
Gastrointestinal Microbiota and Parasite-Fauna of Wild Dissostichus eleginoides Smitt, 1898 Captured at the South-Central Coast of Chile

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation and Identification of Parasites
2.3. Characterization of the Parasite Community
2.4. Characterization of the Microbial Community
2.4.1. PCR-DGGE and Analysis of DGGE Profiles of the Microbial Community
2.4.2. Genomic DNA Extraction and Massive Sequencing of the Microbial Community
2.5. Data Analyses and Diversity Indices
3. Results
3.1. Parasite-Fauna Community Composition Analysis
3.1.1. Parasitic Structure
3.1.2. Parasite-Fauna Diversity Estimates
3.2. Total Microbial Community Composition Analysis
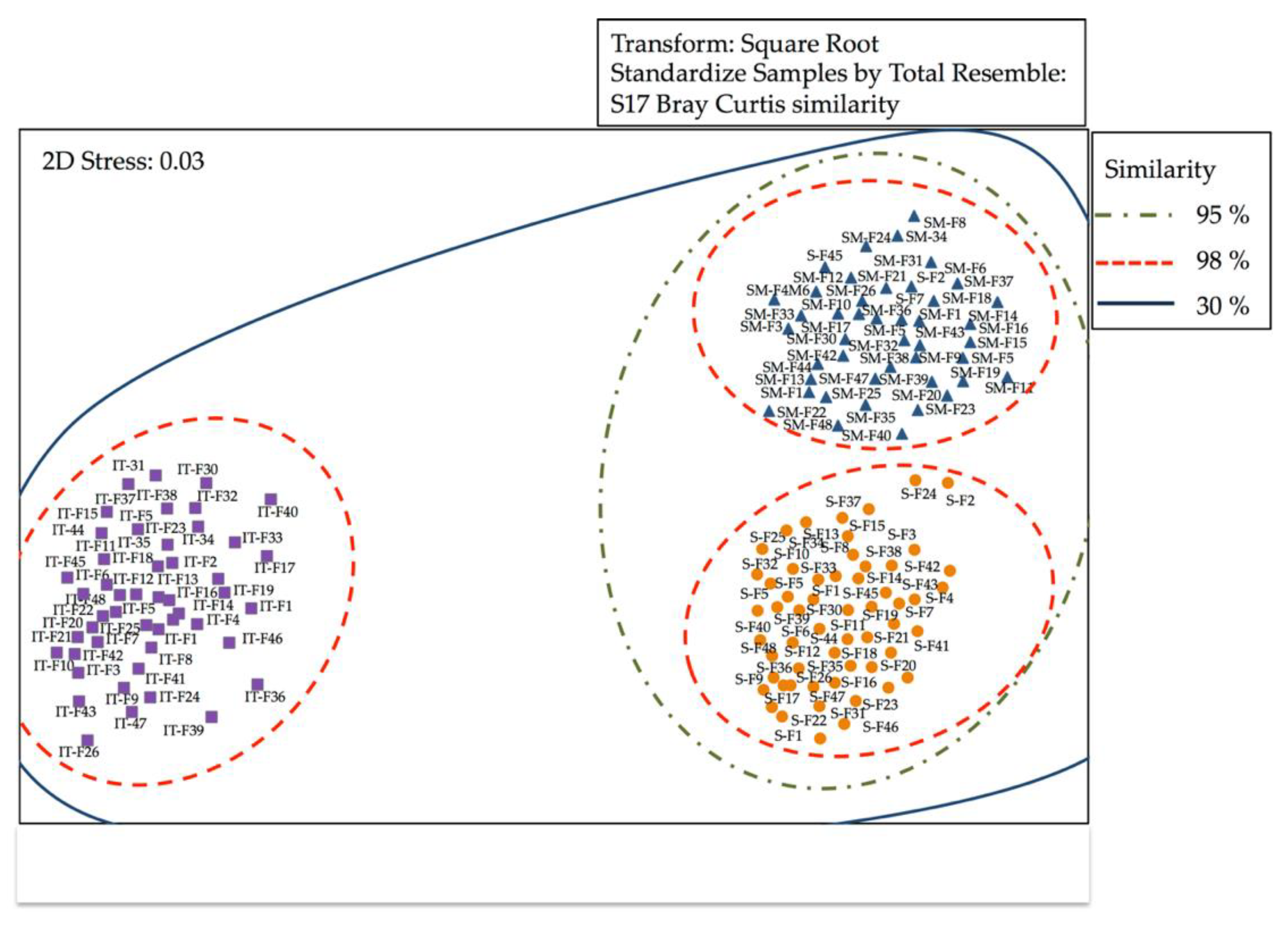
3.2.1. Analysis of Similarity of Bacterial Communities by DGGE
3.2.2. Sequencing Data and Diversity Estimates
3.2.3. Microbial Diversity Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nelson, J.; Grande, T.; Wilson, M. Fishes of the World; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Collins, M.A.; Brickle, P.; Brown, J.; Belchier, M. The Patagonian toothfish: Biology, ecology and fishery. Adv. Mar. Biol. 2010, 58, 227–300. [Google Scholar]
- Murillo, C.; Oyarzún, C.; Fernández, I. Variación latitudinal y estacional en la dieta de Dissostichus eleginoides Smitt, 1898 (Perciformes: Nototheniidae) en ambientes profundos de la costa centro-sur de Chile. Gayana 2008, 72, 94–101. [Google Scholar] [CrossRef]
- Horn, P.L. Age and growth of Patagonian toothfish (Dissostichus eleginoides) and Antarctic toothfish (D. mawsoni) in waters from the New Zealand subantarctic to the Ross Sea, Antarctica. Fish. Res. 2002, 56, 275–287. [Google Scholar] [CrossRef]
- Prenski, L.B.; Almeyda, S.M. Some biological aspects relevant to patogonian toothfish (Dissostichus eleginoides) exploitation in the Agentina exclusive economic zone and adjancent ocean sector. Frente Maritino 2000, 18, 103–124. [Google Scholar]
- FAO. Patagonian Toothfish (Dissostichus eleginoides). 2004. Available online: www.fao.org/docrep/006/y5261e/y5261e09.html (accessed on 10 March 2004).
- Young, Z.; Gili, R.; Cid, L. Prospección de Bacalao de Profundidad Entre las Latitudes 43° y 47° S. Informe Técnico; IFOP: Paris, France, 1995. [Google Scholar]
- Servicio Nacional de Pesca y Acuicultura. Control Cuota Pesquería Bacalao de Profundidad (Dissostichus eleginoides), año 2019. Informe Final. 2020. Available online: https://www.subpesca.cl/portal/616/w3-article-826.html (accessed on 3 March 2020).
- Subsecretaría de Pesca y Acuicultura. Antecedentes para la Elaboración del Plan de Manejo de las Pesquerias de Bacalao de Profundidad (Dissostichus eleginoides). 2015. Available online: https://www.subpesca.cl/portal/616/articles-103137_documento.pdf (accessed on 2 June 2015).
- Arana, P. Reproductive aspects of the Patagonian toothfish (Dissostichus eleginoides) off southern Chile. Lat. Am. J. Aquat. Res. 2009, 37, 381–394. [Google Scholar] [CrossRef]
- Sellanes, J.M.; Pedraza-García, J.; Zapata-Hernández, G. Las áreas de filtración de metano constituyen zonas de agregación del bacalao de profundidad (Dissostichus eleginoides) frente a Chile central? Lat. Am. J. Aquat. Res. 2012, 40, 980–991. [Google Scholar] [CrossRef]
- Gallardo, P. Antecedentes preliminares del cultivo de bacalao de profundidad (Dissostichus eleginoides; Nototheniidae) en la región de Magallanes, Chile. Ann. Inst. Patagon 2016, 44, 77–84. [Google Scholar] [CrossRef][Green Version]
- Canales, C.; Ferrada-Fuentes, S.; Galleguillos, R.; Oyarzún, C.; Hernández, R. Population genetic structure of Patagonian toothfish (Dissostichus eleginoides) in the Southeast Pacific and Southwest Atlantic Ocean. PeerJ 2018, 6, e4173. [Google Scholar] [CrossRef] [PubMed]
- Sáez, S.; Jaramillo, R. Estudio comparativo de escamas de las líneas laterales y corporales del Bacalao de profundidad Dissostichus eleginoides (Teleostei: Nototheniidae). Rev. Biol. Mar. Ocean 2020, 55, 142–149. [Google Scholar] [CrossRef]
- Troccoli, G.; Aguilar, E.; Martínez, P.; Belleggia, M. The diet of the Patagonian toothfish Dissostichus eleginoides, a deep-sea top predator of Southwest Atlantic Ocean. Polar Biol. 2020, 43, 1595–1604. [Google Scholar] [CrossRef]
- Urtubia, R.; Gallardo, P.; Cárdenas, C.; Lavin, P.; González-Aravena, M. First characterization of gastrointestinal culturable bacteria of Patagonian toothfish Dissostichus eleginoides (Nototheniidae). Rev. Biol. Mar. Oceanogr. 2017, 52, 399–404. [Google Scholar] [CrossRef]
- Gaevskaya, A.B.; Kovaljova, A.A.; Parukhin, A.M. Peculiarities and formation of parasitofauna of the Patagonian toothfish Dissostichus eleginoides. Biol. Morya 1990, 4, 23–28. [Google Scholar]
- Brickle, P.; Mackenzie, K.; Pike, A. Parasites of the Patagonian toothfish, Dissostichus eleginoides Smitt, 1898, in different parts of the sub-Antarctic. Pol. Biol. 2005, 28, 663–671. [Google Scholar] [CrossRef]
- Brickle, P.; Mackenzie, K.; Pike, A. Variations in the parasite fauna of the Patagonian toothfish (Dissostichus eleginoides Smitt, 1898), with length, season, and depth of hábitat around the Falkland Islands. J. Parasitol. 2006, 92, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Brickle, P.; Scott, B.E. The parasite fauna of the Patagonian toothfish Dissostichus eleginoides off the Falkland Islands. J. Helminthol. 2013, 87, 501–509. [Google Scholar] [CrossRef]
- Rodríguez, L.; George-Nascimento, M. La fauna de parásitos metazoos del bacalao de profundidad Dissostichus eleginoides Smitt, 1898 (Pisces: Nototheniidae) en Chile central: Aspectos taxonómicos, ecológicos y zoogeográficos. Rev. Chil. Hist. Nat. 1996, 69, 21–33. [Google Scholar]
- Oliva, M.; Fernández, I.; Oyarzún, C.; Murillo, C. Metazoan parasites of the stomach of Dissostichus eleginoides Smitt 1898 (Pisces: Notothenidae) from southern Chile: A tool for stock discrimination? Fish. Res. 2008, 91, 119–122. [Google Scholar] [CrossRef]
- Oyarzún, C. Catálogo de los Peces Presentes en el Sistema de Corrientes de Humboldt Frente a Chile Centro Sur; Departamento de Oceanografía, Universidad de Concepción: Concepción, Spain, 2003. [Google Scholar]
- Muñoz, V.; Dorn, L.; Reyes, H. Examen coproparasitológico. Evaluación de algunas modificaciones al método de Burrows (PAF). Parasitol. Día 1984, 8, 107–111. [Google Scholar]
- Rocka, A. Cestodes of the Antarctic fishes. Polis. Pol. Res. 2003, 24, 261–276. [Google Scholar]
- Rocka, A. Nematodes of the Antarctic fishes. Polis. Pol. Res. 2004, 25, 135–152. [Google Scholar]
- Bray, R.A.; Gibson, D.I.; Jones, A. (Eds.) Keys to the Trematoda; CABI: Wallingford, UK, 2008; Volume 3. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.C.; Price, P.W. Communities of Parasites. In Community Ecology: Patterns and Processes; Anderson, D.J., Kikkawa, J., Eds.; Blackwell Scientific Publications: Oxford, UK, 1986; pp. 187–213. [Google Scholar]
- Moreno, C.E. Métodos para Medir la Biodiversidad; M&T–Manuales y Tesis SEA: Zaragoza, Spain, 2001; Volume 1. [Google Scholar]
- Guzmán-Fierro, V.; Moraga, R.; León, C.; Campos, V.; Smith, C.; Mondaca, M. Isolation and characterization of an aerobic bacterial consortium able to degrade roxarsone. Int. J. Environ. Sci. Technol. 2015, 12, 1353–1362. [Google Scholar] [CrossRef]
- Cuevas, J.; Moraga, R.; Sánchez-Alonzo, K.; Valenzuela, C.; Aguayo, P.; Smith, C.T.; García, A.; Fernandez, Í.; Campos, V.L. Characterization of the Bacterial Biofilm Communities Present in Reverse-Osmosis Water Systems for Haemodialysis. Microorganisms 2020, 8, 1418. [Google Scholar] [CrossRef]
- Campos, V.L.; Valenzuela, C.; Yarza, P.; Kampfer, P.; Vidal, R.; Zaror, C.; Mondaca, M.A.; Lopez-Lopez, A.; Rossello-Mora, R. Pseudomonas arsenicoxydans sp. nov., an arsenite-oxidizing strain isolated from the Atacama Desert. Syst. Appl. Microbiol. 2010, 33, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, P.; Campos, V.L.; Henríquez, C.; Olivares, F.; De la Iglesia, R.; Ulloa, O.; Vargas, C.A. The Influence of pCO2-Driven Ocean Acidification on Open Ocean Bacterial Communities during A Short-Term Microcosm Experiment in the Eastern Tropical South Pacific (ETSP) off Northern Chile. Microorganisms 2020, 8, 1924. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.; Moraga, R.; Bustamante, B.; Vilo, C.; Aguayo, P.; Valenzuela, C.; Smith, C.; Yanez, J.; Fierro, G.V.; Roeckel, M.; et al. Characterization of Arsenite-Oxidizing Bacteria Isolated from Arsenic-Rich Sediments, Atacama Desert, Chile. Microorganisms 2021, 9, 483. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. Foreign: Read Data Stored by Minitab, S, SAS, SPSS, Stata, Systat, Weka, dBase, R Package Version 0.8–61. 2014. Available online: http://CRAN.R-project.org/package=foreign (accessed on 10 August 2021).
- Eastman, J.T. Antarctic Fish Biology: Evolution in a Unique Environment; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Shaw, P.W.; Arkhipkin, P.W.; Al-Khairulla, H. Genetic structuring of Patagonian toothfish populations in the Southwest Atlantic Ocean: The effect of the Antarctic Polar Front and deep-water troughs as barriers to genetic exchange. Mol. Ecol. 2004, 13, 3293–3303. [Google Scholar] [CrossRef]
- Ashford, J.; Jones, C.M.; Hofmann, E.; Everson, I.; Moreno, C.; Duhamel, G.; Williams, R. Can otoliths elemental signatures record the capture site of Patagonian toothfish (Dissostichus eleginoides), a fully marine fish in the Southern Ocean. Can. J. Fish. Aquat. Sci. 2005, 62, 2832–2840. [Google Scholar] [CrossRef]
- Laptikhovsky, V.; Arkhipkin, A.; Brickle, P. Distribution and reproduction of the Patagonian toothfish Dissostichus eleginoides Smitt around the Falkland Islands. J. Fish Biol. 2006, 68, 849–861. [Google Scholar] [CrossRef]
- Campbell, R.; Haedrich, R.; Munro, T. Parasitism and ecological relationships among deep-sea benthic fishes. Mar. Biol. 1980, 57, 301–313. [Google Scholar] [CrossRef]
- Rohde, K. Marine Parasitology; CABI Publishing: Wallingford, UK, 2005. [Google Scholar]
- Noble, E. Parasites and fishes in a deep-sea environment. Adv. Mar. Biol. 1973, 11, 121–195. [Google Scholar]
- Carvajal, J. Records of cestodes from Chilean sharks. J. Parasitol. 1974, 60, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Torres, P.; Hernández, E.; Sandoval, I. Anisakiasis and phocanemiasis in marine fishes from south of Chile. Int. J. Zoonoses 1983, 10, 146–150. [Google Scholar]
- Mercado, R.; Apt, W.; Castillo, D.; Akira, K.; Hiroshi, K.; Toshiaki, K. Hepatoxylon trichiuri. Identificación molecular de un nuevo agente de parasitosis humana en Chile. Parasitol. Lat. 2015, 64, 45–84. [Google Scholar]
- Madrid, V.; Rivera, A.; Fernández, I. Prevalencia de larvas de Anisakidae (Nematoda: Ascaridoidae) en musculatura de merluza chilena, Merluccius sp. comercializada en Concepción, Chile, en distintos períodos. Parasitol. Lat. 2016, 65, 27–31. [Google Scholar]
- Torres, P.; Jercic, M.; Weitz, J.; Dobrew, K.; Mercado, R. Human pseudoterranovosis, an emerging infection in Chile. J. Parasitol. 2007, 93, 440–443. [Google Scholar] [CrossRef]
- Solovyev, M.M.; Izvekova, G.I.; Kashinskaya, E.N.; Gisbert, E. Dependence of pH values in the digestive tract of freshwater fishes on some abiotic and biotic factors. Hydrobiologia 2018, 807, 67–85. [Google Scholar] [CrossRef]
- Ikeda-Ohtsubo, W.; Brugman, S.; Warden, C.H.; Rebel, J.M.J.; Folkerts, G.; Pieterse, C.M.J. How can we define optimal microbiota? A comparative review of structure and functions of microbiota of animals, fish, and plants in agriculture. Front. Nutr. 2018, 5, 90. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Li, L.; Huang, H.; Jiang, K.; Zhang, F.; Chen, X.; Zhao, M.; Ma, L. The Gut Microbial Community of Antarctic Fish Detected by 16S rRNA Gene Sequence Analysis. BioMed Res. Int. 2016, 3241529. [Google Scholar] [CrossRef]
- Wilkins, D.; Van Sebille, E.; Rintoul, S.R.; Lauro, F.M.; Cavicchioli, R. Advection shapes Southern Ocean microbial assemblages independent of distance and environment effects. Nat. Commun. 2013, 4, 3457. [Google Scholar] [CrossRef]
- Nurul, A.N.A.; Muhammad, D.D.; Okomoda, V.T.; Nur, A.A.B. 16S rRNA-Based metagenomic analysis of microbial communities associated with wild Labroides dimidiatus from Karah Island, Terengganu, Malaysia. Biotechnol. Rep. 2019, 21, e00303. [Google Scholar] [CrossRef] [PubMed]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquacult 2017, 467, 149–157. [Google Scholar] [CrossRef]
- Kashinskaya, E.N.; Andree, K.; Simonov, E.P.; Solovyev, M.M. DNA extraction protocols may influence biodiversity detected in the intestinal microbiome: A case study from wild Prussian carp, Carassius gibelio. FEMS Microbiol. Ecol. 2017, 93, fiw240. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Zhou, Z.; Vecino, J.L.G.; Wadsworth, S.; Rinerim, J.; Krogdahl, A.; Olsen, R.E.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nut. 2016, 22, 219–282. [Google Scholar] [CrossRef]
- Vasemägi, A.; Visse, M.; Kisand, V. Effect of environmental factors and an emerging parasitic disease on gut microbiome of wild salmonid fish. mSphere 2017, 2, e00418-17. [Google Scholar] [CrossRef] [PubMed]
- Bolnick, D.; Snowberg, L.; Hirsch, P.; Lauber, C.L.; Org, Q.; Parks, B.; Lousis, A.J.; Knight, R.; Caporaso, J.G.; Svanback, R. Individual diet has sex-dependent effects on vertebrate gut microbiota. Nat. Commun. 2014, 5, 4500. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Refaey, M.M.; Xu, W. High spatial and temporal variations of microbial community along the southern catfish gastrointestinal tract: Insights into dynamic food digestion. Front. Microbiol. 2017, 8, 1531. [Google Scholar] [CrossRef]



{kind=link}
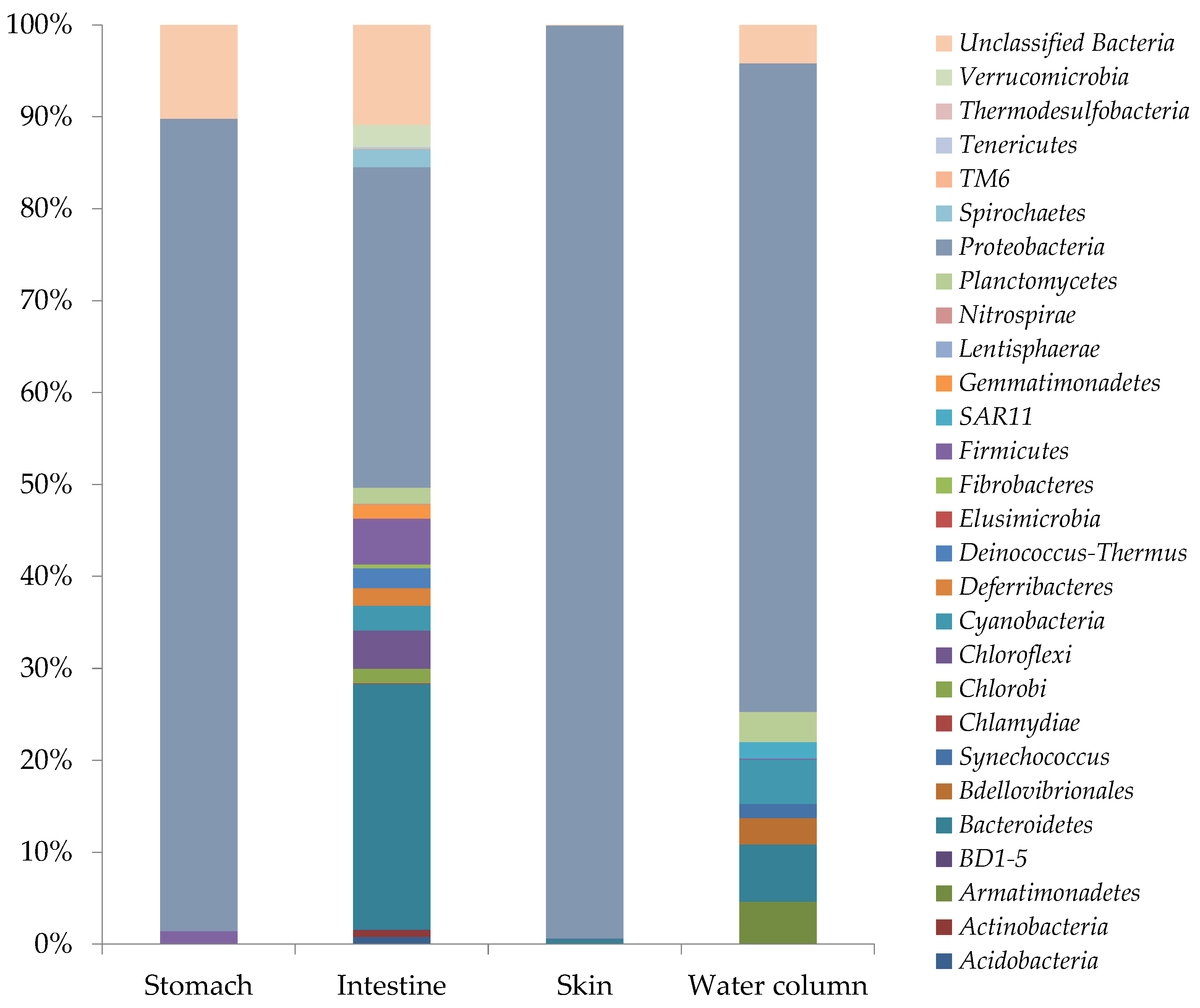{kind=link}
| Parasite | Adult/Larva | P (%) | MA | Site |
|---|---|---|---|---|
| ISOPODA | ||||
| Rocinela aff. australis | 2.12 | 0.02 | Skin | |
| NEMATODA | ||||
| Anisakis spp. | L | 100 | 29.17 | Stomach/Intestine |
| Pseudoterranova spp. | L | 19.14 | 0.51 | Stomach/Intestine |
| Hysterothylacium spp. | A | 55.31 | 3.61 | Stomach/Intestine |
| TREMATODA | ||||
| Brachyphallus crenatus | A | 68.08 | 10.72 | Stomach |
| Lecithochirium spp. | A | 82.97 | 21.23 | Stomach |
| Derogenes varicus | A | 10.63 | 0.38 | Stomach |
| Neolepidapedon spp. | A | 61.7 | 4.23 | Stomach/Intestine |
| Gonocerca spp. | A | 8.5 | 0.97 | Stomach |
| CESTODA | ||||
| Hepatoxylon trichiuri | L | 53.19 | 1.8 | Stomach/Intestine |
| Indexes | Stomach | Intestine |
|---|---|---|
| Shannon (H′) | 1.431 | 1.411 |
| Pielou (j) | 0.6514 | 0.8765 |
| Simpson (λ) | 0.7016 | 0.7445 |
| Margalef | 0.9923 | 0.7287 |
| Indexes | Stomach | Intestine | Skin |
|---|---|---|---|
| Number of high-quality reads | 4131 | 23,703 | 20,847 |
| Shannon (H′) | 0.8837 | 4.515 | 0.6971 |
| Dominance_D | 0.5921 | 0.02719 | 0.6805 |
| Equitability_J | 0.2781 | 0.7211 | 0.1689 |
| Simpson | 0.4079 | 4.515 | 0.6971 |
| Margalef | 2.86 | 5.20 | 0.61 |
| OTUs at 97% (genetic sim) | 3085 | 21,644 | 20,617 |
| Chao1 | 263.8 | 1653.1 | 149.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, I.; de Los Ríos-Escalante, P.; Valenzuela, A.; Aguayo, P.; Smith, C.T.; García-Cancino, A.; Sánchez-Alonso, K.; Oyarzún, C.; Campos, V.L. Gastrointestinal Microbiota and Parasite-Fauna of Wild Dissostichus eleginoides Smitt, 1898 Captured at the South-Central Coast of Chile. Microorganisms 2021, 9, 2522. https://doi.org/10.3390/microorganisms9122522
Fernández I, de Los Ríos-Escalante P, Valenzuela A, Aguayo P, Smith CT, García-Cancino A, Sánchez-Alonso K, Oyarzún C, Campos VL. Gastrointestinal Microbiota and Parasite-Fauna of Wild Dissostichus eleginoides Smitt, 1898 Captured at the South-Central Coast of Chile. Microorganisms. 2021; 9(12):2522. https://doi.org/10.3390/microorganisms9122522
Chicago/Turabian StyleFernández, Italo, Patricio de Los Ríos-Escalante, Ariel Valenzuela, Paulina Aguayo, Carlos T. Smith, Apolinaria García-Cancino, Kimberly Sánchez-Alonso, Ciro Oyarzún, and Víctor L. Campos. 2021. "Gastrointestinal Microbiota and Parasite-Fauna of Wild Dissostichus eleginoides Smitt, 1898 Captured at the South-Central Coast of Chile" Microorganisms 9, no. 12: 2522. https://doi.org/10.3390/microorganisms9122522
APA StyleFernández, I., de Los Ríos-Escalante, P., Valenzuela, A., Aguayo, P., Smith, C. T., García-Cancino, A., Sánchez-Alonso, K., Oyarzún, C., & Campos, V. L. (2021). Gastrointestinal Microbiota and Parasite-Fauna of Wild Dissostichus eleginoides Smitt, 1898 Captured at the South-Central Coast of Chile. Microorganisms, 9(12), 2522. https://doi.org/10.3390/microorganisms9122522