
2-Methylisoborneol (2-MIB) Excretion by Pseudanabaena yagii under Low Temperature

Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Isolation of 2-MIB Producing Cyanobacteria
2.2. Monitoring 2-MIB Production in the Freshwater Samples
2.3. DNA Extraction, PCR Amplification, and Phylogenetic Analysis
2.4. Evaluation of 2-MIB Production during Cultivation of Isolates
2.5. Whole Genome Sequencing
3. Results and Discussion
3.1. Characteristics of the Study Site
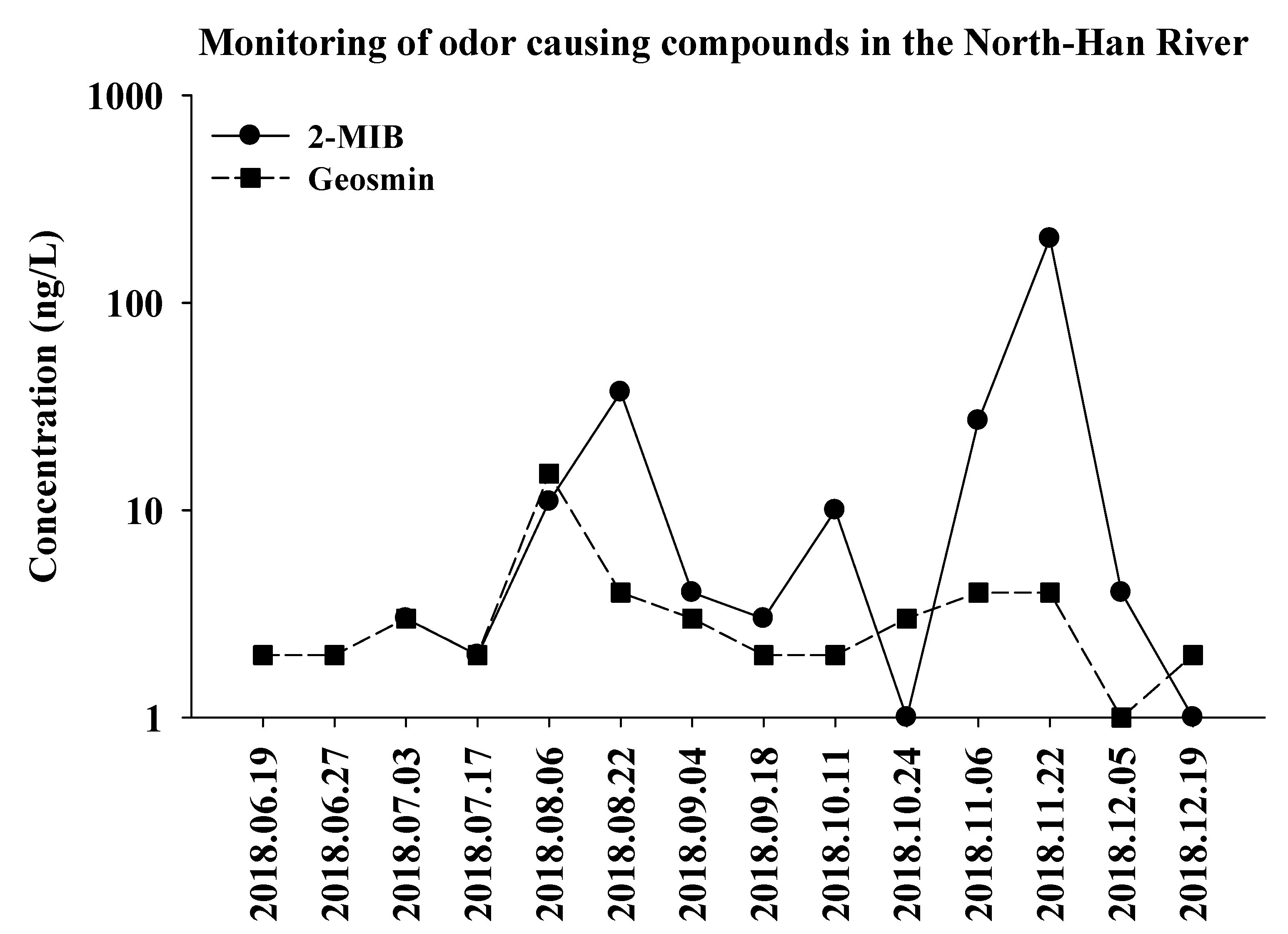
3.2. Odor Identification and Quantification in Freshwater Samples
3.3. Distribution of 2-MIB Synthesis Gene in Odor Emitting Samples
3.4. Isolation and Characterization of 2-MIB Pseudanabaena Species
3.5. Characterization of P. yagii GIHE-NHR1 2-MIB Biosynthesis Genes
3.6. Characteristics of 2-MIB Production in P. yagii GIHE-NHR1
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anuar, N.S.S.; Utsumi, M.; Sugiura, N.; Zakaria, Z.; Othman, N.; Iwamoto, K.; Goto, M.; Hara, H. Characterization of musty odor producing actinomycetes in Malaysia. In Proceedings of the International Conference on Sustainable Initiatives (ICSI 2015), Kuala Lumpur, Malaysia, 24–25 August 2015. [Google Scholar]
- Ho, L.; Hoefel, D.; Bock, F.; Saint, C.P.; Newcombe, G. Biodegradation rates of 2-methylisoborneol (MIB) and geosmin through sand filters and in bioreactors. Chemosphere 2007, 66, 2210–2218. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, K.; Dietrich, N.; Börnick, H.; Worch, E. UV-based advanced oxidation processes for the treatment of odour compounds: Efficiency and by-product formation. Water Res. 2012, 46, 5365–5373. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, S.W.; Lea, J.M.; Zimba, P.V.; Grimm, C.C. Rapid analysis of geosmin and 2-methylisoborneol in water using solid phase micro extraction procedures. Water Res. 1998, 32, 2140–2146. [Google Scholar] [CrossRef]
- Young, W.F.; Horth, H.; Crane, R.; Ogden, T.; Arnott, M. Taste and odour threshold concentrations of potential potable water contaminants. Water Res. 1996, 30, 331–340. [Google Scholar] [CrossRef]
- Giglio, S.; Chou, W.K.W.; Ikeda, H.; Cane, D.E.; Monis, P.T. Biosynthesis of 2-methylisoborneol in cyanobacteria. Environ. Sci. Technol. 2011, 45, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Jüttner, F. Physiology and biochemistry of odorous compounds from freshwater cyanobacteria and algae. Water Sci. Technol. 1995, 31, 69–78. [Google Scholar] [CrossRef]
- Asquith, E.A.; Evans, C.A.; Geary, P.M.; Dunstan, R.H.; Cole, B. The role of Actinobacteria in taste and odour episodes involving geosmin and 2-methylisoborneol in aquatic environments. J. Water Supply Res. Technol.-AQUA 2013, 62, 452–467. [Google Scholar] [CrossRef]
- Mattheis, J.P.; Roberts, R.G. Identification of geosmin as a volatile metabolite of Penicillium expansum. Appl. Environ. Microbiol. 1992, 58, 3170–3172. [Google Scholar] [CrossRef]
- Butakova, E.A. Specific features of odor-causing compounds (geosmin and 2-methylisoborneol) as secondary metabolites of cyanobacteria. Russ. J. Plant Physiol. 2013, 60, 507–510. [Google Scholar] [CrossRef]
- Izaguirre, G.; Taylor, W.D. A Pseudanabaena species from Castaic Lake, California, that produces 2-methylisoborneol. Water Res. 1998, 32, 1673–1677. [Google Scholar] [CrossRef]
- Chae, A.N.; Shin, J.W.; Cho, K.W.; Lee, B.C.; Song, K.G. Removal of geosmin and 2-methylisoborneol in drinking water by powdered activated carbon. J. Korean Soc. Civ. Eng. 2017, 37, 475–483. [Google Scholar] [CrossRef]
- Tan, F.; Chen, H.; Wu, D.; Lu, N.; Gao, Z. Optimization of Removal of 2-methylisoborneol from Drinking Water using UV/H2O2. J. Adv. Oxid. Technol. 2016, 19, 98–104. [Google Scholar] [CrossRef][Green Version]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Ömür-Özbek, P.; Little, J.C.; Dietrich, A.M. Ability of humans to smell geosmin, 2-MIB and nonadienal in indoor air when using contaminated drinking water. Water Sci. Technol. 2007, 55, 249–256. [Google Scholar] [CrossRef]
- Liu, S.; Tang, L.; Wu, M.; Fu, H.Z.; Xu, J.; Chen, W.; Ma, F. Parameters influencing elimination of geosmin and 2-methylisoborneol by K2FeO4. Sep. Purif. Technol. 2017, 182, 128–133. [Google Scholar] [CrossRef]
- Devi, A.; Chiu, Y.-T.; Hsueh, H.-T.; Lin, T.-F. Quantitative PCR based detection system for cyanobacterial geosmin/2-methylisoborneol (2-MIB) events in drinking water sources: Current status and challenges. Water Res. 2021, 188, 116478. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.-H.; Hwang, S.-J.; Kim, B.-H.; Park, J.-R.; Lee, J.-K.; Lim, B.-J. Relationship between a dense population of cyanobacteria and odorous compounds in the North Han River system in 2014 and 2015. Korean J. Ecol. Environ. 2015, 48, 263–271. [Google Scholar] [CrossRef]
- Lawton, L.A.; Codd, G.A. Cyanobacterial (blue-green algal) toxins and their significance in UK and European waters. Water Environ. J. 1991, 5, 460–465. [Google Scholar] [CrossRef]
- Park, H.K.; Jheong, W.H. Long-term changes of algal growth in Lake Paldang. J. Korean Soc. Water Qual. 2003, 19, 673–684. [Google Scholar]
- Kim, J.-A.; Kim, G.-A.; Yun, C.-J.; Park, H.-G.; Jung, E.-Y.; Cha, D.-J.; Choi, J.-T.; Son, H.-J. A study on the correlation between odorous compounds, Actinomycetes and algae in drinking water source of Nakdong river. J. Korean Soc. Environ. Eng. 2013, 35, 213–219. [Google Scholar] [CrossRef]
- Hosaka, M.; Otsuhata, M. Isolation of 2-methylisoborneol producing two species and geosmin producing one species of attached blue-green algae from Saka River. Jpn. J. Water Treat. Biol. 1988, 24, 1–11. [Google Scholar] [CrossRef][Green Version]
- Kim, Y.-J.; Youn, S.-J.; Kim, H.-N.; Hwang, M.-Y.; Park, J.-R.; Lee, B.-C.; Lee, J.-K. Formation of phytoplankton community and occurrences of odorous compounds for sediment incubation by water temperature. J. Korean Soc. Water Environ. 2015, 31, 460–467. [Google Scholar] [CrossRef][Green Version]
- Sugiura, N.; Yagi, O.; Sudo, R. Musty odor from blue-green alga, Phormidium tenue in Lake Kasumigaura. Environ. Technol. 1986, 7, 77–86. [Google Scholar]
- Kim, K.; Yoon, Y.; Cho, H.; Hwang, S.-J. Molecular Probes to Evaluate the Synthesis and Production Potential of an Odorous Compound (2-methylisoborneol) in Cyanobacteria. Int. J. Environ. Res. Public Health 2020, 17, 1933. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-T.; Yen, H.-K.; Lin, T.-F. An alternative method to quantify 2-MIB producing cyanobacteria in drinking water reservoirs: Method development and field applications. Environ. Res. 2016, 151, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gertz, E.M.; Agarwala, R.; Schäffer, A.A.; Yu, Y.-K. PSI-BLAST pseudocounts and the minimum description length principle. Nucleic Acids Res. 2009, 37, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Brambilla, E.; De Wit, R.; Wilmotte, A. Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): A morphological and molecular approach. Appl. Environ. Microbiol. 2003, 69, 5157–5169. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-Y.; Lee, S.-H.; Yun, M.-R.; Oh, S.-E.; Kim, T.-H.; Yoon, M.-H.; Park, H.-D. Draft Genome Sequence of Putative 2-Methylisoborneol-Producing Pseudanabaena yagii Strain GIHE-NHR1, Isolated from the North Han River in South Korea. Microbiol. Resour. Announc. 2020, 9, e00420–e00431. [Google Scholar] [CrossRef]
- Chin, C.-S.; Peluso, P.; Sedlazeck, F.J.; Nattestad, M.; Concepcion, G.T.; Clum, A.; Dunn, C.; O’Malley, R.; Figueroa-Balderas, R.; Morales-Cruz, A. Phased diploid genome assembly with single-molecule real-time sequencing. Nat. Methods 2016, 13, 1050–1054. [Google Scholar] [CrossRef]
- Haft, D.H.; DiCuccio, M.; Badretdin, A.; Brover, V.; Chetvernin, V.; O’Neill, K.; Li, W.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R. RefSeq: An update on prokaryotic genome annotation and curation. Nucleic Acids Res. 2018, 46, D851–D860. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Wacklin, P.; Hoffmann, L.; Komárek, J. Nomenclatural validation of the genetically revised cyanobacterial genus Dolichospermum (Ralfs ex Bornet et Flahault) comb. nova. Fottea 2009, 9, 59–64. [Google Scholar] [CrossRef]
- You, K.-A.; Byeon, M.-S.; Youn, S.-J.; Hwang, S.-J.; Rhew, D.-H. Growth characteristics of blue-green algae (Anabaena spiroides) causing tastes and odors in the North-Han River, Korea. Korean J. Ecol. Environ. 2013, 46, 135–144. [Google Scholar] [CrossRef]
- Park, M.-H.; Lim, B.-J.; Seo, W.-B.; Park, C.-H.; Kim, K.-H.; Hwang, S.-J. Akinete germination and algal growth potential test of cyanobacterium Anabaena circinalis on different waters in Lake Paldang. Korean J. Ecol. Environ. 2015, 48, 287–295. [Google Scholar] [CrossRef]
- Tuji, A.; Niiyama, Y. Two new Pseudanabaena (Cyanobacteria, Synechococcales) species from Japan, Pseudanabaena cinerea and Pseudanabaena yagii, which produce 2-methylisoborneol. Phycol. Res. 2018, 66, 291–299. [Google Scholar] [CrossRef]
- Tawong, W. Diversity of the potential 2-methylisoborneol-producing genotypes in Thai strains of Planktothricoides (Cyanobacteria). Braz. Arch. Biol. Technol. 2017, 60, e17160567. [Google Scholar] [CrossRef]
- Su, M.; Yu, J.; Zhang, J.; Chen, H.; An, W.; Vogt, R.D.; Andersen, T.; Jia, D.; Wang, J.; Yang, M. MIB-producing cyanobacteria (Planktothrix sp.) in a drinking water reservoir: Distribution and odor producing potential. Water Res. 2015, 68, 444–453. [Google Scholar] [CrossRef]
- Martin, J.F.; Izaguirre, G.; Waterstrat, P. A planktonic Oscillatoria species from Mississippi catfish ponds that produces the off-flavor compound 2-methylisoborneol. Water Res. 1991, 25, 1447–1451. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Z.; Chen, X.-P.; Zhang, D.; Zhou, J.; Wang, X.; Gao, N. The predominant phytoplankton of Pseudoanabaena holding specific biosynthesis gene-derived occurrence of 2-MIB in a drinking water reservoir. Environ. Sci. Pollut. Res. 2018, 25, 19134–19142. [Google Scholar] [CrossRef]
- Yu, G.; Zhu, M.; Chen, Y.; Pan, Q.; Chai, W.; Li, R. Polyphasic characterization of four species of Pseudanabaena (Oscillatoriales, Cyanobacteria) from China and insights into polyphyletic divergence within the Pseudanabaena genus. Phytotaxa 2015, 192, 7. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, L.; Li, L.; Song, L. 2-Methylisoborneol production characteristics of Pseudanabaena sp. FACHB 1277 isolated from Xionghe Reservoir, China. J. Appl. Phycol. 2016, 28, 3353–3362. [Google Scholar] [CrossRef]
- Kakimoto, M.; Ishikawa, T.; Miyagi, A.; Saito, K.; Miyazaki, M.; Asaeda, T.; Yamaguchi, M.; Uchimiya, H.; Kawai-Yamada, M. Culture temperature affects gene expression and metabolic pathways in the 2-methylisoborneol-producing cyanobacterium Pseudanabaena galeata. J. Plant Physiol. 2014, 171, 292–300. [Google Scholar] [CrossRef]
- Tuji, A.; Niiyama, Y. The identity and phylogeny of Pseudanabaena strain, NIES-512, producing 2-methylisoborneol (2-MIB). Bull. Natl. Mus. Nat. Sci. 2016, 42, 83–89. [Google Scholar]
- Wang, Z.; Xu, Y.; Shao, J.; Wang, J.; Li, R. Genes associated with 2-methylisoborneol biosynthesis in cyanobacteria: Isolation, characterization, and expression in response to light. PLoS ONE 2011, 6, e18665. [Google Scholar] [CrossRef]
- Komatsu, M.; Tsuda, M.; Ōmura, S.; Oikawa, H.; Ikeda, H. Identification and functional analysis of genes controlling biosynthesis of 2-methylisoborneol. Proc. Natl. Acad. Sci. USA 2008, 105, 7422–7427. [Google Scholar] [CrossRef]
- Zhou, K.; Peters, R.J. Investigating the conservation pattern of a putative second terpene synthase divalent metal binding motif in plants. Phytochemistry 2009, 70, 366–369. [Google Scholar] [CrossRef]
- Gao, J.; Zhu, J.; Wang, M.; Dong, W. Dominance and growth factors of Pseudanabaena sp. in drinking water source reservoirs, Southern China. Sustainability 2018, 10, 3936. [Google Scholar] [CrossRef]
- Wert, E.C.; Korak, J.A.; Trenholm, R.A.; Rosario-Ortiz, F.L. Effect of oxidant exposure on the release of intracellular microcystin, MIB, and geosmin from three cyanobacteria species. Water Res. 2014, 52, 251–259. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GC (7890A, Agilent) | MS (5975C, Agilent) |
|---|---|
| Column: HP-5MS | SIM Mode |
| 30 mm (L) × 0.25 mm (ID) × 0.25 μm | Selected ion |
| 2-MIB (95,108,135) | |
| Oven Temp. | |
| Initial Temp. 40 °C, Hold 5 min | |
| 1st rate 20 to 65 °C/min (5 min) | |
| 2nd rate 8 to 215 °C/min | |
| 3rd rate 30 to 300 °C/min (3 min) |
| Name | Target | Sequence (5′ to 3′) | Product Size (bp) | Annealing Temp. (°C) | Reference |
|---|---|---|---|---|---|
| MIBS02F | Monoterpene cyclase mibC | ACCTGTTACGCCACCTTCT | 307 | 63 | [26] |
| MIBS02R | CCGCAATCTGTAGCACCATG | ||||
| 16S27F | 16S rRNA + 16S-23S r/RNA ITS | AGAGTTTGATCCTGGCTCAG | 2080 | 57 | [33] |
| 23S30R | CTTCGCCTCTGTGTGCCTAGGT |
| Cyanobacterial Strain | Motif 1 | Motif 2 |
|---|---|---|
| Pseudanabaena yagii GIHE-NHR1 | -DDYYADDTE- | -NDLLSVAKD- |
| Pseudanabaena yagii NIVA-CYA 111 | -DDYYADDTE- | -NDLLSVAKD- |
| Pseudanabaena galeata NIES-512 | -DDYYADDTE- | -NDLLSVAKD- |
| Pseudanabaena sp. dqh15 | -DGYYADDTE- | -NDLLSVAKD- |
| Pseudanabaena limnetica str. Castaic Lake | -DDYYADDSE- | -NDLLSVAKD- |
| Pseudanabaena galeata NRERC-312 | -DDYYADDSE- | -NDLLSVAKD- |
| Planktothricoides raciborskii CHAB3331 | -DDYYADDSD- | -NDLLSVNKD- |
| Planktothricoides raciborskii BWN4 | -DDYYADDSD- | -NDLLSVNKD- |
| Microcoleus pseudautumnalis Ak1609 | -DDYYADDSE- | -NDLLSVNKD- |
| Oscillatoria limosa LBD | -DDYYADDSE- | -NDLLSVNKD- |
| Leptolyngbya sp. A2 | -DDYYADDSE- | -NDLLSVAKD- |
| Oscillatoria sp. 327/2 | -DDYYADDSE- | -NDLLSVAKD- |
| Planktothrix sp. 328 | -DDYYADDSE- | -NDLLSVAKD- |
| Oscillatoria prolifera OpA | -DDYYADDSE- | -NDLLSVAKD- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.-Y.; Lee, S.-H.; Yun, M.-R.; Oh, S.-E.; Lee, K.-H.; Park, H.-D. 2-Methylisoborneol (2-MIB) Excretion by Pseudanabaena yagii under Low Temperature. Microorganisms 2021, 9, 2486. https://doi.org/10.3390/microorganisms9122486
Jeong J-Y, Lee S-H, Yun M-R, Oh S-E, Lee K-H, Park H-D. 2-Methylisoborneol (2-MIB) Excretion by Pseudanabaena yagii under Low Temperature. Microorganisms. 2021; 9(12):2486. https://doi.org/10.3390/microorganisms9122486
Chicago/Turabian StyleJeong, Ju-Yong, Sang-Hoon Lee, Mi-Ra Yun, Seung-Eun Oh, Kyong-Hee Lee, and Hee-Deung Park. 2021. "2-Methylisoborneol (2-MIB) Excretion by Pseudanabaena yagii under Low Temperature" Microorganisms 9, no. 12: 2486. https://doi.org/10.3390/microorganisms9122486
APA StyleJeong, J.-Y., Lee, S.-H., Yun, M.-R., Oh, S.-E., Lee, K.-H., & Park, H.-D. (2021). 2-Methylisoborneol (2-MIB) Excretion by Pseudanabaena yagii under Low Temperature. Microorganisms, 9(12), 2486. https://doi.org/10.3390/microorganisms9122486