
Cardiovascular Health and The Intestinal Microbial Ecosystem: The Impact of Cardiovascular Therapies on The Gut Microbiota

 ,
,  and
and
Abstract
:1. Introduction
2. Pharmacological Therapeutics in the Treatment of CVDs
3. Gut Microbiota: Link with Cardiovascular Diseases
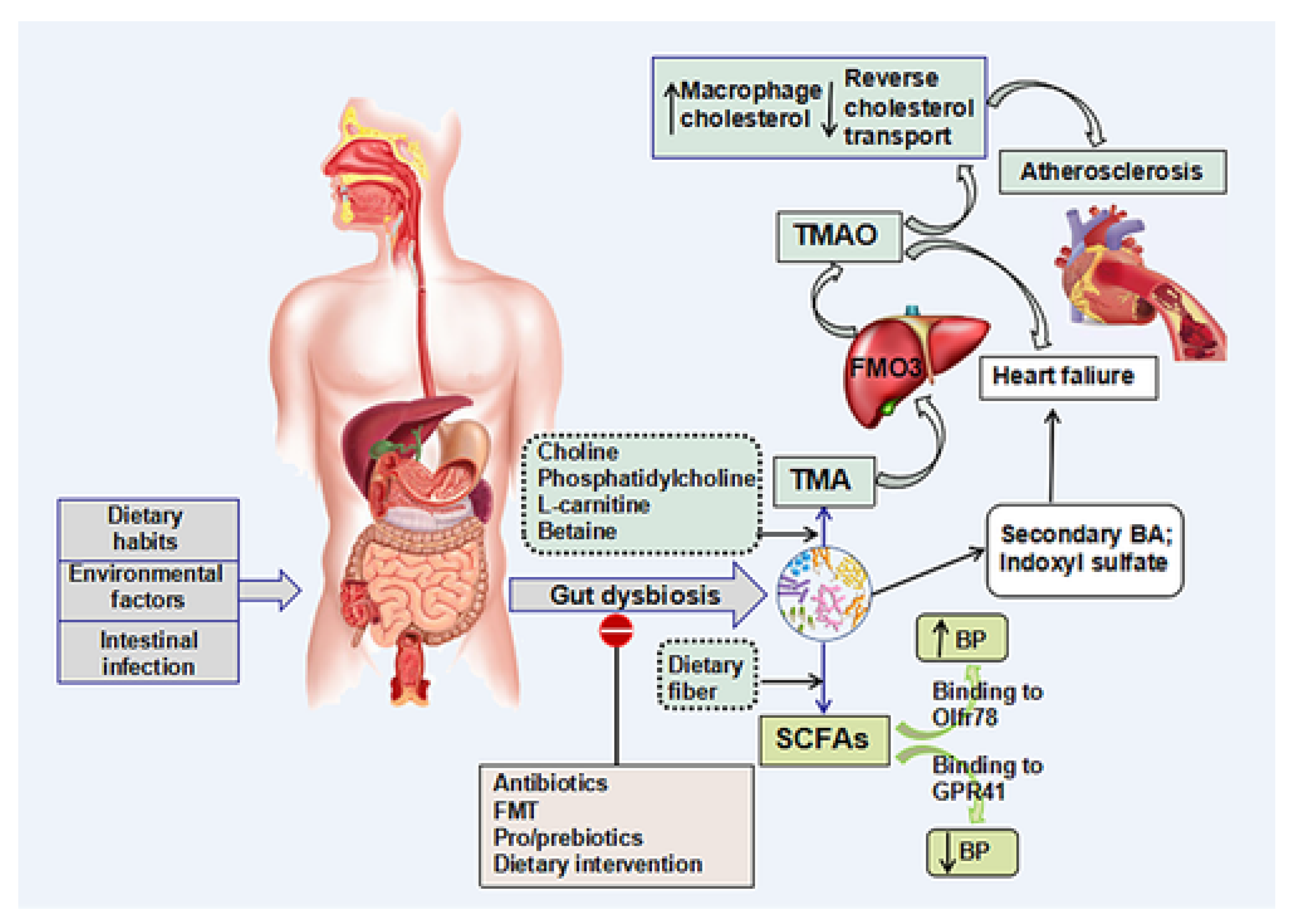
3.1. TMAO
3.2. SCFAs
3.3. Bile Acids (BA)
4. Gut Barrier Function and Bacterial Component Translocation
4.1. LPS
4.2. Peptidoglycans
5. Clinical-Trails and Animal Studies Demonstrated the Relationship between Dysbiosis and CVDs
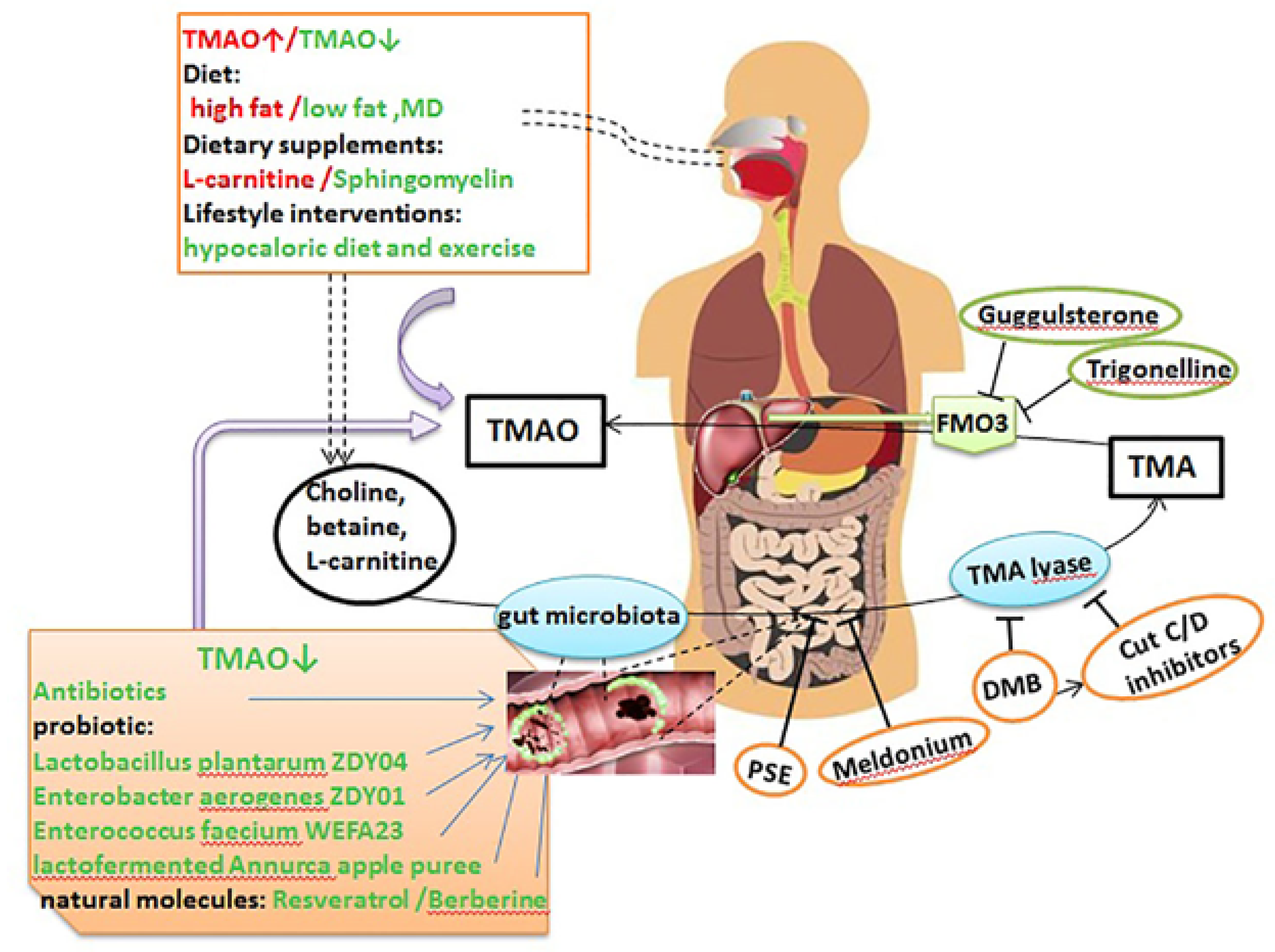
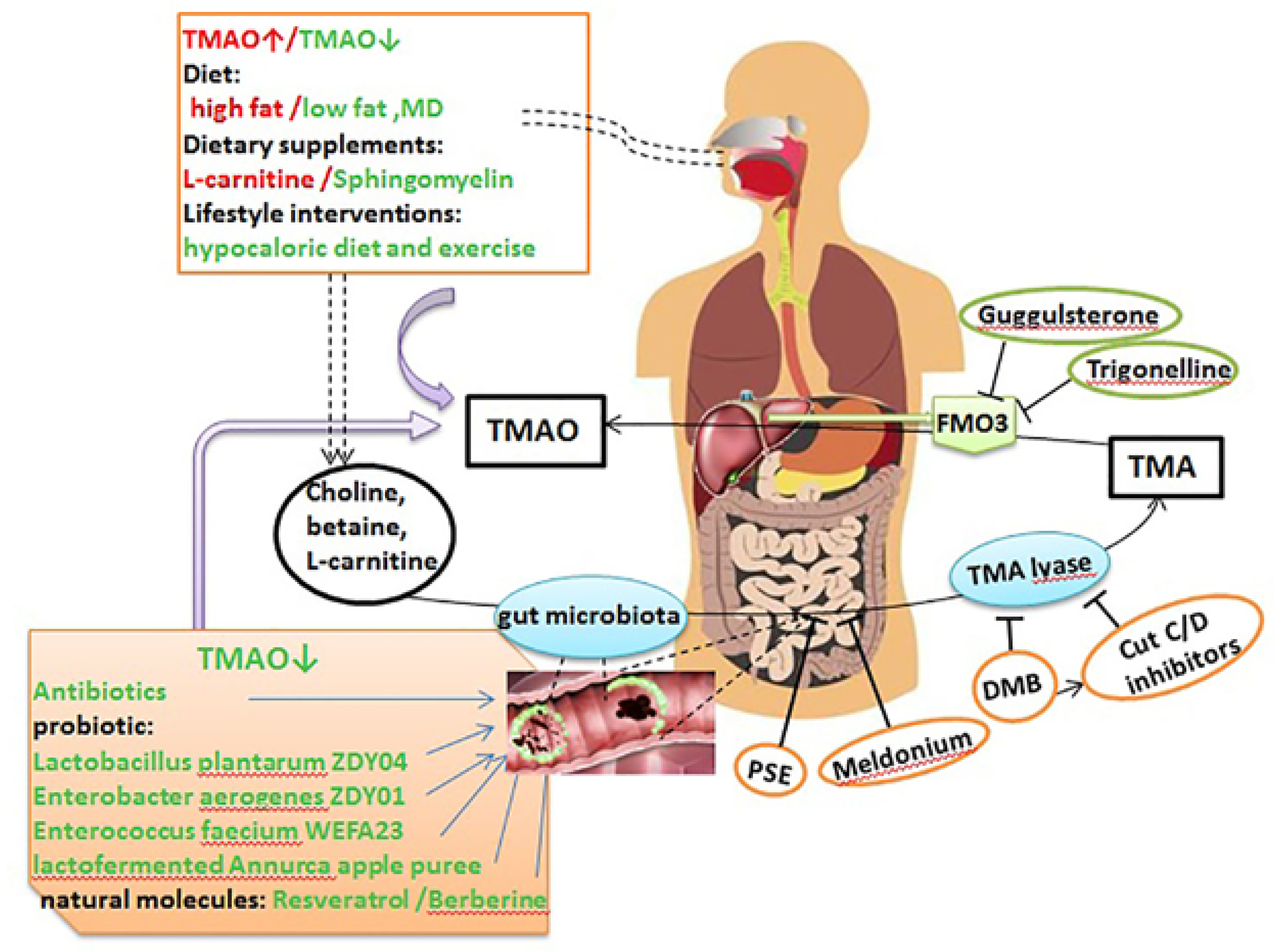
6. Therapeutic Gut-Microbiome Interaction
6.1. Antibiotics
6.2. Fecal Microbiota Transplantation (FMT)
6.3. Probiotics and Prebiotics
6.4. Dietary Intervention
6.5. Aspirin and Gut Microbiota
6.6. The Effects of Beta Blockers, ACEi, and ARBs on Gut Microbiota
6.7. Statin and Gut Microbiota
6.8. Other Therapies
7. Clinical Applications of the Drug-Gut Interactions
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Awasthi, A. Enhancing the potential preclinical and clinical benefits of quer-cetin through novel drug delivery systems. Drug Discov. Today 2020, 25, 209–222. [Google Scholar] [CrossRef]
- Roth, G.A.; Johnson, C.; Abajobir, A.; Abd-Allah, F.; Abera, S.F.; Abyu, G.; Ahmed, M.; Aksut, B.; Alam, T.; Alam, K.; et al. Global, Regional, and National Burden of Cardiovascular Diseases for 10 Causes, 1990 to 2015. J. Am. Coll. Cardiol. 2017, 70, 1–25. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Hersi, A.; Mashhoud, W.; Arafah, M.R.; Abreu, P.C.; Al Rowaily, M.A.; Al-Mallah, M.H. Cardiovascular risk factors burden in Saudi Arabia: The Africa Middle East Cardiovascular Epidemiological (ACE) study. J. Saudi Hear. Assoc. 2017, 29, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Mensah, G.A.; Roth, G.A.; Fuster, V. The Global Burden of Cardiovascular Diseases and Risk Factors: 2020 and Beyond; American College of Cardiology Foundation: Washington, DC, USA, 2019. [Google Scholar]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E.; Collins, K.J.; Himmelfarb, C.D.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, 1269–1324. [Google Scholar] [PubMed]
- Aldiab, A.; Shubair, M.M.; Al-Zahrani, J.M.; Aldossari, K.K.; Al-Ghamdi, S.; Househ, M.; Razzak, H.A.; El-Metwally, A.; Jradi, H. Prevalence of hypertension and prehypertension and its associated cardioembolic risk factors; a population based cross-sectional study in Alkharj, Saudi Arabia. BMC Public Health 2018, 18, 1327. [Google Scholar] [CrossRef] [PubMed]
- Savarese, G.; Lund, L.H. Global Public Health Burden of Heart Failure. Card. Fail. Rev. 2017, 3, 7–11. [Google Scholar] [CrossRef]
- Amitava, B.; Shanthi, M. Editorial (Heart Failure: The Need for Global Health Perspective). Curr. Cardiol. Rev. 2013, 9, 97–98. [Google Scholar]
- Scott, J. Pathophysiology and biochemistry of cardiovascular disease. Curr. Opin. Genet. Dev. 2004, 14, 271–279. [Google Scholar] [CrossRef]
- Tang, W.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Qian, Z.; Yin, J.; Xu, W.; Zhou, X. The role of intestinal microbiota in cardiovascular disease. J. Cell. Mol. Med. 2019, 23, 2343–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirtz, V.J.; Kaplan, W.; Kwan, G.; Laing, R.O. Access to Medications for Cardiovascular Diseases in Low- and Middle-Income Countries. Circulation 2016, 133, 2076–2085. [Google Scholar] [CrossRef]
- Kazemian, N.; Mahmoudi, M.; Halperin, F.; Wu, J.C.; Pakpour, S. Gut microbiota and cardiovascular disease: Opportunities and challenges. Microbiome 2020, 8, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Khursheed, R.; Singh, S.; Wadhwa, S.; Kapoor, B.; Gulati, M.; Kumar, R.; Ramanunny, A.K.; Awasthi, A.; Dua, K. Treatment strategies against diabetes: Success so far and challenges ahead. Eur. J. Pharmacol. 2019, 862, 172625. [Google Scholar] [CrossRef]
- Kasahara, K.; E Rey, F. The emerging role of gut microbial metabolism on cardiovascular disease. Curr. Opin. Microbiol. 2019, 50, 64–70. [Google Scholar] [CrossRef]
- Dannenberg, L.; Zikeli, D.; Benkhoff, M.; Ahlbrecht, S.; Kelm, M.; Levkau, B.; Polzin, A. Targeting the human microbiome and its metabolite TMAO in cardiovascular prevention and therapy. Pharmacol. Ther. 2020, 213, 107584. [Google Scholar] [CrossRef]
- Guo, F.; Zhou, J.; Li, Z.; Yu, Z.; Ouyang, D. The Association between Trimethylamine N-Oxide and Its Predecessors Choline, L-Carnitine, and Betaine with Coronary Artery Disease and Artery Stenosis. Cardiol. Res. Pract. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Ge, X.; Zheng, L.; Zhuang, R.; Yu, P.; Xu, Z.; Liu, G.; Xi, X.; Zhou, X.; Fan, H. The Gut Microbial Metabolite Trimethylamine N-Oxide and Hypertension Risk: A Systematic Review and Dose–Response Meta-analysis. Adv. Nutr. 2019, 11, 66–76. [Google Scholar] [CrossRef]
- Chen, X.-F.; Chen, X.; Tang, X. Short-chain fatty acid, acylation and cardiovascular diseases. Clin. Sci. 2020, 134, 657–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhu, Q.; Lu, A.; Liu, X.; Zhang, L.; Xu, C.; Liu, X.; Li, H.; Yang, T. Sodium butyrate suppresses angiotensin II-induced hy-pertension by inhibition of renal (pro) renin receptor and intrarenal renin–angiotensin system. J. Hypertens. 2017, 35, 1899–1908. [Google Scholar] [CrossRef]
- Roshanravan, N.; Mahdavi, R.; Alizadeh, E.; Jafarabadi, M.A.; Hedayati, M.; Ghavami, A.; Alipour, S.; Alamdari, N.M.; Barati, M.; Ostadrahimi, A. Effect of Butyrate and Inulin Supplementation on Glycemic Status, Lipid Profile and Glucagon-Like Peptide 1 Level in Patients with Type 2 Diabetes: A Randomized Double-Blind, Placebo-Controlled Trial. Horm. Metab. Res. 2017, 49, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Chen, W.-D.; Wang, Y.-D. Gut Microbiota: An Integral Moderator in Health and Disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Sayin, S.I.; Wahlström, A.; Felin, J.; Jäntti, S.; Marschall, H.-U.; Bamberg, K.; Angelin, B.; Hyötyläinen, T.; Orešič, M.; Bäckhed, F. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wostmann, B.S. Intestinal Bile Acids and Cholesterol Absorption in the Germfree Rat. J. Nutr. 1973, 103, 982–990. [Google Scholar] [CrossRef]
- Islam, K.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T.; Ogura, Y.; Hayashi, T.; Yokota, A. Bile acid is a host factor that reg-ulates the composition of the cecal microbiota in rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef]
- Shao, T.; Zhao, C.; Li, F.; Gu, Z.; Liu, L.; Zhang, L.; Wang, Y.; He, L.; Liu, Y.; Liu, Q.; et al. Intestinal HIF-1α deletion exacerbates alcoholic liver disease by inducing intestinal dysbiosis and barrier dysfunction. J. Hepatol. 2018, 69, 886–895. [Google Scholar] [CrossRef]
- Hamilton, M.K.; Boudry, G.; Lemay, D.; Raybould, H.E. Changes in intestinal barrier function and gut microbiota in high-fat diet-fed rats are dynamic and region dependent. Am. J. Physiol. Liver Physiol. 2015, 308, G840–G851. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, W.; Zuo, L.; Dong, J.; Zhu, W.; Li, Y.; Gu, L.; Gong, J.; Li, Q.; Li, N.; et al. Intestinal dysbacteriosis contributes to decreased intestinal mucosal barrier function and increased bacterial translocation. Lett. Appl. Microbiol. 2013, 58, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-Y.; Wang, M.; Zhang, J.; Barve, S.S.; McClain, C.J.; Joshi-Barve, S. Acrolein Disrupts Tight Junction Proteins and Causes Endoplasmic Reticulum Stress-Mediated Epithelial Cell Death Leading to Intestinal Barrier Dysfunction and Permeability. Am. J. Pathol. 2017, 187, 2686–2697. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.; McNabb, W.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of Tight Junction Permeability by Intestinal Bacteria and Dietary Components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Caesar, R.; Fåk, F.; Bäckhed, F. Effects of gut microbiota on obesity and atherosclerosis via modulation of inflammation and lipid metabolism. J. Intern. Med. 2010, 268, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Munford, R.S. Endotoxemia—Menace, marker, or mistake? J. Leukoc. Biol. 2016, 100, 687–698. [Google Scholar] [CrossRef]
- Laugerette, F.; Vors, C.; Peretti, N.; Michalski, M.-C. Complex links between dietary lipids, endogenous endotoxins and metabolic inflammation. Biochimie 2011, 93, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Manco, M.; Putignani, L.; Bottazzo, G.F. Gut microbiota, lipopolysaccharides, and innate immunity in the pathogenesis of obe-sity and cardiovascular risk. Endocr. Rev. 2010, 31, 817–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control meta-bolic endotoxemia-induced inflammation in high-fat diet–induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoshal, S.; Witta, J.; Zhong, J.; de Villiers, W.; Eckhardt, E. Chylomicrons promote intestinal absorption of lipopolysaccharides. J. Lipid Res. 2009, 50, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef] [Green Version]
- Kiechl, S.; Egger, G.; Mayr, M.; Wiedermann, C.J.; Bonora, E.; Oberhollenzer, F.; Muggeo, M.; Xu, Q.; Wick, G.; Poewe, W. Chronic infections and the risk of carotid atherosclerosis: Prospective results from a large population study. Circulation 2001, 103, 1064–1070. [Google Scholar] [CrossRef] [Green Version]
- Wiedermann, C.J.; Kiechl, S.; Dunzendorfer, S.; Schratzberger, P.; Egger, G.; Oberhollenzer, F.; Willeit, J. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: Prospective results from the bruneck study. J. Am. Coll. Cardiol. 1999, 34, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Niebauer, J.; Volk, H.-D.; Kemp, M.; Dominguez, M.; Schumann, R.R.; Rauchhaus, M.; Poole-Wilson, P.A.; Coats, A.J.; Anker, S.D. Endotoxin and immune activation in chronic heart failure: A prospective cohort study. Lancet 1999, 353, 1838–1842. [Google Scholar] [CrossRef]
- Rice, J.B.; Stoll, L.L.; Li, W.-G.; Denning, G.M.; Weydert, J.; Charipar, E.; Richenbacher, W.E.; Miller, F.J., Jr.; Weintraub, N.L. Low-level endotoxin induces potent inflammatory activation of human blood vessels: Inhibition by statins. Arterioscler. Throm-Bosis Vasc. Biol. 2003, 23, 1576–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoll, L.L.; Denning, G.; Li, W.-G.; Rice, J.B.; Harrelson, A.L.; Romig, S.A.; Gunnlaugsson, S.T.; Miller, F.J.; Weintraub, N.L. Regulation of Endotoxin-Induced Proinflammatory Activation in Human Coronary Artery Cells: Expression of Functional Membrane-Bound CD14 by Human Coronary Artery Smooth Muscle Cells. J. Immunol. 2004, 173, 1336–1343. [Google Scholar] [CrossRef] [Green Version]
- Philpott, D.J.; Sorbara, M.T.; Robertson, S.J.; Croitoru, K.; Girardin, S.E. NOD proteins: Regulators of inflammation in health and disease. Nat. Rev. Immunol. 2013, 14, 9–23. [Google Scholar] [CrossRef]
- Laman, J.D.; Schoneveld, A.H.; Moll, F.L.; van Meurs, M.; Pasterkamp, G. Significance of peptidoglycan, a proinflammatory bacterial antigen in atherosclerotic arteries and its association with vulnerable plaques. Am. J. Cardiol. 2002, 90, 119–123. [Google Scholar] [CrossRef]
- Serrano, M.; Moreno-Navarrete, J.M.; Puig, J.; Moreno, M.; Guerra, E.; Ortega, F.J.; Xifra, G.; Ricart, W.; Fernández-Real, J.M. Serum lipopolysaccharide-binding protein as a marker of atherosclerosis. Atherosclerosis 2013, 230, 223–227. [Google Scholar] [CrossRef]
- Lee, Y.; Wang, Z.; Lai, H.; Otto, M.D.O.; Lemaitre, R.; Fretts, A.; Sotoodehnia, N.; Budoff, M.; DiDonato, J.; McKnight, B.; et al. Longitudinal Measures of Trimethylamine N-oxide and Incident Atherosclerotic Cardiovascular Disease Events in Older Adults: The Cardiovascular Health Study. Curr. Dev. Nutr. 2020, 4, 1434. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, M. Trimethylamine N-Oxide Generated by the Gut Microbiota Is Associated with Vascular Inflammation: New Insights into Atherosclerosis. Mediat. Inflamm. 2020, 2020, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ivashkin, V.T.; Kashukh, Y.A. Impact of L-carnitine and phosphatidylcholine containing products on the proatherogenic me-tabolite TMAO production and gut microbiome changes in patients with coronary artery disease. Vopr. Pitan. 2019, 88, 25–33. [Google Scholar]
- Liu, H.-H.; Zhu, C.-G.; Cui, C.-J.; Cao, Y.-X.; Sun, D.; Wu, N.-Q.; Guo, Y.-L.; Gao, Y.; Dong, Q.-T.; Santos, R.D. Lipopolysaccharide-nuclear factor-kappa B pathway and lipoprotein apheresis effects in patients with familial hypercholesterolemia and coronary artery disease. Circulation 2020, 142, A15197. [Google Scholar] [CrossRef]
- Yoshida, N.; Yamashita, T.; Emoto, T.; Tabata, T.; Saito, Y.; Watanabe, H.; Yamada, T.; Hirata, K.-I. Abstract 10273, Bacteroides Protect Against Atherosclerosis by Regulating Gut Microbial Lipopolysaccharide Production. Circulation 2019, 140 (Suppl. 1), A10273. [Google Scholar]
- Ebner, N.; Földes, G.; Schomburg, L.; Renko, K.; Springer, J.; Jankowska, E.A.; Sharma, R.; Genth-Zotz, S.; Doehner, W.; Anker, S.D.; et al. Lipopolysaccharide responsiveness is an independent predictor of death in patients with chronic heart failure. J. Mol. Cell. Cardiol. 2015, 87, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhmoud, T.; Kumar, A.; Lo, C.-C.; Al-Sadi, R.; Clegg, S.; Alomari, I.; Zmeili, T.; Gleasne, C.D.; McMurry, K.; Dichosa, A.E.K.; et al. Investigating intestinal permeability and gut microbiota roles in acute coronary syndrome pa-tients. Hum. Microbiome J. 2019, 13, 100059. [Google Scholar] [CrossRef]
- Pastori, D.; Carnevale, R.; Nocella, C.; Novo, M.; Santulli, M.; Cammisotto, V.; Menichelli, D.; Pignatelli, P.; Violi, F. Gut-Derived Serum Lipopolysaccharide is associated with Enhanced Risk of Major Adverse Cardiovascular Events in Atrial Fibrillation: Effect of Adherence to Mediterranean Diet. J. Am. Hear. Assoc. 2017, 6, e005784. [Google Scholar] [CrossRef]
- Li, L.; Bian, T.; Lyu, J.; Cui, D.; Lei, L.; Yan, F. Human β-defensin-3 alleviates the progression of atherosclerosis accelerated by Porphyromonas gingivalis lipopolysaccharide. Int. Immunopharmacol. 2016, 38, 204–213. [Google Scholar] [CrossRef]
- Andoh, Y.; Ogura, H.; Satoh, M.; Shimano, K.; Okuno, H.; Fujii, S.; Ishimori, N.; Eshima, K.; Tamauchi, H.; Otani, T.; et al. Natural killer T cells are required for lipopolysaccharide-mediated enhancement of ather-osclerosis in apolipoprotein E-deficient mice. Immunobiology 2013, 218, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Li, Y.; Brinson, C.W.; Lopes-Virella, M.F.; Huang, Y. Cooperative stimulation of atherogenesis by lipopolysaccharide and palmitic acid-rich high fat diet in low-density lipoprotein receptor-deficient mice. Atherosclerosis 2017, 265, 231–241. [Google Scholar] [CrossRef]
- Boini, K.M.; Puchchakayala, G.; Zhang, Y.; Koka, S. TMAO Activates Carotid Endothelial Inflammasomes Leading to Enhanced Neointimal Formation in Nlrp3 Mice. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, C.; Huang, R.; Song, J.; Li, D.; Xia, M. Butyrate from pectin fermentation inhibits intestinal cholesterol absorption and attenuates atherosclerosis in apolipoprotein E-deficient mice. J. Nutr. Biochem. 2018, 56, 175–182. [Google Scholar] [CrossRef]
- Brandsma, E.; Kloosterhuis, N.J.; Koster, M.; Dekker, D.C.; Gijbels, M.J.J.; Velden Svd Ríos-Morales, M.; Faassen MJRv Loreti, M.G.; Bruin Ad Fu, J.; Kuipers, F.; Bakker, B.M.; et al. A Proinflam-matory Gut Microbiota Increases Systemic Inflammation and Accelerates Atherosclerosis. Circ. Res. 2019, 124, 94–100. [Google Scholar] [CrossRef]
- Kappel, B.A.; De Angelis, L.; Heiser, M.; Ballanti, M.; Stoehr, R.; Goettsch, C.; Mavilio, M.; Artati, A.; Paoluzi, O.A.; Adamski, J.; et al. Cross-omics analysis revealed gut microbiome-related metabolic pathways underlying atherosclerosis development after antibiotics treatment. Mol. Metab. 2020, 36, 100976. [Google Scholar] [CrossRef]
- Liu, M.; Han, Q.; Yang, J. Trimethylamine-N-oxide (TMAO) increased aquaporin-2 expression in spontaneously hypertensive rats. Clin. Exp. Hypertens. 2018, 41, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; He, Q.; Wang, F.; Schulte, C.; Moore, V.; Redington, A.N. Abstract 15759, Remote Ischemic Preconditioning Attenuates Lipopolysaccharide-Induced Septic Cardiomyopathy by Regulating Circulating Inflammatory Mediators. Circulation 2018, 138 (Suppl. 1), A15759. [Google Scholar]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; A Koeth, R.; Levison, B.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeth, R.A.; Lam-Galvez, B.R.; Kirsop, J.; Wang, Z.; Levison, B.S.; Gu, X.; Copeland, M.F.; Bartlett, D.; Cody, D.B.; Dai, H.J.; et al. l-Carnitine in omnivorous diets induces an atherogenic gut microbial pathway in humans. J. Clin. Investig. 2018, 129, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphati-dylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lever, M.; George, P.M.; Slow, S.; Bellamy, D.; Young, J.M.; Ho, M.; McEntyre, C.J.; Elmslie, J.L.; Atkinson, W.; Molyneux, S.L. Betaine and trimethylamine-N-oxide as predictors of cardiovascular outcomes show different patterns in diabetes mellitus: An observa-tional study. PLoS ONE 2014, 9, e114969. [Google Scholar]
- Mente, A.; Chalcraft, K.; Ak, H.; Davis, A.D.; Lonn, E.; Miller, R.; Potter, M.A.; Yusuf, S.; Anand, S.S.; McQueen, M.J. The Relationship Between Trimethylamine-N-Oxide and Prevalent Cardiovascular Disease in a Multiethnic Population Living in Canada. Can. J. Cardiol. 2015, 31, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, W.H.W.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur. Heart J. 2014, 35, 904–910. [Google Scholar] [CrossRef]
- Klinder, A.; Shen, Q.; Heppel, S.; Lovegrove, J.A.; Rowland, I.; Tuohy, K. Impact of increasing fruit and vegetables and flavonoid intake on the human gut microbiota. Food Funct. 2016, 7, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, J. TMAO and heart disease: The new red meat risk? JAMA 2019, 321, 2149–2151. [Google Scholar] [CrossRef]
- Heianza, Y.; Ma, W.; Manson, J.E.; Rexrode, K.M.; Qi, L. Gut microbiota metabolites and risk of major adverse cardiovascular dis-ease events and death: A systematic review and meta-analysis of prospective studies. J. Am. Heart Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; You, T.; Li, X.; Pan, T.; Xiang, L.; Han, Y.; Zhu, L. Circulating trimethylamine N-oxide and the risk of cardiovascular diseases: A systematic review and meta-analysis of 11 prospective cohort studies. J. Cell. Mol. Med. 2017, 22, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.-E.; Liao, P.-D.; Zhao, X.-J.; Wang, L. Trimethylamine-N-oxide has prognostic value in coronary heart disease: A me-ta-analysis and dose-response analysis. BMC Cardiovasc. Disord. 2020, 20, 7. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. L-Carnitine Consumption, Its Metabolism by Intestinal Microbiota, and Cardiovascular Health. Mayo Clin. Proc. 2013, 88, 786–789. [Google Scholar] [CrossRef] [Green Version]
- Landfald, B.; Valeur, J.; Berstad, A.; Raa, J. Microbial trimethylamine-N-oxide as a disease marker: Something fishy? Microb. Ecol. Health Dis. 2017, 28, 1327309. [Google Scholar] [CrossRef] [Green Version]
- Ussher, J.R.; Lopaschuk, G.D.; Arduini, A. Gut microbiota metabolism of L-carnitine and cardiovascular risk. Atherosclerosis 2013, 231, 456–461. [Google Scholar] [CrossRef]
- Meyer, K.A.; Benton, T.Z.; Bennett, B.J.; Jacobs, D.R., Jr.; Lloyd-Jones, D.M.; Gross, M.D.; Carr, J.J.; Gordon-Larsen, P.; Zeisel, S.H. Microbiota-dependent metabolite trimethylamine N-oxide and coronary artery calcium in the coronary artery risk development in young adults study (CARDIA). J. Am. Heart Assoc. 2016, 5, e003970. [Google Scholar] [CrossRef] [Green Version]
- Huart, J.; Leenders, J.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.-M.; Melin, P.; De Tullio, P.; Jouret, F. Gut mi-crobiota and fecal levels of short-chain fatty acids differ upon 24-hour blood pressure levels in men. Hypertension 2019, 74, 1005–1013. [Google Scholar] [CrossRef]
- Lam, O.L.; Zhang, W.; Samaranayake, L.P.; Li, L.S.; McGrath, C.P.J. A systematic review of the effectiveness of oral health promotion activities among patients with cardiovascular disease. Int. J. Cardiol. 2010, 151, 261–267. [Google Scholar] [CrossRef]
- Renvert, S.; Pettersson, T.; Ohlsson, O.; Persson, G.R. Bacterial Profile and Burden of Periodontal Infection in Subjects with a Diagnosis of Acute Coronary Syndrome. J. Periodontol. 2006, 77, 1110–1119. [Google Scholar] [CrossRef]
- Pussinen, P.J.; Alfthan, G.; Rissanen, H.; Reunanen, A.; Asikainen, S.; Knekt, P. Antibodies to Periodontal Pathogens and Stroke Risk. Stroke 2004, 35, 2020–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, J.D.; Eke, P.; Heiss, G.; Madianos, P.; Couper, D.; Lin, D.; Moss, K.; Elter, J.; Offenbacher, S. Periodontal disease and coronary heart disease: A reappraisal of the exposure. Circulation 2005, 112, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Li, X.; Yang, F.; Zhao, R.; Pan, X.; Liang, J.; Tian, L.; Li, X.; Liu, L.; Xing, Y.; et al. Gut Microbiota-Dependent Marker TMAO in Promoting Cardiovascular Disease: Inflammation Mechanism, Clinical Prognostic, and Potential as a Therapeutic Target. Front. Pharmacol. 2019, 10, 1360. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.; Ahrné, S.; Molin, G.; Berggren, A.; Palmquist, I.; Fredrikson, G.N.; Jeppsson, B. Probiotic therapy to men with incipient arteriosclerosis initiates increased bacterial diversity in colon: A randomized controlled trial. Atherosclerosis 2010, 208, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, I.F. Microbial-dependent TMAO as a prognostic marker in ACS. Nat. Rev. Cardiol. 2017, 14, 128–129. [Google Scholar] [CrossRef]
- Organ, C.L.; Otsuka, H.; Bhushan, S.; Wang, Z.; Bradley, J.; Trivedi, R.; Polhemus, D.J.; Tang, W.W.; Wu, Y.; Hazen, S.L. Choline diet and its gut microbe–derived metabolite, trimethylamine N-oxide, exacerbate pressure overload–induced heart failure. Circ. Heart Fail. 2016, 9, e002314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, O.; Spor, A.; Felin, J.; Fak, F.; Stombaugh, J.; Tremaroli, V.; Behre, C.J.; Knight, R.; Fagerberg, B.; Ley, R.E.; et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc. Natl. Acad. Sci. USA 2010, 108, 4592–4598. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.J.; El Mokhtari, N.E.; Musfeldt, M.; Hellmig, S.; Freitag, S.; Rehman, A.; Kühbacher, T.; Nikolaus, S.; Namsolleck, P.; Blaut, M.; et al. Detection of Diverse Bacterial Signatures in Atherosclerotic Lesions of Patients with Coronary Heart Disease. Circulation 2006, 113, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Mattila, K.J.; Nieminen, M.S.; Valtonen, V.V.; Rasi, V.P.; Kesäniemi, Y.A.; Syrjälä, S.L.; Jungell, P.S.; Isoluoma, M.; Hietaniemi, K.; Jokinen, M.J. Association between dental health and acute myocardial infarction. Br. Med. J. 1989, 298, 779–781. [Google Scholar] [CrossRef] [Green Version]
- Hyvärinen, K.; Mäntylä, P.; Buhlin, K.; Paju, S.; Nieminen, M.S.; Sinisalo, J.; Pussinen, P.J. A common periodontal pathogen has an adverse association with both acute and stable coronary artery disease. Atherosclerosis 2012, 223, 478–484. [Google Scholar] [CrossRef]
- Fåk, F.; Tremaroli, V.; Bergström, G.; Bäckhed, F. Oral microbiota in patients with atherosclerosis. Atherosclerosis 2015, 243, 573–578. [Google Scholar] [CrossRef] [PubMed]
- e Silva Filho, W.S.; Casarin, R.C.; Junior, E.L.N.; Passos, H.M.; Sallum, A.W.; Gonçalves, R.B. Microbial diversity similarities in perio-dontal pockets and atheromatous plaques of cardiovascular disease patients. PLoS ONE 2014, 9, e109761. [Google Scholar]
- Jonsson, A.L.; Bäckhed, F. Role of gut microbiota in atherosclerosis. Nat. Rev. Cardiol. 2016, 14, 79–87. [Google Scholar] [CrossRef]
- Karlsson, F.; Fåk, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Bäckhed, F.; Nielsen, J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef] [Green Version]
- Dziubak, A.; Wójcicka, G.; Wojtak, A.; Bełtowski, J. Metabolic Effects of Metformin in the Failing Heart. Int. J. Mol. Sci. 2018, 19, 2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCreight, L.J.; Bailey, C.J.; Pearson, E.R. Metformin and the gastrointestinal tract. Diabetologia 2016, 59, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, S.; Ferguson, J.F. Gut Microbiome and Response to Cardiovascular Drugs. Circ. Genom. Precis. Med. 2019, 12, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Haiser, H.J.; Seim, K.L.; Balskus, E.P.; Turnbaugh, P.J. Mechanistic insight into digoxin inactivation by Eggerthella lenta augments our understanding of its pharmacokinetics. Gut Microbes 2014, 5, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; He, J.; Jiaojiao, H. The influence of gut microbiota on drug metabolism and toxicity. Expert Opin. Drug Metab. Toxicol. 2015, 12, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhang, J.; Zhao, A.; Li, W.; Feng, Q.; Wang, R. Effects of intestinal flora on the pharmacokinetics and pharmacodynamics of aspirin in high-altitude hypoxia. PLoS ONE 2020, 15, e0230197. [Google Scholar] [CrossRef]
- Gulati, M.; Singh, S.K.; Corrie, L.; Chandwani, L.; Singh, A.; Kapoor, B.; Kumar, R.; Pandey, N.K.; Kumar, B.; Awasthi, A.; et al. Fecal Microbiota Transplant: Latest Addition to Arsenal Against Recurrent Clostridium Difficile Infection. Recent Pat. Anti-Infect. Drug Discov. 2020, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cole-Jeffrey, C.T.; Liu, M.; Katovich, M.J.; Raizada, M.K.; Shenoy, V. ACE2 and microbiota: Emerging targets for cardiopulmonary disease therapy. J. Cardiovasc. Pharmacol. 2015, 66, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamo, T.; Akazawa, H.; Suda, W.; Saga-Kamo, A.; Shimizu, Y.; Yagi, H.; Liu, Q.; Nomura, S.; Naito, A.T.; Takeda, N.; et al. Dysbiosis and compositional alterations with aging in the gut microbiota of patients with heart failure. PLoS ONE 2017, 12, e0174099. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P.; Bienenstock, J.; Kunze, W.A. Vagal pathways for microbiome-brain-gut axis communication. Microb. Endocrinol. Microbiota-Gut-Brain Axis Health Dis. 2014, 817, 115–133. [Google Scholar]
- Moludi, J.; Khedmatgozar, H.; Nachvak, S.M.; Abdollahzad, H.; Moradinazar, M.; Tabaei, A.S. The effects of co-administration of probiotics and prebiotics on chronic inflammation, and depression symptoms in patients with coronary artery diseases: A randomized clinical trial. Nutr. Neurosci. 2021, 1–10. [Google Scholar] [CrossRef]
- Thushara, R.M.; Gangadaran, S.; Solati, Z.; Moghadasian, M.H. Cardiovascular benefits of probiotics: A review of experimental and clinical studies. Food Funct. 2016, 7, 632–642. [Google Scholar] [CrossRef]
- Yang, F.; Chen, H.; Gao, Y.; An, N.; Li, X.; Pan, X.; Yang, X.; Tian, L.; Sun, J.; Xiong, X.; et al. Gut microbiota-derived short-chain fatty acids and hypertension: Mechanism and treatment. Biomed. Pharmacother. 2020, 130, 110503. [Google Scholar] [CrossRef]
- Chen, M.-L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.-D.; Zhang, Q.-Y.; Mi, M.-T. Resveratrol Attenuates Trimethyla-mine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. mBio 2016, 7, e02210–e02215. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-J.; Tran, T.T. Current Studies of Aspirin as an Anticancer Agent and Strategies to Strengthen its Therapeutic Application in Cancer. Curr. Pharm. Des. 2021, 27, 2209–2220. [Google Scholar] [CrossRef]
- Kim, I.S.; Yoo, D.-H.; Jung, I.-H.; Lim, S.; Jeong, J.-J.; Kim, K.-A.; Bae, O.-N.; Yoo, H.H.; Kim, D.-H. Reduced metabolic activity of gut mi-crobiota by antibiotics can potentiate the antithrombotic effect of aspirin. Biochem. Pharmacol. 2016, 122, 72–79. [Google Scholar] [CrossRef]
- Mortensen, B.; Murphy, C.; O’Grady, J.; Lucey, M.; Elsafi, G.; Barry, L.; Westphal, V.; Wellejus, A.; Lukjancenko, O.; Eklund, A.C.; et al. Bifidobacterium breve Bif195 Protects Against Small-Intestinal Damage Caused by Acetylsalicylic Acid in Healthy Volunteers. Gastroenterology 2019, 157, 637–646.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, T.J.; Ahmed, Y.M.; Zamzami, M.A.; Siddiqui, A.M.; Khan, I.; Baothman, O.A.; Mehanna, M.G.; Kuerban, A.; Kaleemuddin, M.; Yasir, M. Atorvastatin Treatment Modulates the Gut Microbiota of the Hypercholesterolemic Patients. OMICS A J. Integr. Biol. 2018, 22, 154–163. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; De La Visitación, N.; Aguilera-Sánchez, N.; Redondo, J.M.; Duarte, J. Protective Effects of Short-Chain Fatty Acids on Endothelial Dysfunction Induced by Angiotensin II. Front. Physiol. 2020, 11, 277. [Google Scholar] [CrossRef] [Green Version]
- Fennema, D.; Phillips, I.R.; Shephard, E.A. Trimethylamine and Trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-Mediated Host-Microbiome Metabolic Axis Implicated in Health and Disease. Drug Metab. Dispos. 2016, 44, 1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, J.; Griffin, B.T.; Clarke, G.; Hyland, N.P. Drug-gut microbiota interactions: Implications for neuropharmacology. Br. J. Pharmacol. 2018, 175, 4415–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthrie, L.; Kelly, L. Bringing microbiome-drug interaction research into the clinic. EBioMedicine 2019, 44, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Shi, X.; Yang, J.; Zhao, Y.; Xue, L.; Xu, L.; Cai, J. Gut microbes in cardiovascular diseases and their potential therapeutic applications. Protein Cell 2020, 12, 346–359. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Therapeutic Group According to EML | International Non-Proprietary Name (INN) |
|---|---|
| Angiotensin converting enzyme inhibitors | Lisinopril |
| Anticoagulants | Heparin, Warfarin |
| Antithrombotic Agents | Streptokinase |
| Antiarrthymics | Lidocaine, Amiodarone |
| Arteriolar dilator | Hydralazine |
| Beta adrenergic receptor blockers | Bisoprolol, Carvedilol |
| Beta Lactam Antibacterials † | Benzathine benzylpenicillin, Phenoxymethylpenicillin |
| Cardiac glycosides | Digoxin |
| Cardioselective calcium channel blockers | Verapamil |
| Centrally acting sympatholytics | Methyldopa † |
| High-efficacy diuretics | Furosemide |
| Insulins and oral antidiabetic medications | Insulin injection, Intermediate-acting insulin, Glicazide, Glucagon Metformin |
| Lipid modifying agents | Simvastatin |
| Low efficacy diuretics | Hydrochlorothiazide |
| Organic nitrates | Glyceryl Trinitrate, Isosorbide Dinitrate |
| Other analgesics and antipyretics | Acetylsalicylic acid |
| Positive inotropes except digoxin | Dopamine, Adrenaline |
| Potassium sparing diuretics | Spironolactone |
| Vasoselective calcium channel blockers | Amlodipine |
| Clinical Studies | ||||
|---|---|---|---|---|
| CVD | No. of Patients | Change in Gut Microbiota Composition/Metabolites | Outcome | Reference |
| Atherosclerosis | 332 | Increased LBP | Increased carotid intima media thickness | [46] |
| 4144 | Increased TMAO | Increased atherosclerotic risk | [47] | |
| CAD | 2255 | Increased risk of artery infarction | [48] | |
| 59 | Increased L-carnitine | Increased TMAO in CAD patients | [49] | |
| 126 | Increased LPS | Increased inflammatory cytokines | [50] | |
| 30 | Reduced Bacteroides vulgatus and B. dorei and LPS | Increased lesions | [51] | |
| CAD and artery stenosis | 169 | Increased TMAO | Increased risk of CAD and artery stenosis | [17] |
| Heart failure | 122 | Increased LPS | LPS translocation through leaky gut, resulting in inflammation | [52] |
| 452 | Endotoxemia inflammation and oxidative stress | [52] | ||
| Heart attack | 38 | Increased proteobacteria LPS and leaky gut | Increased endotoxemia | [53] |
| Atrial fibrillation | 912 | Increased LPS | Increased platelet activation | [54] |
| Animal studies | ||||
| CVD | Animal | Change in gut microbiota composition/metabolites | Outcome | Reference |
| Atherosclerosis | Mice | Increased LPS | Activation of NF-κB and JNK pathways | [55] |
| Mice | Increased size of atherosclerotic lesions | [56] | ||
| Mice | Increased proinflammatory cytokines | [57] | ||
| Mice | Increased TMAO | Nlrp3 inflammasome stimulation and endothelial dysfunction | [58] | |
| Mice | Increased plague area | [48] | ||
| Increased expression of inflammatory genes | [48] | |||
| Mice | Butyrate supplementation | Reduced cholesterol absorption and atherosclerotic lesion | [59] | |
| Mice | Reduced SCFAs and Akkermansia, Clostridium, and Odoribacter | Increased plague size | [60] | |
| Mice | Reduced Bacteroidetes and Clostridia | Increased dyslipidaemia | [61] | |
| Heart failure | Mice | Increased TMAO | Increased severity of heart failure | [48] |
| Hypertension | Rat | Increased osmotic pressure and water reabsorption | [62] | |
| Cardiomyopathy | Mice | Increased LPS | Increased inflammatory markers | [63] |
| Drug | Bacteria | Mechanism(s) | Outcome | Reference |
|---|---|---|---|---|
| Known microbiome–drug interactions | ||||
| Digoxin | Eggerthella lenta | Inactivation by reduction | Bacterial reductase activity decreases the amount of the active drug reaching target tissues | [98] |
| Proposed microbiome–drug interactions | ||||
| Simvastatin | Not known | Microbial-derived bile acids competing for host uptake transporters Alteration in bacterial communities with bile salt hydrolase (bsh) activity | Decreased amount of the drug reaching target tissues Variability in FXR receptor signaling | [98,99] |
| Rosuvastatin | Not known | Alteration in host gene expression in bile acid metabolism pathways Alteration in bacterial communities with bile salt hydrolase (bsh) activity | Variability in FXR receptor signaling | [98] |
| Atorvastatin | Not known | Decreased amount of secondary bile acids | Variability in FXR receptor signaling | [98] |
| Lovastatin | Not known | Increase metabolism (hydrolysis) | Altering its lipid lowering activity | [100] |
| Amlodipine | Not known | Pre-systemic metabolism by dehydrogenation | Decreased amount of the active drug reaching target tissues | [98] |
| Nifedipine | Not known | Decreased absorption | Decreased potency of the drug | [101] |
| Captopril | Not known | Not known | Decreased intestinal permeability and improved villi length | [98] |
| Enalapril | Not known | De-esterification | Increased biotransformation | [101] |
| Aspirin | Not known | Hypoxic conditions alter the metabolic activity of the intestinal flora | Increase absorption of aspirin in rats and increased risk of bleeding | [101] |
| Amiodarone | Escherichia coli Nissle 1917 | Not known | Increased activity | [100] |
| Glyceryl trinitrate | Denitration | Decreased activity | [100] | |
| Quercetin-3-glucoside | Eubacterium ramulus and Enterococcus casselilfavus | Deglycosylation | Decreased activity | [100] |
| FXR—farnesoid X receptor | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhajri, N.; Khursheed, R.; Ali, M.T.; Abu Izneid, T.; Al-Kabbani, O.; Al-Haidar, M.B.; Al-Hemeiri, F.; Alhashmi, M.; Pottoo, F.H. Cardiovascular Health and The Intestinal Microbial Ecosystem: The Impact of Cardiovascular Therapies on The Gut Microbiota. Microorganisms 2021, 9, 2013. https://doi.org/10.3390/microorganisms9102013
Alhajri N, Khursheed R, Ali MT, Abu Izneid T, Al-Kabbani O, Al-Haidar MB, Al-Hemeiri F, Alhashmi M, Pottoo FH. Cardiovascular Health and The Intestinal Microbial Ecosystem: The Impact of Cardiovascular Therapies on The Gut Microbiota. Microorganisms. 2021; 9(10):2013. https://doi.org/10.3390/microorganisms9102013
Chicago/Turabian StyleAlhajri, Noora, Rubiya Khursheed, Mohammad Taher Ali, Tareq Abu Izneid, Oumaima Al-Kabbani, Mahdia B. Al-Haidar, Fatima Al-Hemeiri, Mohamed Alhashmi, and Faheem Hyder Pottoo. 2021. "Cardiovascular Health and The Intestinal Microbial Ecosystem: The Impact of Cardiovascular Therapies on The Gut Microbiota" Microorganisms 9, no. 10: 2013. https://doi.org/10.3390/microorganisms9102013
APA StyleAlhajri, N., Khursheed, R., Ali, M. T., Abu Izneid, T., Al-Kabbani, O., Al-Haidar, M. B., Al-Hemeiri, F., Alhashmi, M., & Pottoo, F. H. (2021). Cardiovascular Health and The Intestinal Microbial Ecosystem: The Impact of Cardiovascular Therapies on The Gut Microbiota. Microorganisms, 9(10), 2013. https://doi.org/10.3390/microorganisms9102013