Incidental Prophylactic Appendectomy Is Associated with a Profound Microbial Dysbiosis in the Long-Term

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Analysis of Anthropometric and Biochemical Variables
2.3. DNA and RNA Extraction
2.4. Microbiota Analysis
2.5. Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
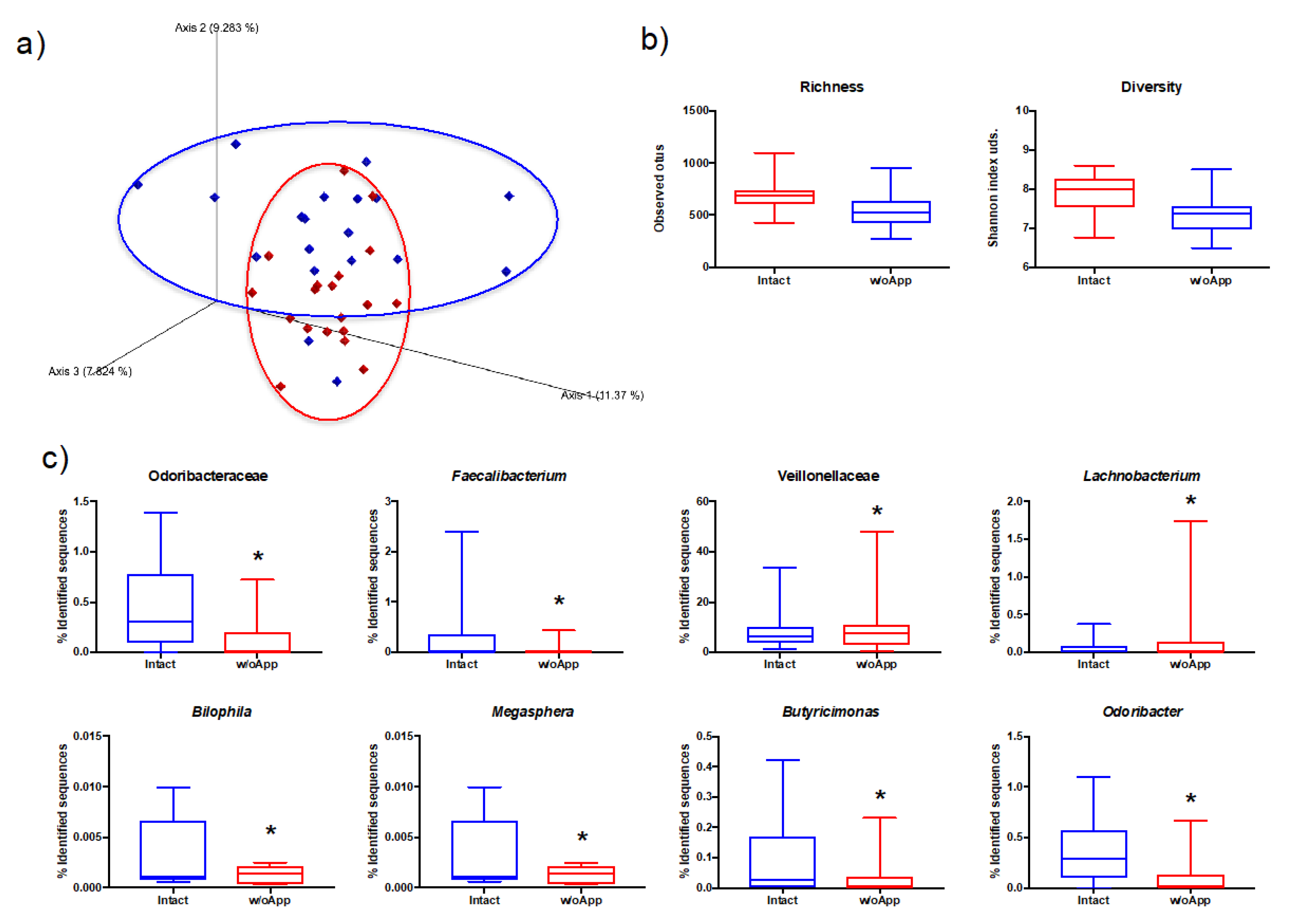
3.1. Intact and Appendectomized Patients Differed in Their Gut Microbiota Populations
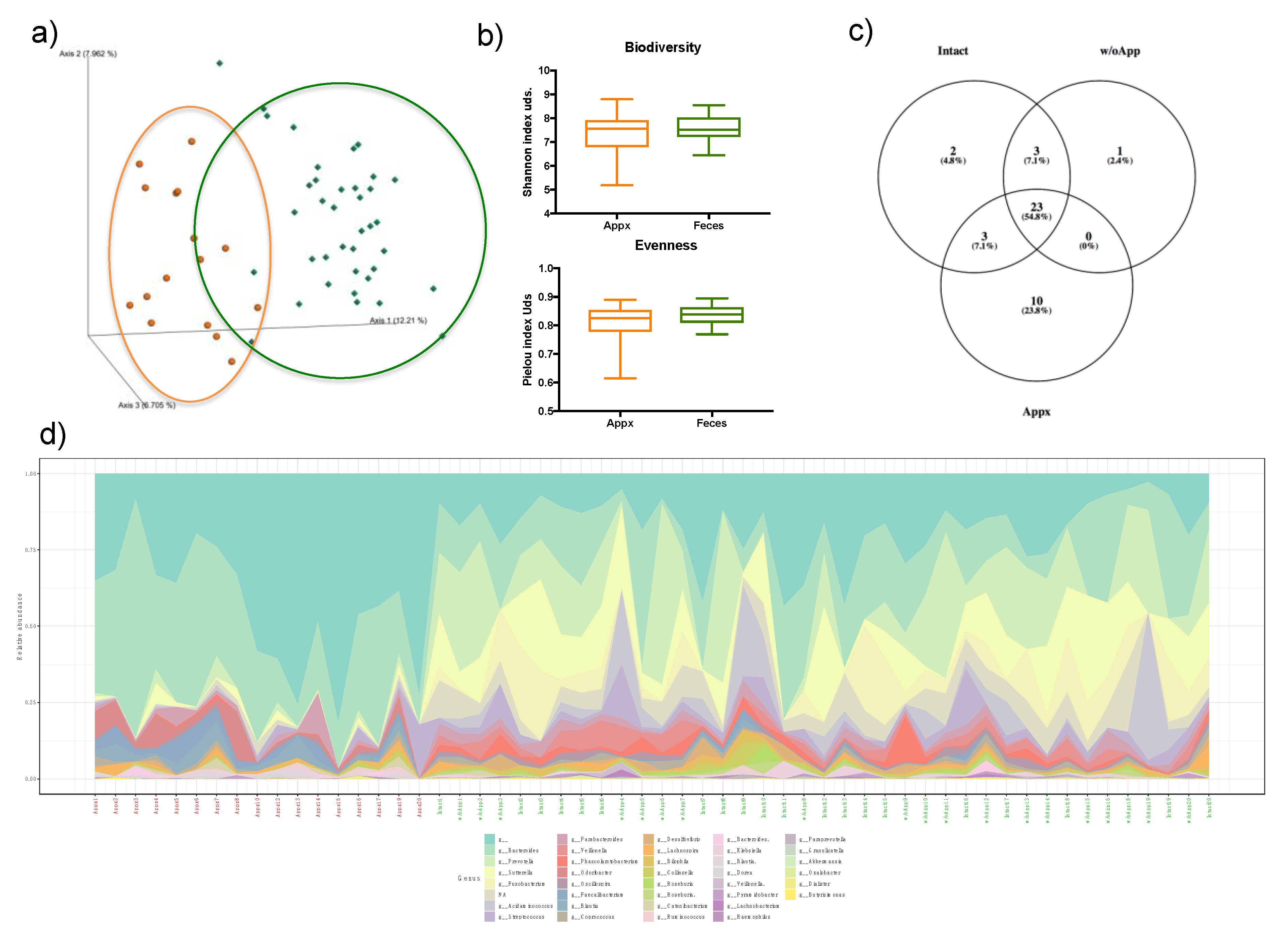
3.2. Appendix and Stool Microbiota Populations
3.3. Odoribacter and Butyricimonas are Related to Insulin Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, C.R.; Trevatt, A.E.J.; Dixit, A.; Datta, V. Systematic review of clinical outcomes after prophylactic surgery. Ann. R. Coll. Surg. Engl. 2016, 98, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Livingston, E.H.; Woodward, W.A.; Sarosi, G.A.; Haley, R.W. Disconnect between incidence of nonperforated and perforated appendicitis: Implications for pathophysiology and management. Ann. Surg. 2007, 245, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Kooij, I.A.; Sahami, S.; Meijer, S.L.; Buskens, C.J.; te Velde, A.A. The immunology of the vermiform appendix: A review of the literature. Clin. Exp. Immunol. 2016, 186, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Randal Bollinger, R.; Barbas, A.S.; Bush, E.L.; Lin, S.S.; Parker, W. Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theor. Biol. 2007, 249, 826–831. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Cardona, F.; Tinahones, F.J.; Queipo-Ortuno, M.I. Impact of the gut microbiota on the development of obesity and type 2 diabetes mellitus. Front. Microbiol. 2014, 5, 190. [Google Scholar] [CrossRef]
- Adak, A.; Khan, M.R. An Insight into Gut Microbiota and its Functionalities. Cell. Mol. Life Sci. Cmls 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Anders, S.; McCarthy, D.J.; Chen, Y.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J. Venny. An interactive tool for comparing lists with Venn’s diagrams. 2007–2015.
- Mehran, A.; Liberman, M.; Rosenthal, R.; Szomstein, S. Ruptured Appendicitis after Laparoscopic Roux-enY Gastric Bypass: Pitfalls in Diagnosing a Surgical Abdomen in the Morbidly Obese. Obes. Surg. 2003, 13, 938–940. [Google Scholar] [CrossRef] [PubMed]
- Dilek, O.N.; Guler, O.; Guler, A.A.; Demirtas, I.; Altindis, M.; Dilek, F.H.; Ozgoren, E. Prophylactic appendectomy: Is it worth to be done? Acta Chir. Belg. 2001, 101, 65–67. [Google Scholar] [PubMed]
- Rajani, C.; Jia, W. Disruptions in gut microbial-host co-metabolism and the development of metabolic disorders. Clin. Sci. (Lond. Engl. 1979) 2018, 132, 791–811. [Google Scholar] [CrossRef]
- Krishnan, S.; Alden, N.; Lee, K. Pathways and functions of gut microbiota metabolism impacting host physiology. Curr. Opin. Biotechnol. 2015, 36, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.D.; Young, W.; Maclean, P.H.; Cookson, A.L.; Bermingham, E.N. Metagenomic insights into the roles of Proteobacteria in the gastrointestinal microbiomes of healthy dogs and cats. MicrobiologyOpen 2018, 7, e00677. [Google Scholar] [CrossRef]
- Joshi, T.; Elderd, B.D.; Abbott, K.C. No appendix necessary: Fecal transplants and antibiotics can resolve Clostridium difficile infection. J. Theor. Biol. 2018, 442, 139–148. [Google Scholar] [CrossRef]
- Reese, A.T.; Dunn, R.R. Drivers of Microbiome Biodiversity: A Review of General Rules, Feces, and Ignorance. mBio 2018, 9, e01294–e01318. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M. Increased Systolic and Diastolic Blood Pressure Is Associated with Altered Gut Microbiota Composition and Butyrate Production in Early Pregnancy. Hypertension (Dallas Tex. 1979) 2016, 68, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Tanaka, Y.; Benno, Y.; Ohkuma, M. Butyricimonas faecihominis sp. nov. and Butyricimonas paravirosa sp. nov., isolated from human faeces, and emended description of the genus Butyricimonas. Int. J. Syst. Evol. Microbiol. 2014, 64, 2992–2997. [Google Scholar] [CrossRef] [PubMed]
- Cushing, K.; Alvarado, D.M.; Ciorba, M.A. Butyrate and Mucosal Inflammation: New Scientific Evidence Supports Clinical Observation. Clin. Transl. Gastroenterol. 2015, 6, e108. [Google Scholar] [CrossRef] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Whitford, M.F.; Yanke, L.J.; Forster, R.J.; Teather, R.M. Lachnobacterium bovis gen. nov., sp. nov., a novel bacterium isolated from the rumen and faeces of cattle. Int. J. Syst. Evol. Microbiol. 2001, 51, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.E.; Dai, Y. Metabolome of human gut microbiome is predictive of host dysbiosis. GigaScience 2015, 4, 42. [Google Scholar] [CrossRef]
- Bernard, D.; Verschraegen, G.; Claeys, G.; Lauwers, S.; Rosseel, P. Bilophila wadsworthia bacteremia in a patient with gangrenous appendicitis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1994, 18, 1023–1024. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.-L.; Rainteau, D.; Bridonneau, C.; da Costa, G.; van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; García-Fuentes, E.; Cardona, F.; Queipo-Ortuño, M.I.; Tinahones, F.J. Insulin resistance is associated with specific gut microbiota in appendix samples from morbidly obese patients. Am. J. Transl. Res. 2016, 8, 5672–5684. [Google Scholar]
- Lee, H.; Lee, Y.; Kim, J.; An, J.; Lee, S.; Kong, H.; Song, Y.; Lee, C.-K.; Kim, K. Modulation of the gut microbiota by metformin improves metabolic profiles in aged obese mice. Gut Microbes 2018, 9, 155–165. [Google Scholar] [CrossRef]
- Yamashita, M.; Okubo, H.; Kobuke, K.; Ohno, H.; Oki, K.; Yoneda, M.; Tanaka, J.; Hattori, N. Alteration of gut microbiota by a Westernized lifestyle and its correlation with insulin resistance in non-diabetic Japanese men. J. Diabetes Invest. 2019. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Intact Group | w/oApp Group | p-Value | ||||
|---|---|---|---|---|---|---|
| preSurgery | StudyTime | preSurgery | StudyTime | preSurgery | StudyTime | |
| Age (years) | 43.5 ± 9.12 | 53.65 ± 8.87 | 38.73 ± 9.25 | 49.45 ± 9.98 | ||
| Gender (M/F) | 8/12 | 5/15 | ||||
| BMI (kg/m2) | 52.36 ± 7.33 | 36.97 ± 7.09 * | 54.80 ± 3.81 | 36.10 ± 6.13 * | 0.159 | 0.682 |
| Waist (cm) | 138.26 ± 15.56 | 116.84 ± 16.97 * | 137.00 ± 12.34 | 111.8 ± 13.50 * | 0.850 | 0.305 |
| Glucose (mg/dL) | 119.74 ± 39.86 | 98.35 ± 37.25 * | 113.72 ± 21.84 | 94.55 ± 24.46 * | 0.558 | 0.705 |
| Cholesterol (mg/dL) | 206.21 ± 36.25 | 151.40 ± 39.30 * | 200.06 ± 40.78 | 140.45 ± 32.91 * | 0.618 | 0.345 |
| HDL-Chol | 47.37 ± 12.68 | 56.60 ± 15.50 * | 43.94 ± 10.19 | 61.10 ± 14.90 * | 0.283 | 0.355 |
| TG (mg/dL) | 133.94 ± 57.69 | 103.75 ± 58.99 | 143.12 ± 83.61 | 90.55 ± 37.75 * | 0.851 | 0.405 |
| SBP (mmHg) | 141.17 ± 24.86 | 122.10 ± 14.87 * | 141.07 ± 20.20 | 120.95 ± 14.03 * | 0.813 | 0.803 |
| DBP (mmHg) | 81.94 ± 15.72 | 66.05 ± 7.21 * | 84.93 ± 9.51 | 64.85 ± 8.76 * | 0.435 | 0.639 |
| Insulin (mg/dL) | 23.08 ± 12.41 | 7.93 ± 4.73 * | 21.78 ± 12.91 | 15.70 ± 22.51 | 0.585 | 0.149 |
| HOMA-IR | 6.38 ± 3.02 | 2.13 ± 1.81 * | 6.21 ± 4.25 | 4.21 ± 6.67 * | 0.730 | 0.193 |
| CRP (mg/dL) | 6.28 ± 7.64 | 3.44 ± 0.95 | 5.32 ± 3.64 | 4.25 ± 2.71 | 0.656 | 0.217 |
| Insulin Levels | ||||||
|---|---|---|---|---|---|---|
| Intact (R = 0.717, R2 adj = 0.204, p = 0.218) | w/oApp (R = 0.818, R2 adj = 0.511, p = 0.027) | |||||
| ß | p-Value | 95% CI | ß | p-Value | 95% CI | |
| Sex | −0.370 | 0.145 | −8.508–1.431 | 0.003 | 0.987 | −22.142–22.470 |
| Age | 0.340 | 0.165 | −0.090–0.465 | −0.163 | 0.404 | −1.335–0.580 |
| BMI | 0.345 | 0.144 | −0.093–0.563 | −0.075 | 0.692 | −1.756–1.208 |
| Bilophila | 0.465 | 0.115 | −0.671–5.350 | −0.169 | 0.455 | −60.468–28.983 |
| Butyricimonas | 0.101 | 0.756 | −20.423–27.339 | 0.630 | 0.011 | 71.340–448.017 |
| Lachnobacterium | 0.154 | 0.614 | −24.290–39.289 | −0.039 | 0.855 | −27.116–22.873 |
| Odoribacter | −0.191 | 0.634 | −14,455–9.189 | 0.470 | 0.057 | −2.180–132.586 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Alcoholado, L.; Fernández-García, J.C.; Gutiérrez-Repiso, C.; Bernal-López, M.R.; Ocaña-Wilhelmi, L.; García-Fuentes, E.; Moreno-Indias, I.; Tinahones, F.J. Incidental Prophylactic Appendectomy Is Associated with a Profound Microbial Dysbiosis in the Long-Term. Microorganisms 2020, 8, 609. https://doi.org/10.3390/microorganisms8040609
Sánchez-Alcoholado L, Fernández-García JC, Gutiérrez-Repiso C, Bernal-López MR, Ocaña-Wilhelmi L, García-Fuentes E, Moreno-Indias I, Tinahones FJ. Incidental Prophylactic Appendectomy Is Associated with a Profound Microbial Dysbiosis in the Long-Term. Microorganisms. 2020; 8(4):609. https://doi.org/10.3390/microorganisms8040609
Chicago/Turabian StyleSánchez-Alcoholado, Lidia, José Carlos Fernández-García, Carolina Gutiérrez-Repiso, M Rosa Bernal-López, Luis Ocaña-Wilhelmi, Eduardo García-Fuentes, Isabel Moreno-Indias, and Francisco J. Tinahones. 2020. "Incidental Prophylactic Appendectomy Is Associated with a Profound Microbial Dysbiosis in the Long-Term" Microorganisms 8, no. 4: 609. https://doi.org/10.3390/microorganisms8040609
APA StyleSánchez-Alcoholado, L., Fernández-García, J. C., Gutiérrez-Repiso, C., Bernal-López, M. R., Ocaña-Wilhelmi, L., García-Fuentes, E., Moreno-Indias, I., & Tinahones, F. J. (2020). Incidental Prophylactic Appendectomy Is Associated with a Profound Microbial Dysbiosis in the Long-Term. Microorganisms, 8(4), 609. https://doi.org/10.3390/microorganisms8040609