Analysis of Cytoplasmic and Secreted Proteins of Staphylococcus aureus Revealed Adaptive Metabolic Homeostasis in Response to Changes in the Environmental Conditions Representative of the Human Wound Site

 , and
, and
Abstract
1. Introduction
2. Experimental Procedures
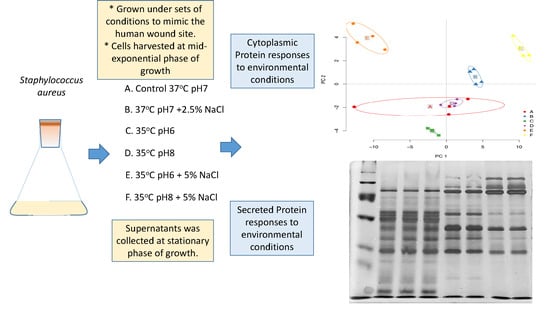
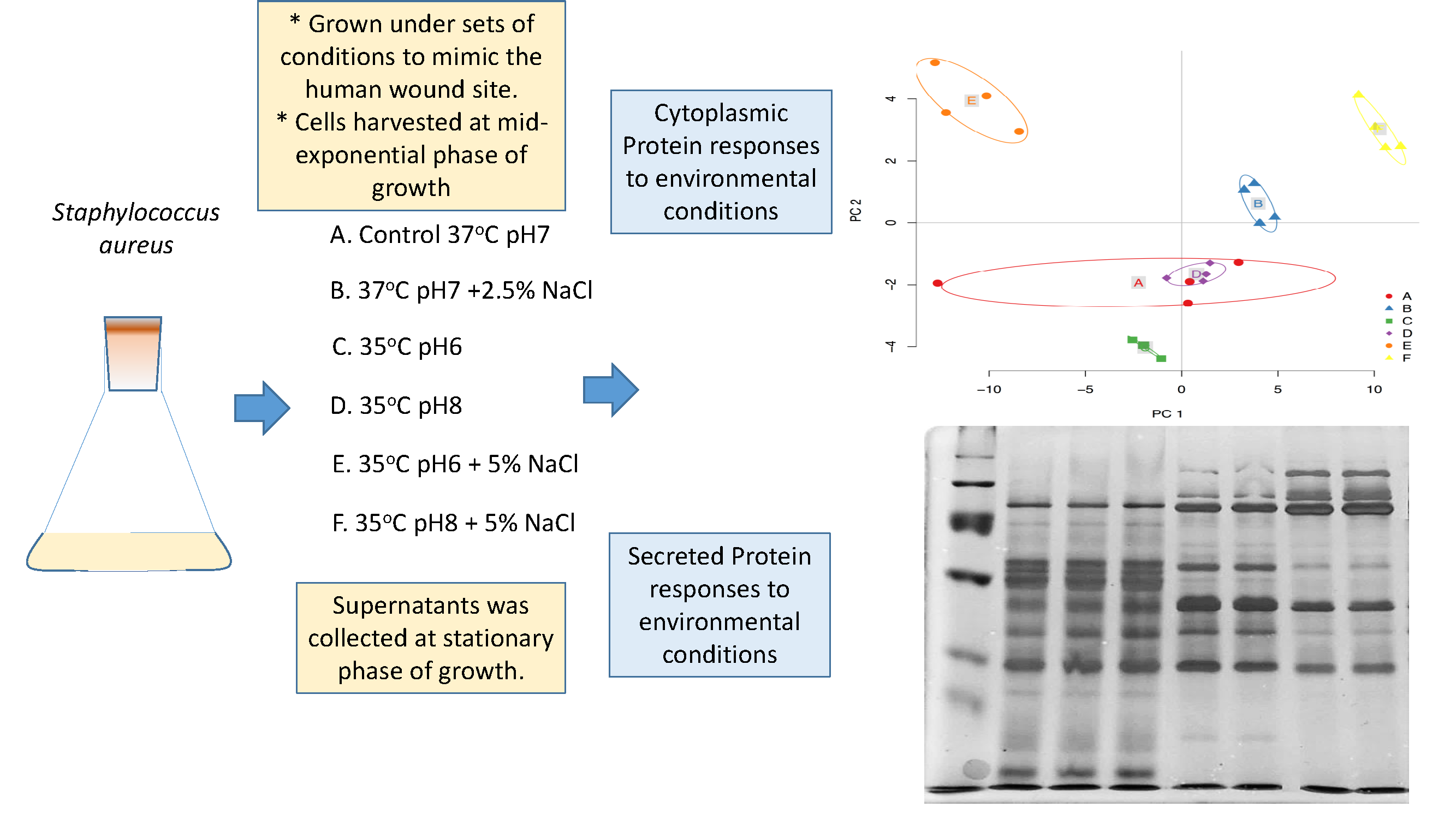
2.1. Bacterial Strain and Growth Conditions
2.2. Experimental Design and Statistical Rational
2.3. Extraction and Estimation of Cytoplasmic Proteins
2.4. Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Staining Procedures
2.5. Protein Reduction, Alkylation and Digestion
2.6. LC–MS/MS Analysis of Cytoplasmic Proteins
2.7. Database Search and Protein Quantification
2.8. Analysis of Secreted Proteins
3. Results
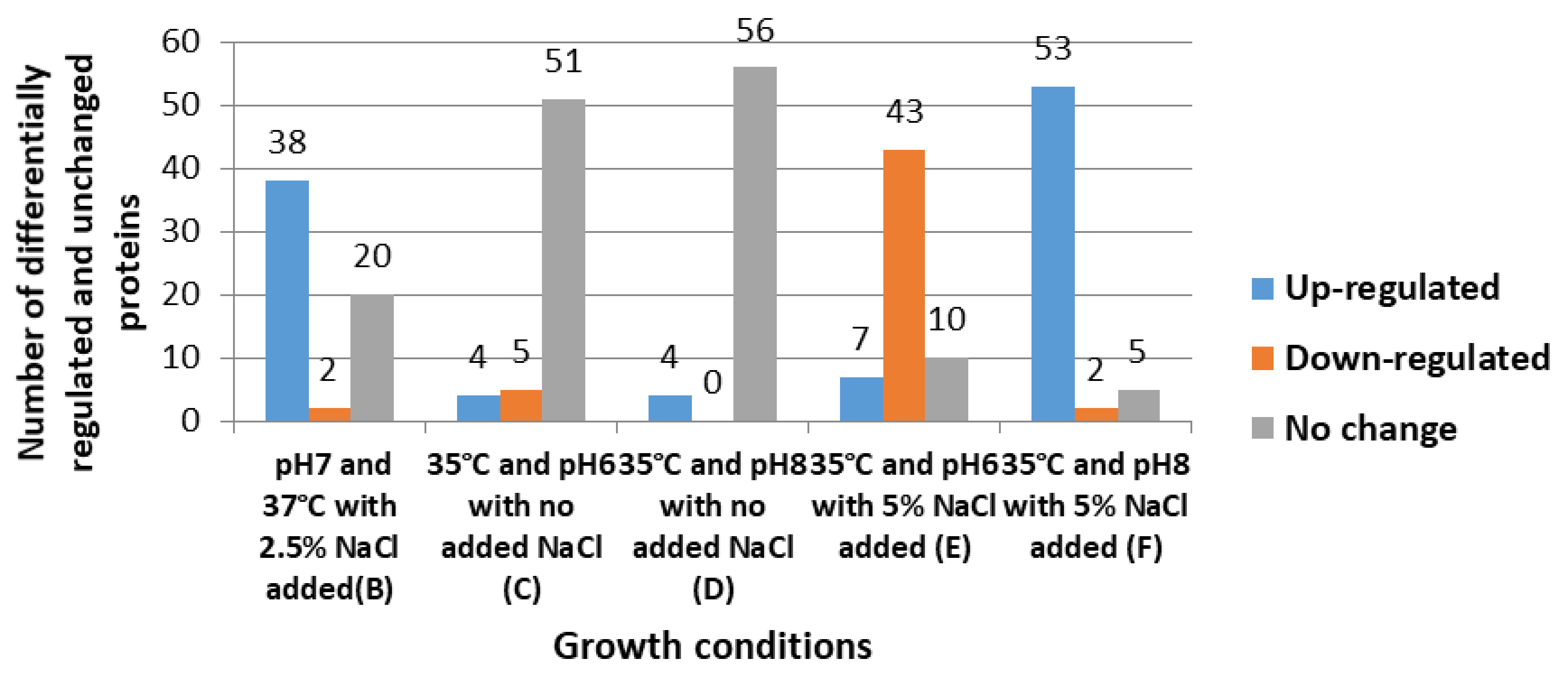
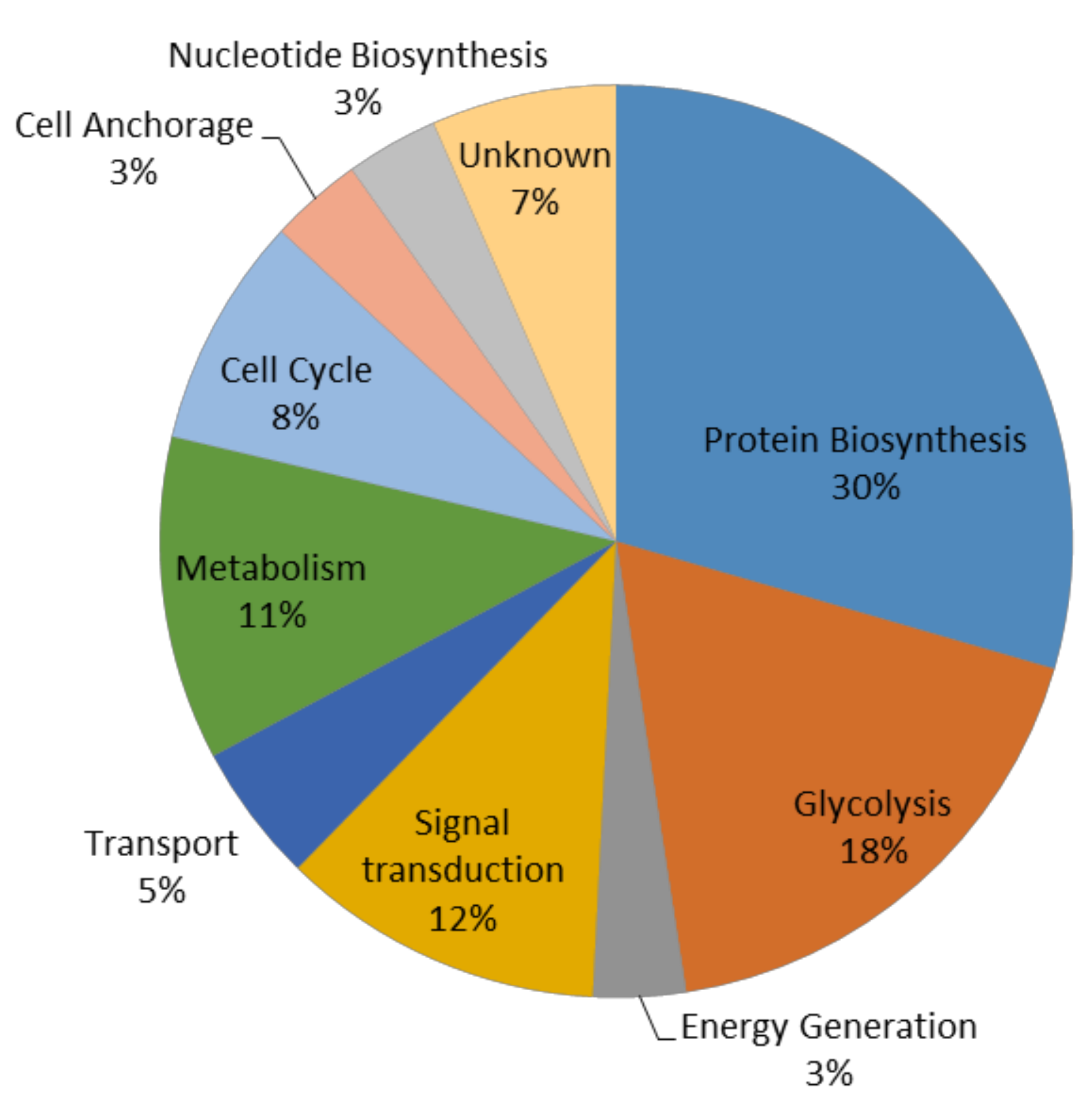
3.1. Cytoplasmic Proteomic Analysis
3.2. Clustering Analysis
3.3. Secreted Protein Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foster, T.J. Colonization and infection of the human host by staphylococci: Adhesion, survival and immune evasion. Vet. Dermatol. 2009, 20, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Coates, R.; Moran, J.; Horsburgh, M.J. Staphylococci: Colonizers and pathogens of human skin. Future Microbiol. 2014, 9, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; McCann, M.; Kavanagh, K. Proteomic analysis of the proteins released from Staphylococcus aureus following exposure to Ag(I). Toxicol. Vitro 2013, 27, 1644–1648. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Onyango, L.A.; Alreshidi, M.M. Adaptive Metabolism in Staphylococci: Survival and Persistence in Environmental and Clinical Settings. J. Pathog. 2018, 2018, 1092632. [Google Scholar] [CrossRef]
- Alreshidi, M.M.; Dunstan, R.H.; Onyango, L.A.; Roberts, T.K. Staphylococcal phenomics: Metabolomic and proteomic responses to environmental stessors. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013; pp. 690–701. [Google Scholar]
- Alreshidi, M.M.; Dunstan, R.H.; Gottfries, J.; Macdonald, M.M.; Crompton, M.J.; Ang, C.S.; Williamson, N.A.; Roberts, T.K. Changes in the Cytoplasmic Composition of Amino Acids and Proteins Observed in Staphylococcus aureus during Growth under Variable Growth Conditions Representative of the Human Wound Site. PLoS ONE 2016, 11, e0159662. [Google Scholar] [CrossRef]
- Alreshidi, M.M.; Dunstan, R.H.; Macdonald, M.M.; Smith, N.D.; Gottfries, J.; Roberts, T.K. Metabolomic and proteomic responses of Staphylococcus aureus to prolonged cold stress. J. Proteomics. 2015, 121, 44–55. [Google Scholar] [CrossRef]
- Onyango, L.A.; Dunstan, R.H.; Gottfries, J.; Eiff, C.V.; Roberts, T.K. Effect of Low Temperature on Growth and Ultra-Structure of Staphylococcus spp. PLoS ONE 2012, 7, e29031. [Google Scholar] [CrossRef]
- Onyango, L.A.; Dunstan, R.H.; Roberts, T.K.; Macdonald, M.M.; Gottfries, J. Phenotypic variants of staphylococci and their underlying population distributions following exposure to stress. PLoS ONE 2013, 8, e77614. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Hussain, M.; Völker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Rorh, J.; Proctor, R.A.; Becker, K.; Peters, G.; et al. Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 2011, 3, 129–141. [Google Scholar] [CrossRef]
- Proctor, R.A.; Kriegeskorte, A.; Kahl, B.C.; Becker, K.; Löffler, B.; Peters, G. Staphylococcus aureus Small Colony Variants (SCVs): A road map for the metabolic pathways involved in persistent infections. Front. Cell Infect. Microbiol. 2014, 4, 99. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Loffler, B.; Proctor, R.A. Persistence of Staphylococcus aureus: Multiple Metabolic Pathways Impact the Expression of Virulence Factors in Small-Colony Variants (SCVs). Front. Microbiol. 2020, 11, 1028. [Google Scholar] [CrossRef] [PubMed]
- Von Eiff, C. Staphylococcus aureus small colony variants: A challenge to microbiologists and clinicians. Int. J. Antimicrob. Agents 2008, 31, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Sotto, A.; Lina, G.; Richard, J.L.; Combescure, C.; Bourg, G.; Vidal, L.; Jourdan, N.; Etienne, J.; Lavihne, J.P. Virulence potential of Staphylococcus aureus strains isolated from diabetic foot ulcers: A new paradigm. Diabetes Care 2008, 31, 2318–2324. [Google Scholar] [CrossRef] [PubMed]
- Almeida, G.C.; Santos, M.M.; Lima, N.G.; Cidral, T.A.; Melo, M.C.; Lima, K.C. Prevalence and factors associated with wound colonization by Staphylococcus spp. and Staphylococcus aureus in hospitalized patients in inland northeastern Brazil: A cross-sectional study. BMC Infect. Dis. 2014, 14, 328. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.A.; Korber, A.; Grabbe, S.; Dissemond, J. Influence of pH on wound-healing: A new perspective for wound-therapy? Arch. Dermatol. Res. 2007, 298, 413–420. [Google Scholar] [CrossRef]
- Lenhardt, R.; Sessler, D.I. Estimation of mean body temperature from mean skin and core temperature. Anesthesiology 2006, 105, 1117–1121. [Google Scholar] [CrossRef]
- Celeste, C.J.; Deschesne, K.; Riley, C.B.; Theoret, C.L. Skin Temperature during Cutaneous Wound Healing in an Equine Model of Cutaneous Fibroproliferative Disorder: Kinetics and Anatomic-Site Differences. Vet. Surg. 2013, 42, 147–153. [Google Scholar] [CrossRef]
- Crompton, M.J.; Dunstan, R.H.; Macdonald, M.M.; Gottfries, J.; von Eiff, C.; Roberts, T.K. Small changes in environmental parameters lead to alterations in antibiotic resistance, cell morphology and membrane fatty acid composition in Staphylococcus lugdunensis. PLoS ONE 2014, 9, e92296. [Google Scholar] [CrossRef][Green Version]
- Alreshidi, M.M.; Dunstan, R.H.; Macdonald, M.M.; Gottfries, J.; Roberts, T.K. The Uptake and Release of Amino Acids by Staphylococcus aureus at Mid-Exponential and Stationary Phases and Their Corresponding Responses to Changes in Temperature, pH and Osmolality. Front. Microbiol. 2020, 10, 3059. [Google Scholar] [CrossRef]
- Wehrli, P.M.; Lindberg, E.; Svensson, O.; Sparén, A.; Josefson, M.; Dunstan, R.H.; Wold, A.E.; Gottfries, J. Exploring bacterial phenotypic diversity using factorial design and FTIR multivariate fingerprinting. Chemometrics 2014, 30, 283–289. [Google Scholar] [CrossRef]
- Butt, H.L.; Dunstan, R.H.; McGregor, N.R.; Roberts, T.K.; Zerbes, M.; Klineberg, I.J. An association of membrane-damaging toxins from coagulase-negative staphylococci and chronic orofacial muscle pain. J. Med. Microbiol. 1998, 47, 577–584. [Google Scholar] [CrossRef]
- Alreshidi, M.M. Selected Metabolites Profiling of Staphylococcus aureus Following Exposure to Low Temperature and Elevated Sodium Chloride. Front. Microbiol. 2020, 11, 834. [Google Scholar] [CrossRef] [PubMed]
- Missiakas, D.M.; Schneewind, O. Growth and laboratory maintenance of Staphylococcus aureus. Curr. Protoc. Microbiol. 2013. Chapter 9, Unit 9C 1. [Google Scholar] [CrossRef]
- Brown, G.K.; Martin, A.R.; Roberts, T.K.; Aitken, R.J. Detection of Ehrlichia platys in dogs in Australia. Aust. Vet. J. 2001, 79, 554–558. [Google Scholar] [CrossRef]
- Scharfman, A.; Kroczynski, H.; Carnoy, C.; Brussel, E.V.; Lamblin, G.; Ramphal, R.; Roussel, P. Adhesion of Pseudomonas aeruginosa to respiratory mucins and expression of mucin-binding proteins are increased by limiting iron during growth. Infect. Immun. 1996, 64, 5417–5420. [Google Scholar] [CrossRef] [PubMed]
- Weschler, L.B. Sweat electrolyte concentrations obtained from within occlusive coverings are falsely high because sweat itself leaches skin electrolytes. J. Appl. Physiol. 2008, 105, 1376–1377. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Siqueira, W.L.; Bakkal, M.; Xiao, Y.; Sutton, J.N.; Mendes, F.M. Quantitative proteomic analysis of the effect of fluoride on the acquired enamel pellicle. PLoS ONE 2012, 7, e42204. [Google Scholar] [CrossRef]
- Kriegeskorte, A.; König, S.; Sander, G.; Pirkl, A.; Mahabir, E.; Proctor, R.A.; von Eiff, C.; Peters, G.; Becker, K. Small colony variants of Staphylococcus aureus reveal distinct protein profiles. Proteomics 2011, 11, 2476–2490. [Google Scholar] [CrossRef] [PubMed]
- Ming, T.; Geng, L.; Feng, Y.; Lu, C.; Zhou, J.; Li, Y.; Zhang, D.; He, S.; Li, Y.; Cheong, L.; et al. iTRAQ-Based Quantitative Proteomic Profiling of Stapfhylococcus aureus Under Different Osmotic Stress Conditions. Front. Microbiol. 2019, 10, 1082. [Google Scholar] [CrossRef] [PubMed]
- Tobin, P.J.; Mani, N.; Jayaswal, R.K. Effect of physiological conditions on the autolysis of Staphylococcus aureus strains. Antonie Van Leeuwenhoek 1994, 65, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Sugai, M.; Komatsuzawa, H.; Akiyama, T.; Hong, Y.M.; Oshida, T.; Yamaguchi, T.; Suginaka, H. Identification of endo-beta-N-acetylglucosaminidase and N-acetylmuramyl-L-alanine amidase as cluster-dispersing enzymes in Staphylococcus aureus. J. Bacteriol. 1995, 177, 1491–1496. [Google Scholar] [CrossRef]
- Singh, V.K. High Level Expression and Purification of Atl, the Major Autolytic Protein of Staphylococcus aureus. Int. J. Microbiol. 2014, 2014, 615965. [Google Scholar] [CrossRef]
- Sudagidan, M.; Cavusoglu, C.; Bacakoglu, F. Investigation of the virulence genes in methicillin-resistant Staphylococcus aureus strains isolated from biomaterial surfaces. Mikrobiyol. Bul. 2008, 42, 29–39. [Google Scholar]
- Merghni, A.; Nejma, M.B.; Hentati, H.; Mahjoub, A.; Mastouri, M. Adhesive properties and extracellular enzymatic activity of Staphylococcus aureus strains isolated from oral cavity. Microb. Pathog. 2014, 73, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Xiong, N.; Zhang, Y.; Rayner, S.; Chen, S. Functional characterization of lipase in the pathogenesis of Staphylococcus aureus. Biochem. Biophys. Res. Commun. 2012, 419, 617–620. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Gotz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef]
- Monzon, M.; Oteiza, C.; Leiva, J.; Amorena, B. Synergy of different antibiotic combinations in biofilms of Staphylococcus epidermidis. J. Antimicrob. Chemother. 2001, 48, 793–801. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins--an update. Mol. Biosyst. 2009, 5, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Squires, C.L.; Zaporojets, D. Proteins shared by the transcription and translation machines. Annu. Rev. Microbiol. 2000, 54, 775–798. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Process | Molecular Function | Uniprot Entry Name | Proteins ID | Experimental Conditions | ||||
|---|---|---|---|---|---|---|---|---|
| B | C | D | E | F | ||||
| 37 °C 2.5%NaCl pH7; Centroid | 35 °C 0%NaCl pH6; | 35 C 0%NaCl pH8; | 35 °C 5%NaCl pH6; | 35 °C 5%NaCl pH8; | ||||
| Protein biosynthesis | Elongation factor | EFG_STAA1 | Elongation factor G | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Elongation factor | EFTS_STAA1 | Elongation factor Ts | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Elongation factor | EFTU_STAA1 | Elongation factor Tu | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Elongation factor | EFP_STAA1 | Elongation factor P | NC | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Aminoacyl–tRNA synthetase and ligase | SYV_STAA1 | Valine__tRNA ligase | Up regulated | NC | NC | NC | Up regulated |
| Protein biosynthesis | Initiation factor | IF2_STAA1 | IF2_STAA1 translation initiation factor IF–2 | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Ligase | GATA_STAA2 | Glutamyl_tRNA (Gln) amidotransferase subunit A | Up regulated | NC | Up regulated | Down regulated | Up regulated |
| Protein biosynthesis | Ligase | GATB_STASP | Aspartyl/glutamyl_tRNA (Asn/Gln) amidotransferase subunit B | Up regulated | NC | NC | NC | Up regulated |
| Protein biosynthesis | Aminoacyl–tRNA synthetase and ligase | SYI1_STAAU | Isoleucine__tRNA ligase | Up regulated | NC | NC | NC | Up regulated d |
| Protein biosynthesis | Aminoacyl–tRNA synthetase, ligase | SYP_STAA1 | Proline__tRNA ligase | NC | NC | NC | Up regulated | Up regulated |
| Protein biosynthesis | Glycine–tRNA ligase | SYG_STAA1 | Glycine__tRNA ligase | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein biosynthesis | Aminoacyl–tRNA synthetase and ligase | SYS_STAA1 | Serine__tRNA ligase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycolysis | Oxidoreductase | G3P1_STAAC | Glyceraldehyde_3_phosphate dehydrogenase 1 | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycolysis | Kinase and transferase | KPYK_STAA3 | Pyruvate kinase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycolysis | Oxidoreductase | DLDH_STAAC | Dihydrolipoyl dehydrogenase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycolysis | Oxidoreductase | ODPB_STAAM | Pyruvate dehydrogenase E1 component subunit beta | NC | NC | NC | Down regulated | NC |
| Glycolysis | Lyase | ALF2_STAAC | Fructose_bisphosphate aldolase | NC | NC | NC | Down regulated | Up regulated |
| Glycolysis | Allosteric enzyme, kinase and transferase | K6PF_STAA2 | Dependent 6_phosphofructokinase | Up regulated | NC | NC | Down regulated | Up regulated |
| Gluconeogenesis and glycolysis | Isomerase | G6PI_STAA1 | Glucose_6_phosphate isomerase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycolysis and virulence | Lyase | ENO_STAA1 | Enolase | NC | NC | NC | Down regulated | Up regulated |
| Virulence | Chaperon | CLPL_STAA3 | ATP_dependent Clp protease ATP_binding subunit ClpL | NC | NC | NC | Down regulated | Up regulated |
| Transcription regulation and virulence | Activator, DNA-binding and repressor | MGRA_STAA8 | HTH_type transcriptional regulator MgrA | Up regulated | NC | NC | NC | Up regulated |
| Transcription | Nucleotidyltransferase and transferase | RPOB_STAA9 | DNA directed RNA polymerase subunit beta | Up regulated | NC | NC | NC | Up regulated |
| Stress response | Oxidoreductase | LDH1_STAA1 | L_lactate dehydrogenase 1 | Up regulated | NC | NC | Down regulated | Up regulated |
| Stress response | Chaperone | DNAK_STAA1 | Chaperone protein DnaK | NC | Down regulated | NC | Down regulated | Up regulated |
| General stress protein | Alkaline pH tolerance | ASP23_STAA3 | Alkaline shock protein 23 | NC | NC | NC | Up regulated | NC |
| DNA damage repair and stress response | Hydrolase | HCHA_STAAW | Protein/nucleic acid deglycase HchA | Up regulated | Down regulated | NC | Up regulated | Up regulated |
| ATP synthesis, hydrogen ion transport, ion transport, transport | Translocase | ATPA_STAA1 | ATP synthase subunit alpha | NC | Down regulated | NC | Down regulated | Up regulated |
| Ion storage | Oxidoreductase | FTN_STAA3 | Ferritin | Up regulated | NC | NC | NC | Up regulated |
| Alcohol metabolic process | Oxidoreductase | ADH_STAA3 | Alcohol dehydrogenase | Up regulated | NC | NC | Down regulated | Up regulated |
| Hydrogen peroxide catabolic process a | Oxidoreductase and peroxidase | CATA_STAA3 | Catalase; | Up regulated | NC | Up regulated | NC | Up regulated |
| DNA recombination | Transferase | TKT_STAAC | Transketolase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycine betaine biosynthetic process from choline | Oxidoreductase | BETA_STAA1 | Choline dehydrogenase | NC | NC | NC | NC | Up regulated |
| mRNA catabolic process and RNA processing | Nucleotidyltransferase RNA-binding and transferase | PNP_STAA1 | Polyribonucleotide nucleotidyltransferase | Up regulated | NC | NC | Down regulated | Up regulated |
| Glycerol metabolism | Oxidoreductase | GLPD_STAA3 | Aerobic glycerol_3_phosphate dehydrogenase | Up regulated | NC | NC | Down regulated | Up regulated |
| GMP biosynthesis | Oxidoreductase | IMDH_STAA3 | Inosine_5__monophosphate dehydrogenase | Up regulated | NC | NC | Down regulated | Up regulated |
| Carbohydrate metabolism | Acyltransferase | PFLB_STAA3 | Formate acetyltransferase | Up regulated | NC | NC | Down regulated | Up regulated |
| Carbohydrate metabolic process | Magnesium ion binding and phosphoglucosamine mutase activity | GLMM_STAA1 | Phosphoglucosamine mutase | Up regulated | NC | NC | Down regulated | Up regulated |
| Acetyl-CoA biosynthetic process and organic acid metabolic process | Kinase transferase | ACKA_STAA1 | Acetate kinase | NC | NC | NC | Down regulated | NC |
| Protein transport, translocation and transport | ATPase-coupled protein transmembrane transporter activity, ATP binding and metal ion binding | SECA1_STAA1 | Protein translocase subunit SecA 1 | Up regulated | NC | NC | Down regulated | Up regulated |
| Uncharacterized | Uncharacterized | Y1663_STAAN | UPF0342 protein SA1663 | NC | Down regulated | NC | Up regulated | Up regulated |
| Uncharacterized | Uncharacterized | Y1402_STAAN | UPF0365 protein SA1402 | NC | NC | NC | Down regulated | Up regulated |
| Uncharacterized | Uncharacterized | Y1582_STAA3 | UPF0337 protein SAUSA300_1582 | Up regulated | Down regulated | NC | Up regulated | Up regulated |
| Amino acid biosynthesis | Transferase | CYSK_STAAC | CYSK_STAAC cysteine synthase | NC | NC | NC | Down regulated | NC |
| Amino acid biosynthesis | Ligase | GLNA_STAAC | GLNA_STAAC glutamine synthetase | Up regulated | NC | NC | Down regulated | Up regulated |
| Cell wall biogenesis/degradation | Hydrolase | ATL_STAAR | Bifunctional autolysin precursor | Up regulated | Up regulated | Up regulated | Up regulated | Up regulated |
| Cell wall biosynthesis | Adhesion | SDRD_STAAE | Serine_aspartate repeat_containing protein D | Down regulated | Up regulated | NC | Down regulated | Down regulated |
| Cell redox homeostasis and response to reactive oxygen species | Oxidoreductase | AHPF_STAA8 | Alkyl hydroperoxide reductase subunit F | NC | NC | NC | NC | Up regulated |
| Energy production and conversion | Oxidoreductase | Y807_STAAB | NADH dehydrogenase_like protein SAB0807 | Up regulated | NC | NC | Down regulated | Up regulated |
| Energy production and conversion | Hydrolase | PPAC_STAA1 | Probable manganese_dependent inorganic pyrophosphatase | Up regulated | NC | Up regulated | Down regulated | Up regulated |
| Uncharacterized | Uncharacterized | PLS_STAAM | Putative surface protein SAV2496/SAV2497; Flags: Precursor | Down regulated | Up regulated | NC | Down regulated | Down regulated |
| Asparaginyl–tRNA aminoacylation | Asparagine–tRNA ligase activity, ATP binding and nucleic acid binding | SYN_LISIN | Asparagine__tRNA ligase | Up regulated | NC | NC | Down regulated | Up regulated |
| Protein folding | peptidyl-prolyl cis-trans isomerase activity | PRSA_STAA1 | Foldase protein PrsA | NC | NC | NC | Down regulated | Up regulated |
| Cell cycle, cell division, and Septation | GTPase activity and GTP binding | FTSZ_STAA8 | Cell division protein FtsZ | NC | NC | NC | Down regulated | Up regulated |
| Cell cycle and cell division | Chaperone, isomerase and rotamase | TIG_STAA1 | Trigger factor | NC | NC | NC | Down regulated | Up regulated |
| Cell cycle, cell division, cell shape, cell wall biogenesis/degradation, peptidoglycan synthesis | Transferase | MURA2_STAAN | UDP_N_acetylglucosamine 1_carboxyvinyltransferase 2 | Up regulated | NC | NC | NC | Up regulated |
| Virulence | Hydrolase | SSAA2_STAA8 | Staphylococcal secretory antigen ssaA2 | NC | Up regulated | NC | Up regulated | NC |
| Removal of superoxide radicals | Oxidoreductase | TRXB_STAEQ | Thioredoxin reductase | Up regulated | NC | NC | Down regulated | Up regulated |
| Phosphotransferase system, sugar transport and transport | Kinase and transferase | PT1_STAAC | Phosphoenolpyruvate_protein phosphotransferase | Up regulated | NC | NC | Down regulated | Up regulated |
| Pyrimidine biosynthesis | Allosteric enzyme, kinase and transferase | PYRH_STAAE | Uridylate kinase | Up regulated | NC | NC | Down regulated | Up regulated |
| Band # | Accession | Protein | Scores | MW (kDa) | Peptides |
|---|---|---|---|---|---|
| 1 | LIP1_STAAC | Lipase 1 | 1675.5 | 76.6 | 28 |
| LIP2_STAAM | Lipase 2 precursor | 1592.0 | 76.5 | 25 | |
| tr|F0D7U9|F0D7U9_STAAU | Autolysin | 810.9 | 137.0 | 15 | |
| Accession | Protein | Scores | MW (kDa) | Peptides | |
| 2 | CATA_STAAB | Catalase | 710.7 | 58.3 | 28 |
| RS1_STAAC | 30S ribosomal protein S1 | 615.1 | 43.3 | 9 | |
| H0AR71_STAAU | Elongation factor Tu | 431.5 | 43.1 | 20 | |
| tr|T1Y7Q4|T1Y7Q4_STAAU | Enolase | 406.0 | 47.0 | 8 | |
| tr|T1YAV3|T1YAV3_STAAU | Formate-tetrahydrofolate ligase | 349.2 | 61.1 | 8 | |
| tr|N5DN28|N5DN28_STAAU | ATP synthase subunit beta | 309.1 | 51.4 | 6 | |
| 3 | Accession | Protein | Scores | MW (kDa) | Peptides |
| tr|S9RPT3|S9RPT3_STAAU | Alkyl hydroperoxide reductase subunit C | 518.2 | 20.9 | 7 | |
| 4 | Accession | Protein | Scores | MW (kDa) | Peptides |
| ATL_STAAM | ATL_STAAM Bifunctional autolysin precursor (includes: N-acetylmuramoyl-L-alanine amidase (EC 3.5.1.28); mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase (EC 3.2.1.96)) | 3684.0 | 136.7 | 47 | |
| 5 | Accession | Protein | Scores | MW (kDa) | Peptides |
| ATL_STAAM | Bifunctional autolysin precursor (includes: N-acetylmuramoyl-L-alanine amidase (EC 3.5.1.28); mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase (EC 3.2.1.96) | 3684.2 | 136.7 | 57 | |
| LTAS_STAAC | Glycerol phosphate lipoteichoic acid synthase | 1940.1 | 74.4 | 32 | |
| tr|F3TIS4|F3TIS4_STAAU | Triacylglycerol lipase | 336.5 | 72.2 | 6 | |
| tr|G7ZSU9|G7ZSU9_STAAU | Probable transglycosylase isaA | 607.7 | 24.1 | 7 | |
| 6 | Accession | Protein | Scores | MW (kDa) | Peptides |
| LIP2_STAAC | Lipase 2 precursor (EC 3.1.1.3) | 620.7 | 76.3 | 12 | |
| ISAA_STAAC | Immunodominant staphylococcal antigen A precursor | 441.1 | 24.2 | 5 | |
| 7 | Accession | Protein | Scores | MW (kDa) | Peptides |
| ATL_STAAW | Bifunctional autolysin precursor (includes: N-acetylmuramoyl-L-alanine amidase (EC 3.5.1.28); mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase) | 5871.0 | 137.3 | 88 | |
| SDRD_STAAE | Serine-aspartate repeat-containing protein D | 378.3 | 142.7 | 6 | |
| 8 | Accession | Protein | Scores | MW (kDa) | Peptides |
| ATL_STAAW | Bifunctional autolysin precursor (includes: N-acetylmuramoyl-L-alanine amidase (EC 3.5.1.28); mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase (EC 3.2.1.96)) | 4932.6 | 137.3 | 77 | |
| LIP2_STAAC | Lipase 2 precursor (EC 3.1.1.3) | 404.2 | 76.3 | 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
M. Alreshidi, M.; Dunstan, R.H.; M. Macdonald, M.; K. Singh, V.; K. Roberts, T. Analysis of Cytoplasmic and Secreted Proteins of Staphylococcus aureus Revealed Adaptive Metabolic Homeostasis in Response to Changes in the Environmental Conditions Representative of the Human Wound Site. Microorganisms 2020, 8, 1082. https://doi.org/10.3390/microorganisms8071082
M. Alreshidi M, Dunstan RH, M. Macdonald M, K. Singh V, K. Roberts T. Analysis of Cytoplasmic and Secreted Proteins of Staphylococcus aureus Revealed Adaptive Metabolic Homeostasis in Response to Changes in the Environmental Conditions Representative of the Human Wound Site. Microorganisms. 2020; 8(7):1082. https://doi.org/10.3390/microorganisms8071082
Chicago/Turabian StyleM. Alreshidi, Mousa, R. Hugh Dunstan, Margaret M. Macdonald, Vineet K. Singh, and Tim K. Roberts. 2020. "Analysis of Cytoplasmic and Secreted Proteins of Staphylococcus aureus Revealed Adaptive Metabolic Homeostasis in Response to Changes in the Environmental Conditions Representative of the Human Wound Site" Microorganisms 8, no. 7: 1082. https://doi.org/10.3390/microorganisms8071082
APA StyleM. Alreshidi, M., Dunstan, R. H., M. Macdonald, M., K. Singh, V., & K. Roberts, T. (2020). Analysis of Cytoplasmic and Secreted Proteins of Staphylococcus aureus Revealed Adaptive Metabolic Homeostasis in Response to Changes in the Environmental Conditions Representative of the Human Wound Site. Microorganisms, 8(7), 1082. https://doi.org/10.3390/microorganisms8071082