CrossRef]
- Qi., L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed]
- Fernando, D.M.; Khan, I.U.; Patidar, R.; Lapen, D.R.; Talbot, G.; Topp, E.; Kumar, A. Isolation and Characterization of Acinetobacter baumannii Recovered from Campylobacter Selective Medium. Front. Microbiol. 2016, 7, 1871. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, M.; Moors, M.A.; Portnoy, D.A. Listeria intracellular growth and virulence require host-derived lipoic acid. Science 2003, 302, 462–464. [Google Scholar] [CrossRef]
- Allary, M.; Lu, J.Z.; Zhu, L.; Prigge, S.T. Scavenging of the cofactor lipoate is essential for the survival of the malaria parasite Plasmodium falciparum. Mol. Microbiol. 2007, 63, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.W.; Roche, H.; Trombe, M.C.; Briles, D.E.; Hakansson, A. Characterization of the dihydrolipoamide dehydrogenase from Streptococcus pneumoniae and its role in pneumococcal infection. Mol. Microbiol. 2002, 44, 431–448. [Google Scholar] [CrossRef]










| Name | Sequences (5′–3′) | Function |
|---|---|---|
| ohrR MU F | GCTGTGGGTGGATATCAGGA | Construction of ∆ohrR |
| ohrR MU R | CAGTGACTCGTCTTAAAGAT | Construction of ∆ohrR |
| pk18_ohrup_F | CGAGCTCGGTACCCGGGCGG GACAACGAATAATTTTG | Construction of ∆ohr |
| ohrup-ohrdo-R | CCAATGAAGCAAGTGAAGCA TTATTTAGCTATATTTGTACGGAGC | Construction of ∆ohr |
| ohrup-ohrdo-F | GCTCCGTACAAATATAGCTA AATAATGCTTCACTTGCTTCATTGG | Construction of ∆ohr |
| ohrdo-pK18-R | AACGACGGCCAGTGCCATTT ACGTCTTGCTGGTCGTG | Construction of ∆ohr |
| pk18_ ohrRup_F | CGAGCTCGGTACCCGGGCAT CTACCCCTTTTGGCAAT | Construction of ∆ohrR∆ohr |
| ohrRup- ohrdo-R | CCAATGAAGCAAGTGAAGCA CCTCAGAAAACTAATGGGTGCT | Construction of ∆ohrR∆ohr |
| ohr Rup- ohr do-F | AGCACCCATTAGTTTTCTGA GGTGCTTCACTTGCTTCATTGG | Construction of ∆ohrR∆ohr |
| nptII-F | ATGATTGAACAAGATGGATTGC | Amplification of kanamycin resistant gene |
| nptII-R | TCAGAAGAACTCGTCAAGAAG | Amplification of kanamycin resistant gene |
| gyrase-PF | AACCTATATTTGCTAGGGAG | Amplification of gyraeB promoter region |
| gyrase-PR | TACTAGAGGAATCATAAGCC | Amplification of gyraeB promoter region |
| ohr-1 up | ACCAATTTTTTTACAAAATT GACTTGTTAAAATAATTAAGTCGTG | Amplification of ohr-c promoter region |
| ohr-1 bo | CACGACTTAATTATTTTAAC AAGTCAATTTTGTAAAAAAATTGGT | Amplification of ohr-c promoter region |
| ohr-2 up | TACGATATATTTGCATTCAA TTTAATTTAGGAATAAAAAG | Amplification of ohr promoter region |
| ohr-2 bo | CTTTTTATTCCTAAATTAAA TTGAATGCAAATATATCGTA | Amplification of ohr-c promoter region |
| Ohr-3_PF | TTGACTTGTTAAAATAATTAAGTCG | Amplification of ohr-c promoter region |
| ohr_PF | ACCAATTTTTTTACAAAATT | Amplification of ohr-c promoter region |
| ohr_PR | CTTTTTATTCCTAAATTAAA | Amplification of ohr-C promoter region |
| pMAC_ohr_PF | GGCCGATATAAGCTCTATTT | Amplification of ohr-p promoter region |
| pMAC_ohr_PR | TGTATATTACCTTGCTTAAT | Amplification of ohr-p promoter region |
| ohrR _kpnI_flag-R | GGGGTACCCTTGTCGTCATCGTCTTTG TAGTCTTCAGTCACAATATTAAACG | Construction of pOhrR-C-Pohr-ohr-C |
| ohrR F | CGGGATCCATGGACCAAGACTGTCAAA A | Specific primer of chromosomal ohrR-C |
| ohrR R | GGGGTACCTTATTTAGCTATATTTGTAC | Specific primer of chromosomal ohrR-C |
| ohrR qF | TGGACCAAGACTGTCAAAATC | qRT-PCR primer for ohrR-C |
| ohrR qR | TCCCACAACACCAACATCAC | qRT-PCR primer for ohrR-C |
| ohr qF-2 | AAGCAACAGGTGGCCGTGAT | qRT-PCR and specific primer of chromosomal ohr-C |
| ohr qR-2 | ACCGACTTCACCTTCAACATACGC | qRT-PCR and specific primer of chromosomal ohr-C |
| ABpMAC_ohrR_F | TGTCCAAGAATCAGCTTTGCT | qRT-PCR and specific primer of ohrR-p |
| ABpMAC_ohrR_R | TTTGTCCAAGATCACCCACA | qRT-PCR and specific primer of ohrR-p |
| ABpMAC_ohr_F | AAAGGTGATGCAACGAATCC | qRT-PCR and specific primer of ohr-p |
| ABpMAC_ohr_R | GTCAAGGCAAATCCACCATT | qRT-PCR and specific primer of ohr-p |
| gyrB-F | GGCGGTTTATCTGAGTTTGT | qRT-PCR primer for gyrase gene of A. baumannii |
| gyrB-R | TTTGTGGAATGTTGTTTGTG | qRT-PCR primer for gyrase gene of A. baumannii |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-J.; Shu, H.-Y.; Lin, G.-H. Regulation of tert-Butyl Hydroperoxide Resistance by Chromosomal OhrR in A. baumannii ATCC 19606. Microorganisms 2021, 9, 629. https://doi.org/10.3390/microorganisms9030629
Chen S-J, Shu H-Y, Lin G-H. Regulation of tert-Butyl Hydroperoxide Resistance by Chromosomal OhrR in A. baumannii ATCC 19606. Microorganisms. 2021; 9(3):629. https://doi.org/10.3390/microorganisms9030629
Chicago/Turabian StyleChen, Shih-Jie, Hung-Yu Shu, and Guang-Huey Lin. 2021. "Regulation of tert-Butyl Hydroperoxide Resistance by Chromosomal OhrR in A. baumannii ATCC 19606" Microorganisms 9, no. 3: 629. https://doi.org/10.3390/microorganisms9030629
APA StyleChen, S.-J., Shu, H.-Y., & Lin, G.-H. (2021). Regulation of tert-Butyl Hydroperoxide Resistance by Chromosomal OhrR in A. baumannii ATCC 19606. Microorganisms, 9(3), 629. https://doi.org/10.3390/microorganisms9030629