
Common Features and Intra-Species Variation of Cutibacterium modestum Strains, and Emended Description of the Species

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Bacterial Strains
2.2. Morphological Assessments
2.3. Selection of Genome Data
2.4. Genome Analysis
2.5. 16S rRNA Gene and recA Gene Analyses
2.6. Biochemical Profiling
2.7. MALDI-TOF Mass Spectrometry
3. Results
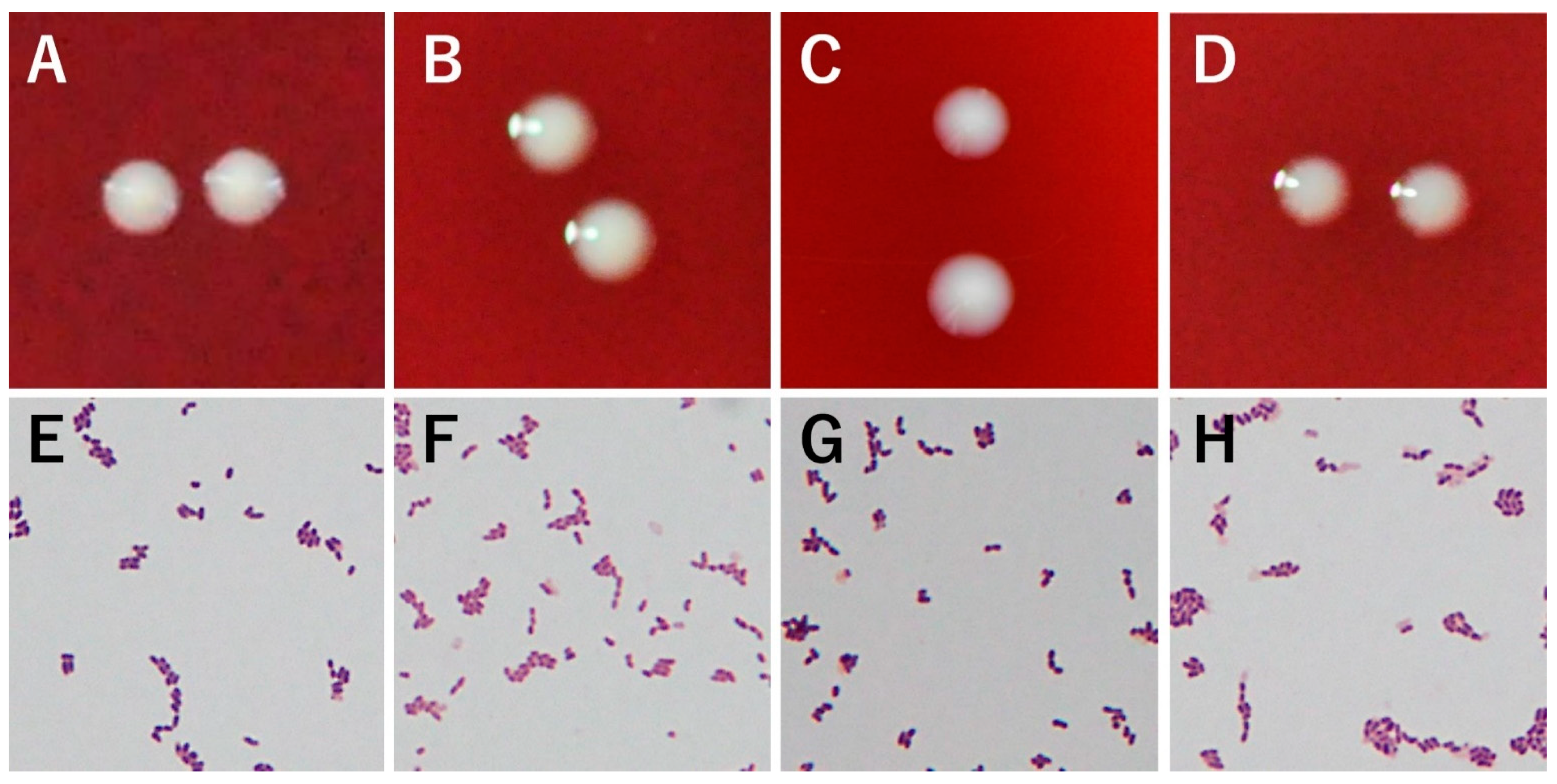
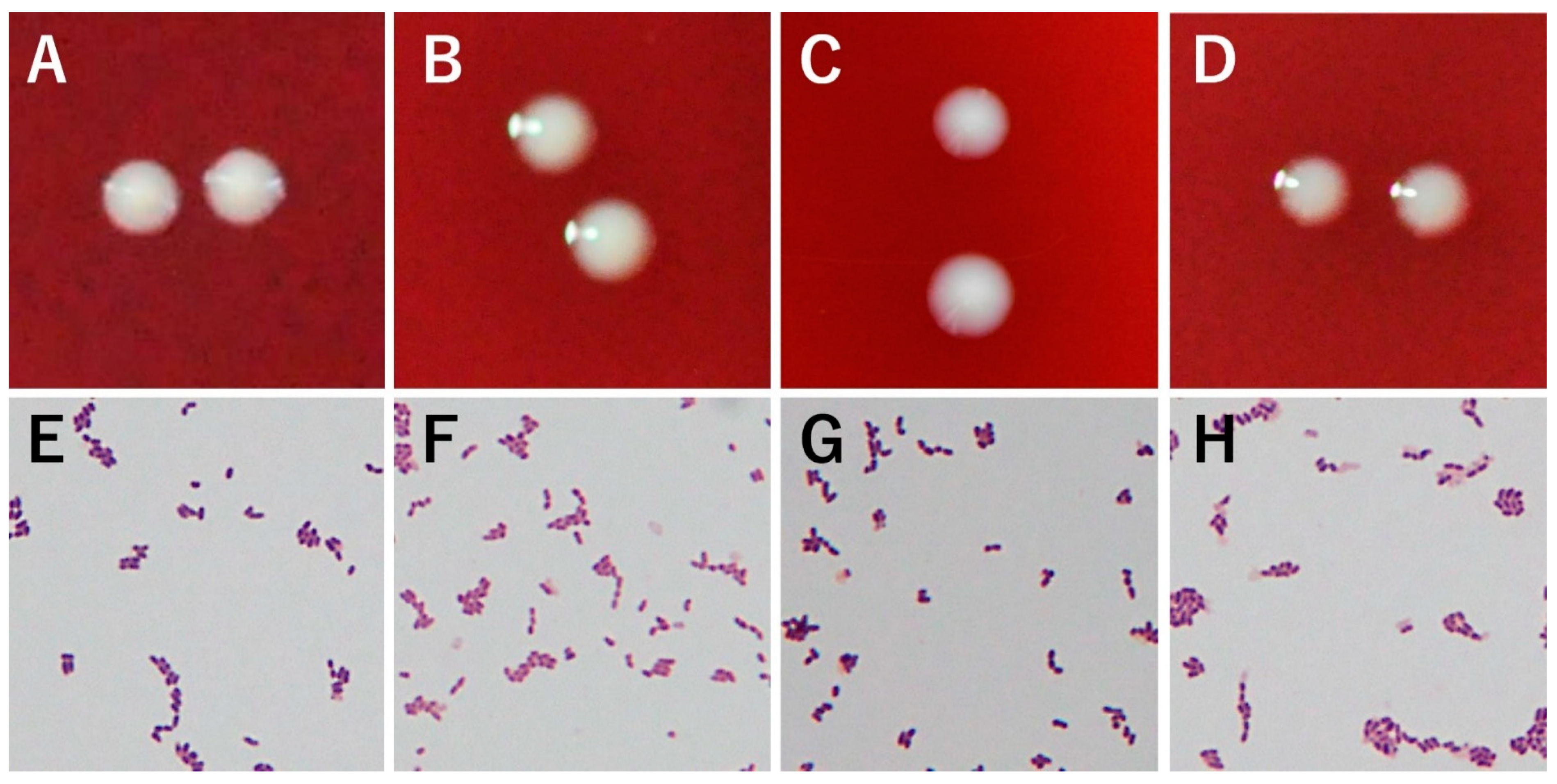
3.1. Morphology
3.2. Genome Analysis
3.3. 16S rRNA Gene and recA Gene Analyses
3.4. Biochemical Analysis
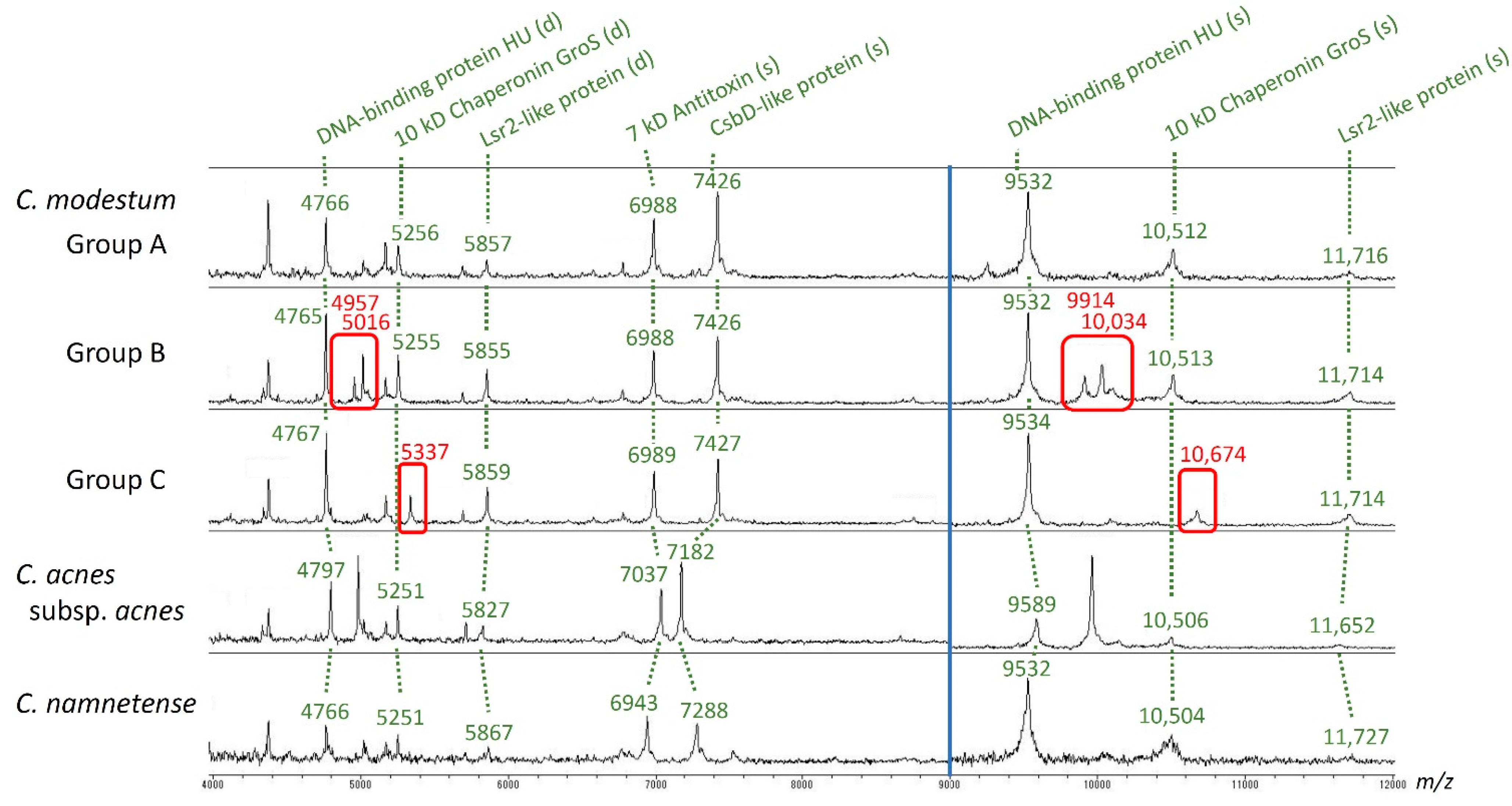
3.5. MALDI-TOF Mass Spectrometry
4. Discussion
4.1. Common Characteristics of Cutibacterium modestum
4.2. Variation of Cutibacterium modestum
4.3. “Propionibacterium humerusii” Is Included in Cutibacterium modestum
4.4. Emended Description of Cutibacterium modestum Dekio et al., 2020
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scholz, C.F.; Kilian, M. The natural history of cutaneous propionibacteria, and reclassification of selected species within the genus Propionibacterium to the proposed novel genera Acidipropionibacterium gen. nov., Cutibacterium gen. nov. and Pseudopropionibacterium gen. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4422–4432. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J. Arachnia propionica (Buchanan and Pine 1962) Pine and Georg 1969 (Approved Lists 1980), Propionibacterium propionicum corrig. (Buchanan and Pine 1962) Charfreitag et al. 1988 and Pseudopropionibacterium propionicum (Buchanan and Pine 1962) Scholz and Kilian 2016 and the nomenclatural consequences of changes in the taxonomy of the genus Propionibacterium. Int. J. Syst. Evol. Microbiol. 2019, 69, 2612–2615. [Google Scholar] [PubMed]
- Dekio, I.; Sakamoto, M.; Suzuki, T.; Yuki, M.; Kinoshita, S.; Murakami, Y.; Ohkuma, M. Cutibacterium modestum sp. nov., isolated from meibum of human meibomian glands, and emended descriptions of Cutibacterium granulosum and Cutibacterium namnetense. Int. J. Syst. Evol. Microbiol. 2020, 70, 2457–2462. [Google Scholar] [CrossRef] [PubMed]
- Dekio, I.; Asahina, A.; Shah, H.N. Unravelling the eco-specificity and pathophysiological properties of Cutibacterium species in the light of recent taxonomic changes. Anaerobe 2021, 71, 102411. [Google Scholar] [CrossRef] [PubMed]
- Butler-Wu, S.M.; Sengupta, D.J.; Kittichotirat, W.; Matsen, F.A.I.I.I.; Bumgarner, R.E. Genome sequence of a novel species, Propionibacterium humerusii. J. Bacteriol 2011, 193, 3678. [Google Scholar] [CrossRef] [PubMed]
- Dekio, I.; Culak, R.; Misra, R.; Gaulton, T.; Fang, M. Dissecting the taxonomic heterogeneity within Propionibacterium acnes: Proposal for Propionibacterium acnes subsp. acnes subsp. nov. and Propionibacterium acnes subsp. elongatum subsp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 4776–4787. [Google Scholar] [CrossRef]
- McDowell, A.; Barnard, E.; Liu, J.; Li, H.; Patrick, S. Proposal to reclassify Propionibacterium acnes type I as Propionibacterium acnes subsp. acnes subsp. nov. and Propionibacterium acnes type II as Propionibacterium acnes subsp. defendens subsp. nov. (Emendation of Propionibacterium acnes subsp. acnes (Dekio et al. 2015) and proposal of Propionibacterium acnes type II as Propionibacterium acnes subsp. defendens subsp. nov.). Int. J. Syst. Evol. Microbiol. 2016, 66, 5358–5365, Corrigendum Int. J. Syst. Evol. Microbiol. 2017, 67, 4880. [Google Scholar]
- Okuda, K.-I.; Nagahori, R.; Yamada, S.; Sugimoto, S.; Sato, C.; Sato, M.; Iwase, T.; Hashimoto, K.; Mizunoe, Y. The composition and structure of biofilms developed by Propionibacterium acnes isolated from cardiac pacemaker devices. Front. Microbiol. 2018, 9, 182. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- McDowell, A.; Nagy, I.; Magyari, M.; Barnard, E.; Patrick, S. The opportunistic pathogen Propionibacterium acnes: Insights into typing, human disease, clonal diversification and CAMP factor evolution. PLoS ONE 2013, 8, e70897. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yan, R.; Zhong, Q.; Ngo, S.; Banyagan, N.J.; Nguyen, L. The diversity and host interactions of Propionibacterium acnes bacteriophages on human skin. ISME J. 2015, 9, 2078–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T. Genome-based taxonomic classification of the phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren , A.; Garrity, G.M. List of new names and new combinations previously effectively, but not validly, published (Validation List No. 184). Int. J. Syst. Evol. Microbiol. 2018, 68, 3379–3393. [Google Scholar] [CrossRef] [PubMed]
- Dekio, I.; McDowell, A.; Sakamoto, M.; Tomida, S.; Ohkuma, M. Proposal of new combination, Cutibacterium acnes subsp. elongatum comb. nov., and emended descriptions of genus Cutibacterium, Cutibacterium acnes subsp. acnes, and Cutibacterium acnes subsp. defendens. Int. J. Syst. Evol. Microbiol. 2019, 69, 1087–1092. [Google Scholar] [PubMed]
- Aubin, G.G.; Bémer, P.; Kambalev, S.; Patel, N.B.; Lemenand, O.; Caillon, J.; Lawson, P.A.; Corvec, S. Propionibacterium namnetense sp. nov., isolated from a human bone infection. Int. J. Syst. Evol. Microbiol. 2016, 66, 3393–3399. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Suzuki, M.; Umeda, M.; Ishikawa, I.; Benno, Y. Reclassification of Bacteroides forsythus (Tanner et al. 1986) as Tannerella forsythensis corrig., gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 841–849. [Google Scholar] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Dekio, I.; Sugiura, Y.; Hamada-Tsutsumi, S.; Murakami, Y.; Tamura, H.; Suematsu, M. What do we see in spectra? Assignment of high-intensity peaks of Cutibacterium and Staphylococcus spectra of MALDI-TOF mass spectrometry by interspecies comparative proteogenomics. Microorganisms 2021, 9, 1243. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V. Complete genome sequence of DSM 30083T, the type strain (U5/41T) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand Genom. Sci. 2014, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Patrick, S.; McDowell, A. Propionibacterium. In Bergey’s Manual Trust. Bergey’s Manual of Systematics of Archaea and Bacteria, Online; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- McDowell, A.; Perry, A.L.; Lambert, P.A.; Patrick, S. A new phylogenetic group of Propionibacterium acnes. J. Med. Microbiol. 2008, 57, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, A.; Gao, A.; Barnard, E.; Fink, C.; Murray, P.I.; Dowson, C.G.; Nagy, I.; Lambert, P.A.; Patrick, S. A novel multilocus sequence typing scheme for the opportunistic pathogen Propionibacterium acnes and characterization of type I cell surface-associated antigens. Microbiology 2011, 157, 1990–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldenberger, D.; Søgaard, K.K.; Cuénod, A.; Seth-Smith, H.; Daniel de Menezes, D.; Vandamme, P.; Egli, A. Cutibacterium modestum and ‘‘Propionibacterium humerusii’’ represent the same species that is commonly misidentified as Cutibacterium acnes. Antonie van Leeuwenhoek 2021, 114, 1315–1320. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Strain Name | Origin | Country (Isolation Year) | Genome | 16S rRNA | recA | Reference |
|---|---|---|---|---|---|---|
| Cutibacterium modestum | ||||||
| (Isolates, gene sequences, and genome data used in this study) | ||||||
| M12T = JCM 33380T | meibomian gland | Japan (2013) | BJEN01 | LC466959 | (BJEN01) | [3] |
| KB17-24694 | vertebral infection | Japan (2017) | AP024747 * | LC637867 * | LC639802 * | |
| KB10376 | blood | Japan (2019) | NA | LC637868 * | LC639803 * | |
| KB11371 | blood | Japan (2019) | NA | LC637869 * | LC639804 * | |
| KB24935 | cerebrospinal fluid | Japan (2020) | NA | LC637870 * | LC639805 * | |
| JK19.3 | cardiac pacemaker device | Japan | NA | LC341281 | LC639806 * | [8] |
| HM-510 (HL037PA2) | skin | US | ADYH01 | (ADYH01) | LC639807 * /(ADYH01) | [5,10,11] |
| HM-515 (HL044PA1) | skin | US | ADZU01 | (ADZU01) | (ADZU01) | [5,10,11] |
| (Only genome data used in this study) | ||||||
| P08 | humeral bone (infected) | US | AFAM01 | (AFAM01) | (AFAM01) | [5,10] |
| T33958 | skin (wound) | US | PCZR01 | (PCZR01) | (PCZR01) | |
| Marseille-P5998 | vagina | France | UWOQ0 | (UWOQ01) | (UWOQ01) | |
| HL037PA3 | skin | US | ADXV01 | (ADXV01) | (ADXV01) | [5,10,11] |
| F0672 | oral cavity | US | CP017040 | (CP017040) | (CP017040) | |
| Cutibacterium acnes subsp. acnes | ||||||
| JCM 6425T = ATCC 6919T | skin (acne lesion) | UK | CP044255 | [6,7,12,13] | ||
| Cutibacterium acnes subsp. defendens | ||||||
| JCM 6473T = ATCC 11828T | subcutaneous abscess | unknown | CP003084 | [7,12,13] | ||
| Cutibacterium acnes subsp. elongatum | ||||||
| JCM 18919T | skin | Japan | BFFM01 | [6,14] | ||
| Cutibacterium namnetense | ||||||
| NTS 31307302T = DSM 29427T | tibial bone (infected) | France | LHWO01 | [12,13,15] | ||
| Cutibacterium avidum | ||||||
| NCTC 11864T = ATCC 25577T | unknown | unknown | AGBA01 | |||
| Cutibacterium granulosum | ||||||
| NCTC 11865T | unknown | unknown | LT906441 | |||
| Strain (Genome Entry) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | BJEN01 | 100 | 99.9 | 99.7 | 99.7 | 99.8 | 99.7 | 98.2 | 99.9 | 31.6 | 32.3 | 31.4 | 30.3 | 23.0 | 22.3 |
| 2 | AP024747 | 100 | 99.8 | 99.9 | 99.9 | 99.9 | 98.5 | 99.9 | 31.4 | 32.1 | 31.4 | 30.3 | 23.1 | 22.5 | |
| 3 | ADYH01 | 100 | 99.7 | 99.7 | 99.7 | 98.2 | 99.8 | 31.6 | 32.3 | 31.3 | 30.3 | 23.1 | 22.2 | ||
| 4 | ADZU01 | 100 | 99.7 | 99.8 | 98.1 | 99.9 | 31.6 | 32.3 | 31.4 | 30.3 | 23.1 | 23.1 | |||
| 5 | AFAM01 | 100 | 99.8 | 98.2 | 99.9 | 31.6 | 32.3 | 31.4 | 30.3 | 23.1 | 22.3 | ||||
| 6 | PCZR01 | 100 | 98.2 | 99.9 | 31.6 | 32.3 | 31.4 | 30.3 | 23.1 | 23.0 | |||||
| 7 | UWOQ01 | 100 | 98.5 | 31.5 | 32.2 | 31.4 | 30.3 | 23.0 | 22.1 | ||||||
| 8 | CP017040 | 100 | 31.4 | 32.1 | 31.4 | 30.3 | 23.1 | 22.5 | |||||||
| 9 | CP044255 | 100 | 75.5 | 73.1 | 36.0 | 23.6 | 22.5 | ||||||||
| 10 | CP003084 | 100 | 74.1 | 35.3 | 23.5 | 22.2 | |||||||||
| 11 | BFFM01 | 100 | 35.4 | 23.3 | 21.7 | ||||||||||
| 12 | LWHO01 | 100 | 23.7 | 21.8 | |||||||||||
| 13 | AGBA01 | 100 | 24.0 | ||||||||||||
| 14 | LT906441 | 100 | |||||||||||||
| Test | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| API Rapid ID 32A | ||||||||||||
| Arginine dihydrolase | − | w | w | w | − | w | w | w | − | − | − | − |
| α-Glucosidase | − | − | − | − | − | − | − | − | − | + | − | + |
| N-Acetyl-β-glucosaminidase | − | + | + | + | + | + | + | w | + | + | + | + |
| Nitrate | + | + | w | + | + | + | + | + | + | + | + | + |
| Indole | − | w | w | − | w | w | w | w | − | w | − | + |
| Proline arylamidase | + | − | + | + | + | + | + | + | + | + | + | + |
| Phenylalanine arylamidase | − | − | − | − | − | − | − | − | w | + | + | w |
| Leucine arylamidase | − | − | − | − | − | − | − | − | − | + | − | w |
| Alanine arylamidase | − | − | − | − | − | − | − | − | + | + | − | + |
| Glycine arylamidase | w | − | w | w | w | w | w | w | + | + | w | + |
| Histidine arylamidase | − | − | − | − | − | − | − | − | − | − | − | w |
| Serine arylamidase | − | − | − | − | − | − | − | − | + | + | − | + |
| API Coryne | ||||||||||||
| Pyrazinamidase | − | − | − | − | − | − | − | − | − | − | − | + |
| Pyrrolidonyl arylamidase | w | w | w | w | − | − | − | − | w | + | + | − |
| β-Galactosidase | − | − | − | − | − | − | − | − | + | + | − | + |
| Gelatin | − | − | − | − | − | − | − | − | + | + | − | + |
| Glucose | + | + | + | + | + | + | − | + | + | + | w | + |
| Ribose | − | − | − | − | − | − | − | − | + | + | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dekio, I.; Okuda, K.-i.; Nishida, M.; Hamada-Tsutsumi, S.; Suzuki, T.; Kinoshita, S.; Tamura, H.; Ohnuma, K.; Murakami, Y.; Kinjo, Y.; et al. Common Features and Intra-Species Variation of Cutibacterium modestum Strains, and Emended Description of the Species. Microorganisms 2021, 9, 2343. https://doi.org/10.3390/microorganisms9112343
Dekio I, Okuda K-i, Nishida M, Hamada-Tsutsumi S, Suzuki T, Kinoshita S, Tamura H, Ohnuma K, Murakami Y, Kinjo Y, et al. Common Features and Intra-Species Variation of Cutibacterium modestum Strains, and Emended Description of the Species. Microorganisms. 2021; 9(11):2343. https://doi.org/10.3390/microorganisms9112343
Chicago/Turabian StyleDekio, Itaru, Ken-ichi Okuda, Masako Nishida, Susumu Hamada-Tsutsumi, Tomo Suzuki, Shigeru Kinoshita, Hiroto Tamura, Kenichiro Ohnuma, Yoshiyuki Murakami, Yuki Kinjo, and et al. 2021. "Common Features and Intra-Species Variation of Cutibacterium modestum Strains, and Emended Description of the Species" Microorganisms 9, no. 11: 2343. https://doi.org/10.3390/microorganisms9112343
APA StyleDekio, I., Okuda, K.-i., Nishida, M., Hamada-Tsutsumi, S., Suzuki, T., Kinoshita, S., Tamura, H., Ohnuma, K., Murakami, Y., Kinjo, Y., & Asahina, A. (2021). Common Features and Intra-Species Variation of Cutibacterium modestum Strains, and Emended Description of the Species. Microorganisms, 9(11), 2343. https://doi.org/10.3390/microorganisms9112343