Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities


Abstract
1. Introduction
2. Materials and Methods
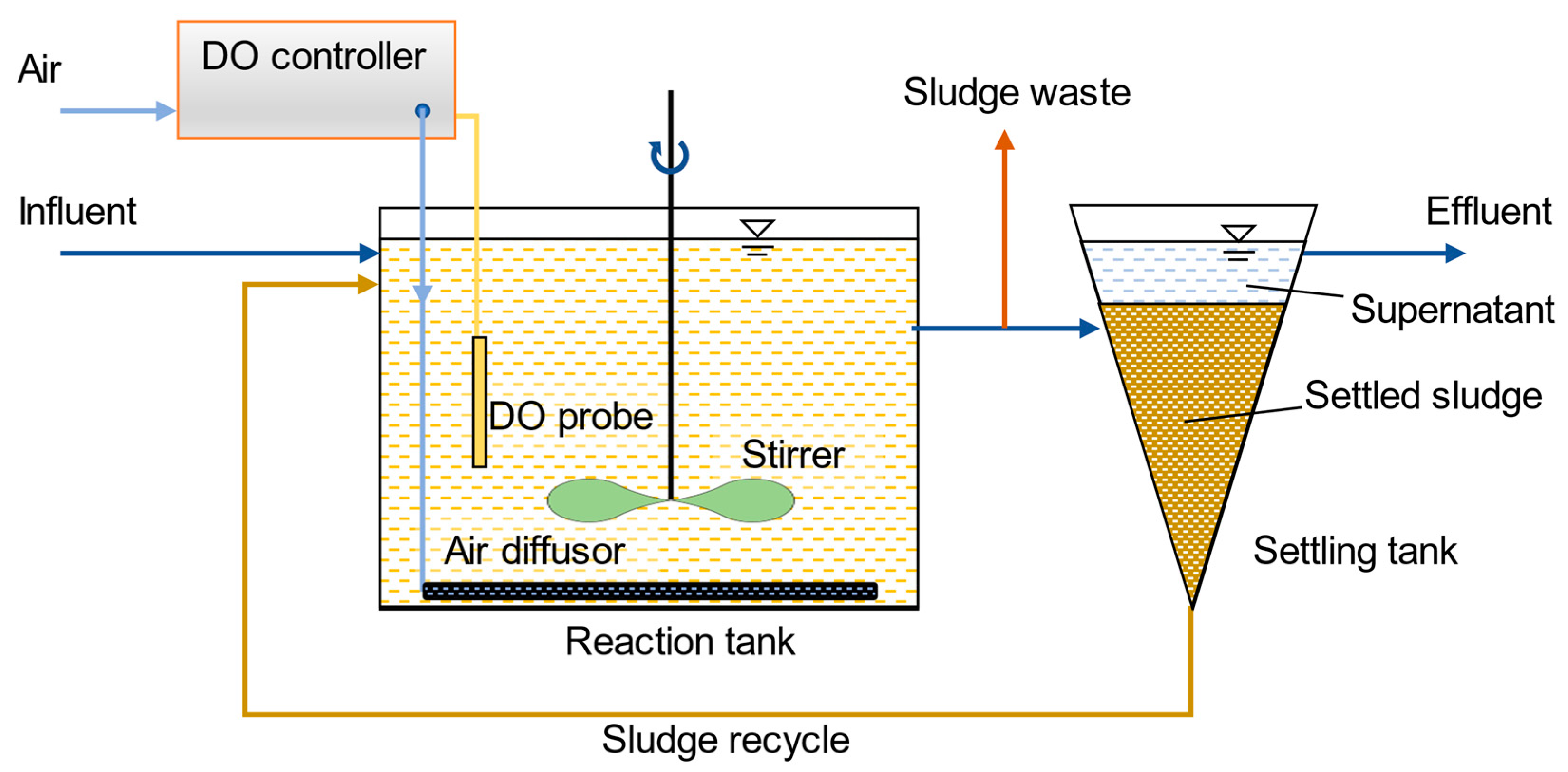
2.1. Reactor Configuration
2.2. Reactor Inoculation, Synthetic Wastewater Composition, and System Operation
2.3. Chemical Analytical Procedures
2.4. Microbial Community Analysis
2.5. Statistics Analysis
3. Results
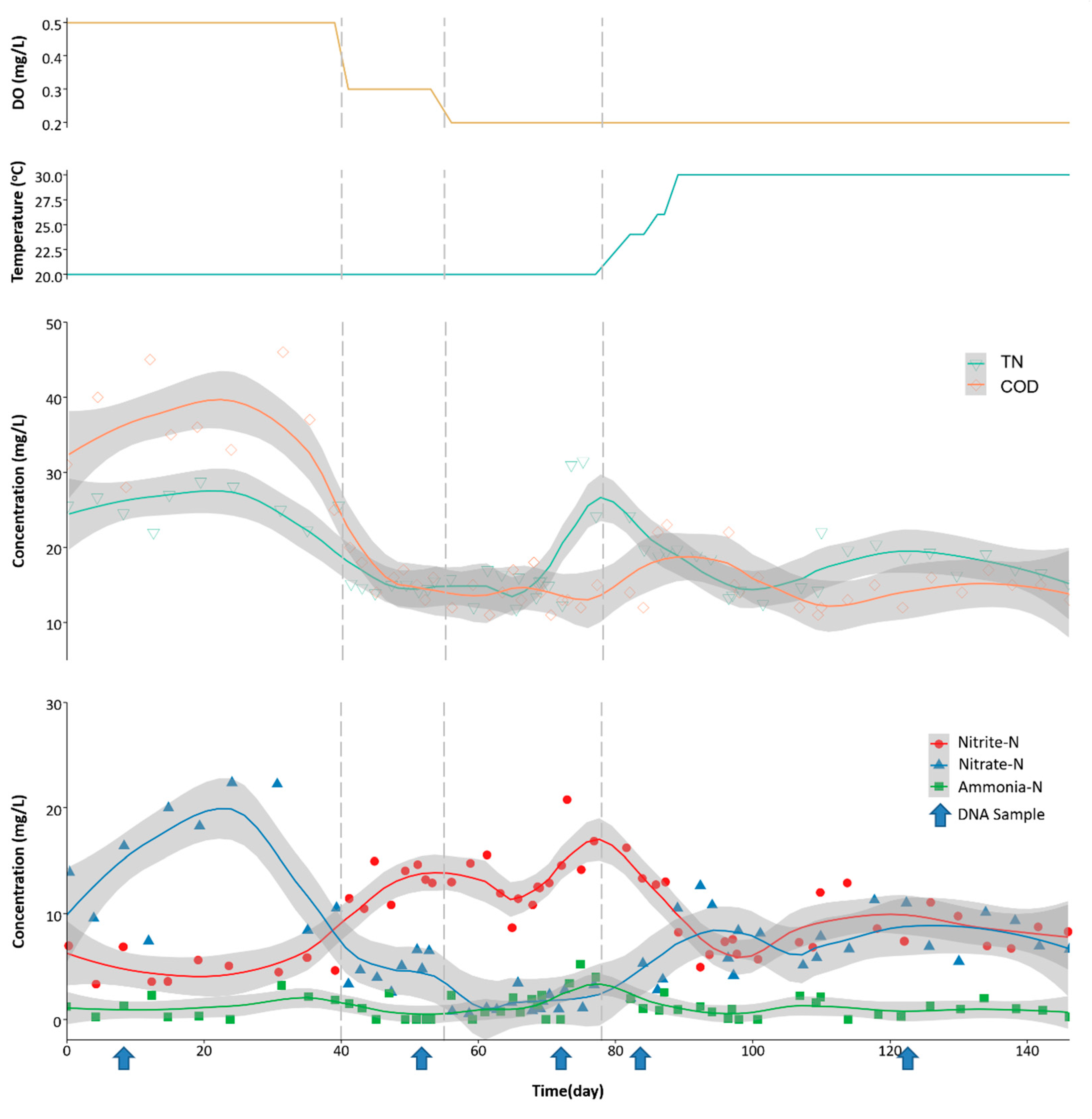
3.1. Reactor Performance
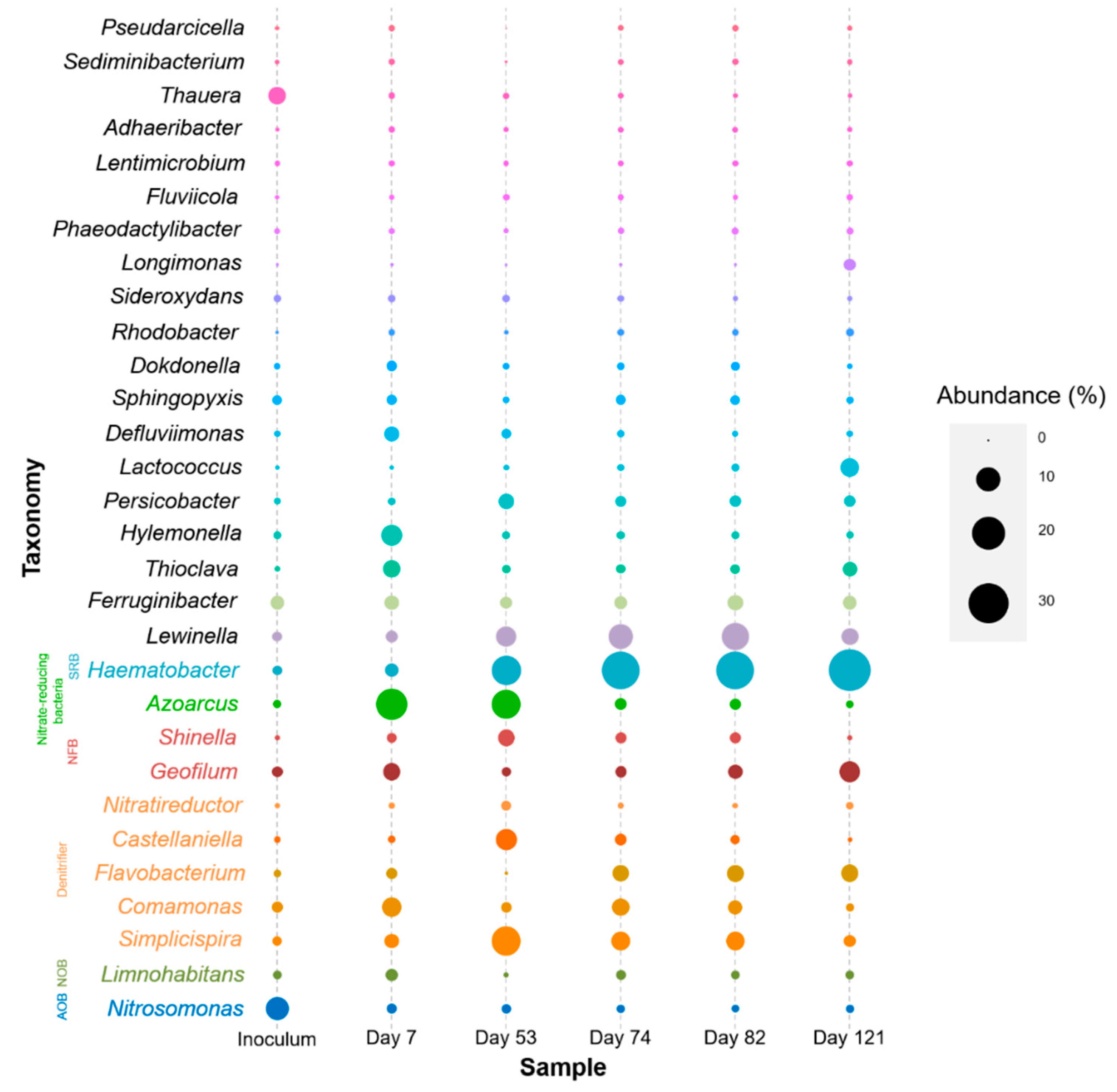
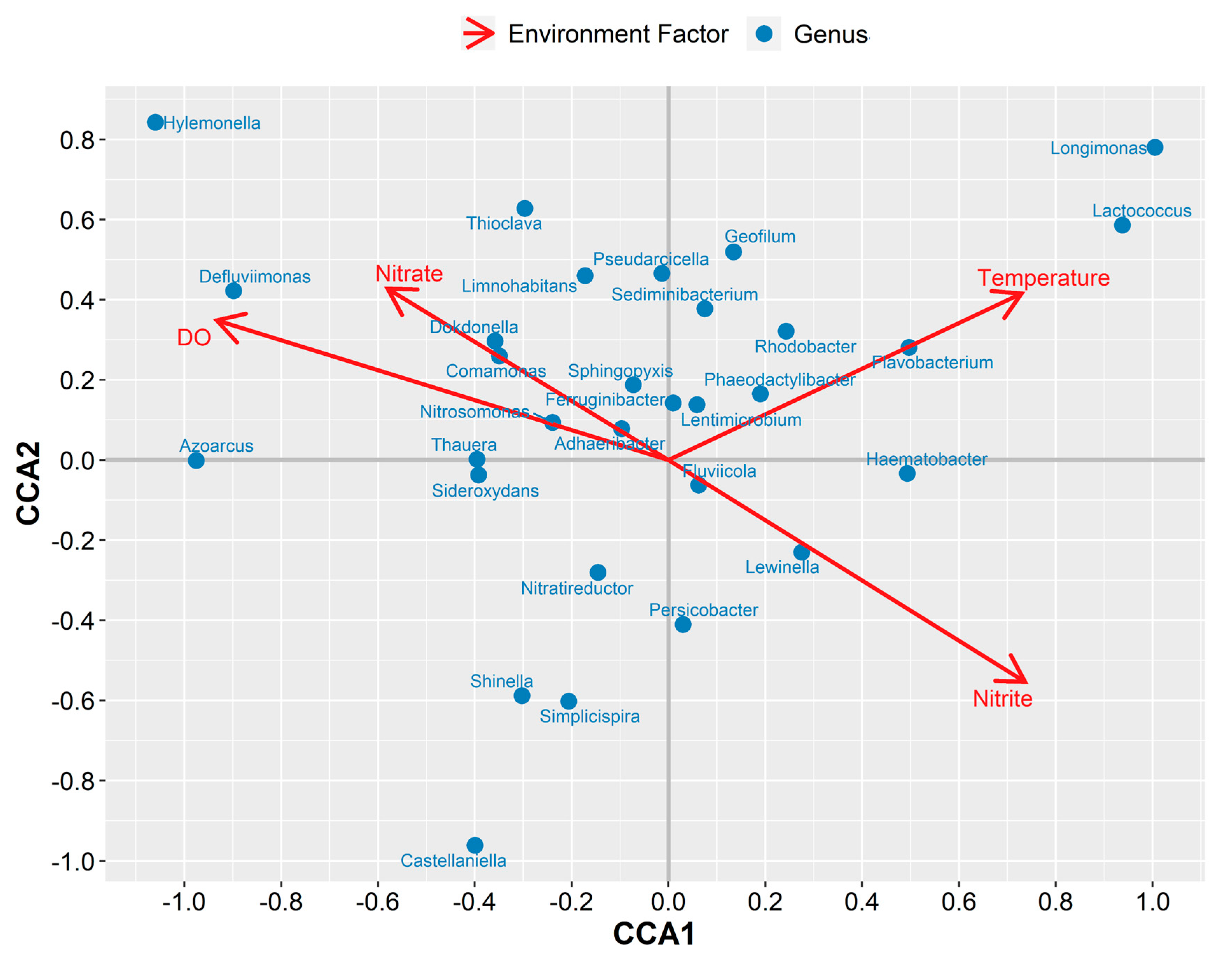
3.2. Microbial Community Changes Influenced by Operation Conditions
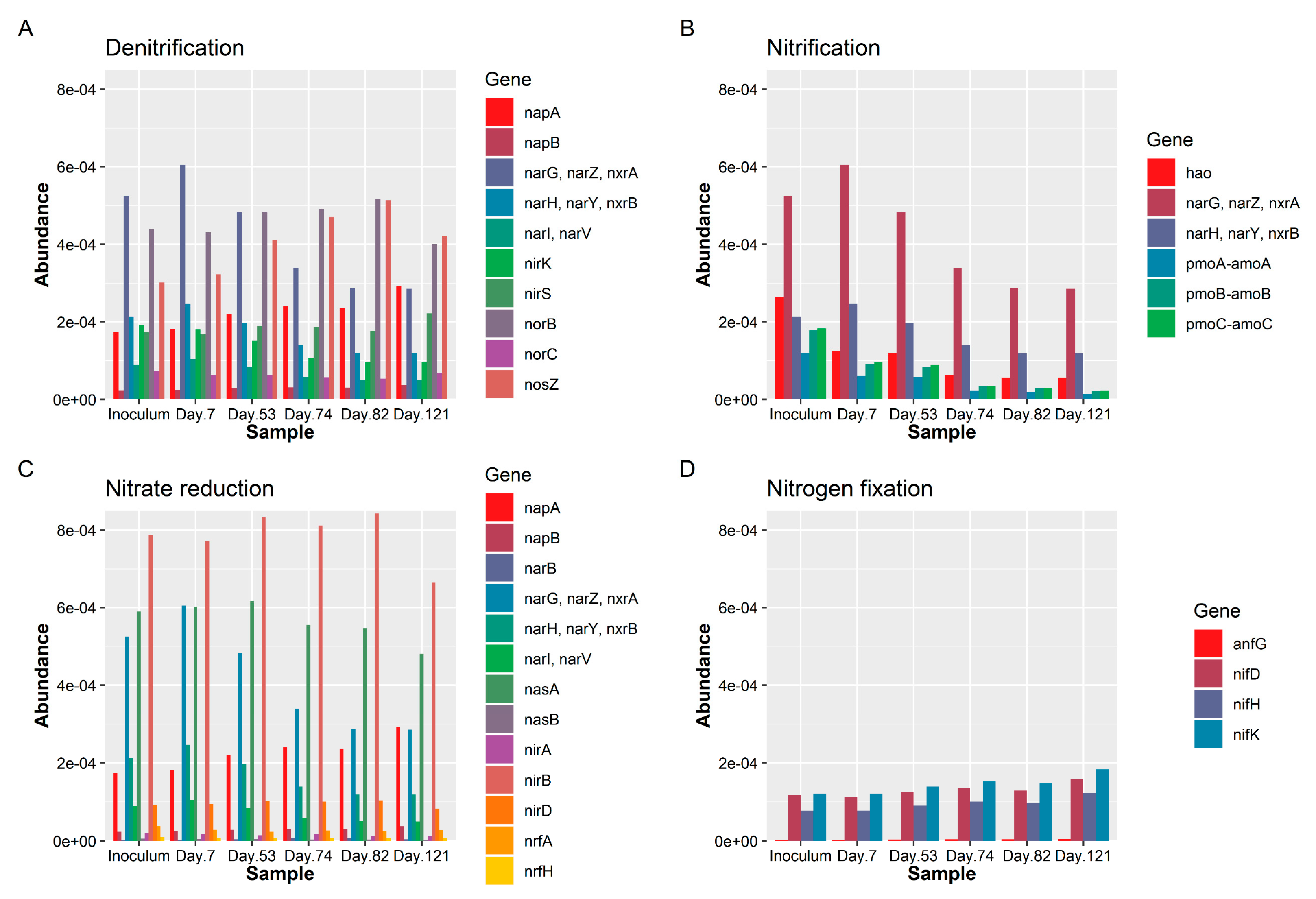
3.3. Phenotype Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, X.; Luo, J.; Guo, G.; Mackey, H.R.; Hao, T.; Chen, G. Seawater-based wastewater accelerates development of aerobic granular sludge: A laboratory proof-of-concept. Water Res. 2017, 115, 210–219. [Google Scholar] [CrossRef]
- Leung, R.; Li, D.; Yu, W.; Chui, H.K.; Lee, T.; Van Loosdrecht, M.; Chen, G. Integration of seawater and grey water reuse to maximize alternative water resource for coastal areas: The case of the Hong Kong International Airport. Water Sci. Technol. 2012, 65, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Regmi, P.; Miller, M.W.; Holgate, B.; Bunce, R.; Park, H.; Chandran, K.; Wett, B.; Murthy, S.; Bott, C.B. Control of aeration, aerobic SRT and COD input for mainstream nitritation/denitritation. Water Res. 2014, 57, 162–171. [Google Scholar] [CrossRef]
- Hellinga, C.; Schellen, A.; Mulder, J.W.; van Loosdrecht, M.v.; Heijnen, J. The SHARON process: An innovative method for nitrogen removal from ammonium-rich waste water. Water Sci. Technol. 1998, 37, 135–142. [Google Scholar] [CrossRef]
- Guo, J.; Peng, Y.; Wang, S.; Zheng, Y.; Huang, H.; Ge, S. Effective and robust partial nitrification to nitrite by real-time aeration duration control in an SBR treating domestic wastewater. Process Biochem. 2009, 44, 979–985. [Google Scholar] [CrossRef]
- Metcalf, L. Wastewater Engineering: Treatment and Reuse; Metcalf & Eddy Inc., McGraw-Hill Inc.: New York, NY, USA, 2003. [Google Scholar]
- Wang, C.-C.; Lee, P.-H.; Kumar, M.; Huang, Y.-T.; Sung, S.; Lin, J.-G. Simultaneous partial nitrification, anaerobic ammonium oxidation and denitrification (SNAD) in a full-scale landfill-leachate treatment plant. J. Hazard. Mater. 2010, 175, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-Y.; Zeng, G.-M.; Yang, Z.-H.; Xiao, Y.; Cao, M.; Sun, H.-S.; Ji, L.-L.; Chen, Y. Biological treatment of landfill leachate with the integration of partial nitrification, anaerobic ammonium oxidation and heterotrophic denitrification. Bioresour. Technol. 2010, 101, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, G.; Jeison, D.; Chamy, R. Nitrification with high nitrite accumulation for the treatment of wastewater with high ammonia concentration. Water Res. 2003, 37, 1371–1377. [Google Scholar] [CrossRef]
- Capodici, M.; Corsino, S.F.; Torregrossa, M.; Viviani, G. Shortcut nitrification-denitrification by means of autochthonous halophilic biomass in an SBR treating fish-canning wastewater. J. Environ. Manag. 2018, 208, 142–148. [Google Scholar] [CrossRef]
- She, Z.; Zhao, L.; Zhang, X.; Jin, C.; Guo, L.; Yang, S.; Zhao, Y.; Gao, M. Partial nitrification and denitrification in a sequencing batch reactor treating high-salinity wastewater. Chem. Eng. J. 2016, 288, 207–215. [Google Scholar] [CrossRef]
- Ye, L.; Peng, C.-y.; Tang, B.; Wang, S.-y.; Zhao, K.-f.; Peng, Y.-z. Determination effect of influent salinity and inhibition time on partial nitrification in a sequencing batch reactor treating saline sewage. Desalination 2009, 246, 556–566. [Google Scholar] [CrossRef]
- Pan, Y.; Ye, L.; Yuan, Z. Effect of H2S on N2O reduction and accumulation during denitrification by methanol utilizing denitrifiers. Environ. Sci. Technol. 2013, 47, 8408–8415. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, H.; Chen, G.-H.; Lau, G.N.; Tsang, W.; van Loosdrecht, M.C. A novel sulfate reduction, autotrophic denitrification, nitrification integrated (SANI) process for saline wastewater treatment. Water Res. 2009, 43, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- United States of Environmental Protection Agency. Method 410.3: Chemical Oxygen Demand (Titrimetric, High Level for Saline Waters) by Titration; Office of Water: Washington, DC, USA, 1978.
- Scheiner, D. Determination of ammonia and Kjeldahl nitrogen by indophenol method. Water Res. 1976, 10, 31–36. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, L.; Mao, Y.; Jiang, X.; Xia, Y.; Yu, K.; Li, B.; Zhang, T. Comammox in drinking water systems. Water Res. 2017, 116, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [PubMed]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Junghare, M.; Patil, Y.; Schink, B. Draft genome sequence of a nitrate-reducing, o-phthalate degrading bacterium, Azoarcus sp. strain PA01 T. Stand. Genom. Sci. 2015, 10, 90. [Google Scholar] [CrossRef]
- Helsel, L.O.; Hollis, D.; Steigerwalt, A.G.; Morey, R.E.; Jordan, J.; Aye, T.; Radosevic, J.; Jannat-Khah, D.; Thiry, D.; Lonsway, D.R. Identification of “Haematobacter”, a new genus of aerobic Gram-negative rods isolated from clinical specimens, and reclassification of Rhodobacter massiliensis as “Haematobacter massiliensis comb. nov.”. J. Clin. Microbiol. 2007, 45, 1238–1243. [Google Scholar] [CrossRef]
- Siddiqi, M.Z.; Sok, W.; Choi, G.; Kim, S.Y.; Wee, J.-H.; Im, W.T. Simplicispira hankyongi sp. nov., a novel denitrifying bacterium isolated from sludge. Antonie Van Leeuwenhoek 2019, 113, 1–8. [Google Scholar] [CrossRef]
- Liu, Q.-M.; Ten, L.N.; Im, W.-T.; Lee, S.-T. Castellaniella caeni sp. nov., a denitrifying bacterium isolated from sludge of a leachate treatment plant. Int. J. Syst. Evol. Microbiol. 2008, 58, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Labbé, N.; Parent, S.; Villemur, R. Nitratireductor aquibiodomus gen. nov., sp. nov., a novel α-proteobacterium from the marine denitrification system of the Montreal Biodome (Canada). Int. J. Syst. Evol. Microbiol. 2004, 54, 269–273. [Google Scholar]
- Lu, H.; Chandran, K.; Stensel, D. Microbial ecology of denitrification in biological wastewater treatment. Water Res. 2014, 64, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Suárez-Ojeda, M.E.; Carrera, J. Biodegradation of a high-strength wastewater containing a mixture of ammonium, aromatic compounds and salts with simultaneous nitritation in an aerobic granular reactor. Process Biochem. 2016, 51, 399–407. [Google Scholar] [CrossRef]
- KEGG. Nitrogen Metabolism-Limnohabitans sp. 103DPR2. Available online: https://www.kegg.jp/kegg-bin/show_pathway?lim00910 (accessed on 20 January 2020).
- Daims, H.; Lücker, S.; Wagner, M. A new perspective on microbes formerly known as nitrite-oxidizing bacteria. Trends Microbiol. 2016, 24, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-M.; Lee, K.; Cho, J.-C. Lewinella antarctica sp. nov., a marine bacterium isolated from Antarctic seawater. Int. J. Syst. Evol. Microbiol. 2009, 59, 65–68. [Google Scholar] [CrossRef]
- Lim, J.H.; Baek, S.-H.; Lee, S.-T. Ferruginibacter alkalilentus gen. nov., sp. nov. and Ferruginibacter lapsinanis sp. nov., novel members of the family ‘Chitinophagaceae’ in the phylum Bacteroidetes, isolated from freshwater sediment. Int. J. Syst. Evol. Microbiol. 2009, 59, 2394–2399. [Google Scholar] [CrossRef] [PubMed]
- Inoue, J.-I.; Oshima, K.; Suda, W.; Sakamoto, M.; Iino, T.; Noda, S.; Hongoh, Y.; Hattori, M.; Ohkuma, M. Distribution and evolution of nitrogen fixation genes in the phylum Bacteroidetes. Microbes Environ. 2015, ME14142. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Faria, C.; Lopes, A.R.; Svensson, L.A.; Moore, E.R.; Nunes, O.C.; Manaia, C.M. Shinella fusca sp. nov., isolated from domestic waste compost. Int. J. Syst. Evol. Microbiol. 2010, 60, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M. Microbiology: A fight for scraps of ammonia. Nature 2017, 549, 162. [Google Scholar] [CrossRef][Green Version]
- Caranto, J.D.; Lancaster, K.M. Nitric oxide is an obligate bacterial nitrification intermediate produced by hydroxylamine oxidoreductase. Proc. Natl. Acad. Sci. USA 2017, 114, 8217–8222. [Google Scholar] [CrossRef]
- Stouthamer, A.; Van’t Riet, J.; Oltmann, L. Respiration with nitrate as acceptor. In Diversity of Bacterial Respiratory Systems; CRC Press: Boca Raton, FL, USA, 2018; pp. 19–48. [Google Scholar]
- Wang, X.; Tamiev, D.; Alagurajan, J.; DiSpirito, A.A.; Phillips, G.J.; Hargrove, M.S. The role of the NADH-dependent nitrite reductase, Nir, from Escherichia coli in fermentative ammonification. Arch. Microbiol. 2019, 201, 519–530. [Google Scholar] [CrossRef]
- Yamamoto-Ikemoto, R.; Matsui, S.; Komori, T.; Bosque-Hamilton, E. Symbiosis and competition among sulfate reduction, filamentous sulfur, denitrification, and poly-P accumulation bacteria in the anaerobic-oxic activated sludge of a municipal plant. Water Sci. Technol. 1996, 34, 119–128. [Google Scholar] [CrossRef]
- Postgate, J. Nitrogen Fixation, 3rd ed.; Cambridge University Press: New York, NY, USA, 1998. [Google Scholar]
- Demtröder, L.; Pfänder, Y.; Schäkermann, S.; Bandow, J.E.; Masepohl, B. NifA is the master regulator of both nitrogenase systems in Rhodobacter capsulatus. MicrobiologyOpen 2019, 8, e921. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Tan, Z.; Hurek, T. Azoarcus. Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–19. [Google Scholar]
- Allito, B.B.; Nana, E.-M.; Alemneh, A.A. Rhizobia strain and legume genome interaction effects on nitrogen fixation and yield of grain legume: A review. Mol. Soil Biol. 2015, 6, 1–6. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, B.X.; Ren, H.; Zhang, J. Composition and functional diversity of microbial community across a mangrove-inhabited mudflat as revealed by 16S rDNA gene sequences. Sci. Total Environ. 2018, 633, 518–528. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Concentration |
|---|---|
| Chemical oxygen demand (COD) | 265 mg/L |
| Ammonium | 30 mg N/L |
| Total nitrogen (TN) | 38–39 mg N/L |
| Seawater | 18% |
| Cl− | 3700–3900 mg/L |
| SO42− | 160–180 mg/L |
| Time (day) | DO Level (mg/L) | Temp. (°C) | TN Removal (%) | NH4+ Removal (%) | COD Removal (%) |
|---|---|---|---|---|---|
| 0–40 | 0.5 | 20 | 31.89 ± 5.63 | 95.73 ± 3.42 | 86.57 ± 2.44 |
| 41–55 | 0.3 | 20 | 60.67 ± 1.36 | 97.91 ± 2.96 | 93.92 ± 0.79 |
| 56–78 | 0.2 | 20 | 52.92 ± 16.67 | 94.09 ± 5.15 | 94.80 ± 0.76 |
| 79–146 | 0.2 | 30 | 54.52 ± 7.24 | 97.01 ± 2.35 | 91.72 ± 6.60 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuang, H.; Wu, Z.; Xu, L.; Leu, S.-Y.; Lee, P.-H. Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities. Microorganisms 2020, 8, 919. https://doi.org/10.3390/microorganisms8060919
Zhuang H, Wu Z, Xu L, Leu S-Y, Lee P-H. Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities. Microorganisms. 2020; 8(6):919. https://doi.org/10.3390/microorganisms8060919
Chicago/Turabian StyleZhuang, Huichuan, Zhuoying Wu, Linji Xu, Shao-Yuan Leu, and Po-Heng Lee. 2020. "Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities" Microorganisms 8, no. 6: 919. https://doi.org/10.3390/microorganisms8060919
APA StyleZhuang, H., Wu, Z., Xu, L., Leu, S.-Y., & Lee, P.-H. (2020). Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities. Microorganisms, 8(6), 919. https://doi.org/10.3390/microorganisms8060919