Abstract
Ticks and tick-borne diseases (TBDs) have a major economic impact on animal production worldwide. In the present study, 2410 ticks were collected from January to August 2017 from livestock and other domestic animals in North Kordofan and Kassala, Sudan, for species identification and investigation of Rickettsia spp. and piroplasms, either individually or as pools containing up to 10 ticks by molecular methods. In total, 13 tick species were identified by morphology and 16S rDNA sequencing. The most frequent tick species were Hyalomma impeltatum (24.90%), Rhipicephalus evertsi evertsi (18.84%), Amblyomma lepidum (16.06%), and Rhipicephalus camicasi (12.49%). A pan-Rickettsia real-time PCR revealed an overall minimum infection rate (MIR) with Rickettsia spp. of 5.64% (136 positive tick pools/2410 total ticks). Rickettsia africae and Rickettsia aeschlimannii were the most frequently identified species by sequencing. Furthermore, the following highly pathogenic livestock parasites were detected: Theileria annulata, Theileria lestoquardi, Theileria equi, and Babesia caballi. The present study documented Rhipicephalus afranicus as well as Rickettsia conorii israelensis, Rickettsia massiliae, and Babesia pecorum for the first time in Sudan. These findings are significant for the animal production sector as well as in terms of One Health, as the detected Rickettsia spp. can cause serious illness in humans.
1. Introduction
Ticks are medically important ectoparasites of mammals, birds, reptiles, and amphibians. Their haematophagous feeding habit may result in blood loss and mechanical damage to the skin [1,2]. Furthermore, ticks can transmit many pathogens to humans and animals, including viruses (e.g., Crimean–Congo haemorrhagic fever (CCHF) virus), bacteria (e.g., Rickettsia spp.) protozoans (e.g., Babesia spp. and Theileria spp.) and helminths [3]. Ticks and tick-borne diseases (TBDs) result in economic losses that are estimated at USD 22–30 billion globally every year [4].
The burden and dynamics of ticks and TBDs are changing with the concurrent global climate change [5]. Moreover, anthropogenic factors can influence the biodiversity and population dynamics of ticks [5]. Consequently, a substantial increase in TBDs has been noted in certain regions, e.g., human cases of TBDs more than doubled in the United States from 2004 to 2016 [6]. Furthermore, some ticks have expanded their habitats to new exotic areas [7]. The introduction of novel tick species may lead to cascading effects on the local tick fauna and TBDs. This is clearly illustrated by the introduction of Rhipicephalus microplus, vector of the cattle parasites Babesia bovis and Babesia bigemina, into several African countries, which led to displacement of native Rhipicephalus spp. and may result in increased Babesia transmission [8,9].
Overall, the reported tick species in Africa belong to 10 different genera, including Amblyomma, Hyalomma, Rhipicephalus, Ixodes, Argas, and Ornithodoros, for example [10]. Tick-borne pathogens of livestock, such as the causative agents of theileriosis, babesiosis and heartwater, have been circulating on the continent, often with a considerable economic cost [11,12,13]. Additionally, zoonotic TBDs, including rickettsioses, pose a serious public health problem [14,15]. Therefore, a contemporary tick and TBD surveillance is relevant in African countries.
Sudan has a huge wealth of livestock that is estimated to be more than 100 million animals [16]. A substantial proportion of the people of Sudan own livestock for subsistence [17]. These people are vulnerable to the negative impact of animal diseases, including TBDs, and ticks. Previous studies have shown a high prevalence of Theileria spp. in Sudanese livestock, especially regarding the highly pathogenic Theileria parva in cattle [18] and Theileria equi in horses [19]. Furthermore, tick-transmitted Rickettsia spp., especially Rickettisa africae, which causes African tick-borne fever, have a high prevalence in the region and represent a public health hazard [15,20,21]. Moreover, in the past few years, changes in the tick fauna have been noted in Sudan [17], probably due to the continuous movement of animals in search of pastures and water as well as due to natural and anthropogenic changes in environment [17]. This highlights the need to monitor how the natural tick population is changing in Sudan, particularly in animal production areas. In this study, we collected 2410 ticks from livestock and other domestic animals in North Kordofan and Kassala states, Sudan, for species identification and investigation of Rickettsia spp., Theileria spp., and Babesia species.
2. Materials and Methods
2.1. Study Area
North Kordofan state is located in the central part of Sudan between latitudes 11° and 16° N and longitudes 27° and 32° E, with a total area of 185,302 km2, while Kassala state is located in the eastern part of the country between latitude 14° and 17° N and longitude 34° and 37° E, covering an area of 42,282 km2 (Figure 1). Annual rainfall (up to 700 mm/year) is concentrated in a single relatively short season from June to September (hot rainy season), followed by a cold dry season from October to January and a hot dry season from February to May. Up to 40% of the states’ total land is cultivable and agriculture and livestock comprises up to 70% of the economic activities. The states have abundant fodder, grazing areas, and water sources, like seasonal rivers (e.g., Kour Abu-Habil in North Kordofan and Atbara and Gaash in Kassala) and hafeers (i.e., rainwater harvesting sites), during the rainy season. This supports a considerably large livestock population, which is estimated at 13,061,246 head in North Kordofan and 4,479,050 in Kassala [16]. Animals are raised in a mixture of farming systems, such as mixed crop–livestock, nomadic, sedentary, and semi-sedentary, for domestic consumption and for export to international markets (e.g., Gulf countries).
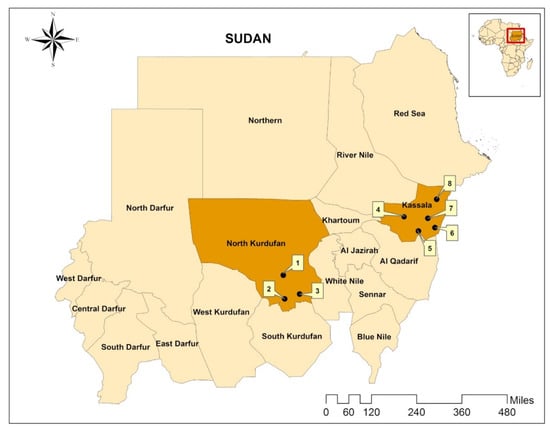
Figure 1.
Map showing the location of Sudan in Africa (small map) and the location of the study areas in Sudan (North Kordofan and Kassala states, indicated by orange). Sampling was conducted in Sheikan (1), Al-Rahad (2), Um-Ruwabah (3), West Kassala (4), Kashm el Griba (5), Kassala (6), Aroomah (7) and Wagar (8). The map was created using ArcGIS v. 10 (esri Inc., Redlands, CA, USA).
2.2. Tick Collection and Identification
In this cross-sectional study, tick specimens were collected from livestock and other domestic animals in the period from January to August 2017, in North Kordofan and Kassala states, Sudan [14]. Samples were collected from three localities in the state of North Kordofan, including Sheikan, Al-Rahad, and Um-Ruwabah, and from five localities in the state of Kassala, namely Aroomah, Wagar, Kassala, West Kassala, and Khashm el Griba (Figure 1). These localities were selected randomly and/or conveniently. In both areas, mostly sheep and goats were examined, followed by cattle, camels and dogs. Sampled animals were thoroughly examined for attached ticks by searching the head and ears, the neck (dewlap), the thoracic area and the abdomen, the udder or scrotum, the fore- and hindlimbs, perineum, and the tail. Attached ticks were either collected by hand- or forceps-picking and stored in 70% ethanol. All ticks collected from the same animal host were put into one tube. Tubes were labelled (location, animal species and date of collection) and sent to the Bundeswehr Institute of Microbiology, Munich, Germany, where ticks were identified to species level using morphological characteristics described by Apanaskevich and Horak [22], Apanaskevich and Horak [23], Apanaskevich and Horak [24], Apanaskevich, et al. [25], Voltzit and Keirans [26] and Walker, et al. [10,27].
2.3. Nucleic Acid Extraction
Total nucleic acid was extracted using the MagNA Pure LC RNA/DNA Kit (Roche, Mannheim, Germany) in a MagNA Pure LC instrument (Roche) according to the manufacturer’s instructions. Total nucleic acid was extracted from individual ticks or pools containing 2–10 specimens per pool, if the ticks shared the same developmental stage and species and were collected from the same animal. The extracted total nucleic acid was stored at −80 °C until use.
2.4. Molecular Tick Species Identification
Identification of ticks that were either damaged or fully engorged, and thus not reliably identifiable based on morphological criteria (n = 23), as well as confirmation of primary morphological determinations (n = 13), was achieved by 16S rDNA sequencing (250 bp fragment) and phylogenetic analysis. The gene was amplified using polymerase chain reaction (PCR) protocols and sequenced in both directions as previously described by Mangold, et al. [28]. Tick sequences generated in this study are available in GenBank (MT535883-MT535906). Additional sequences from GenBank were chosen to cover the range of Rhipicephalus and Hyalomma species that occur in Sudan and closely related species. As the prevalence of misidentified tick species among sequence data in GenBank is a growing problem, the selected sequences were derived from recent studies that included large-scale taxonomic investigations to verify identification by phylogenetic analysis and correlated morphology [29,30,31,32,33,34]. Sequence data were aligned using MAFFT (Q-INS-I, 200PAM/k = 2, Gap opening penalty 1.53) [35], and the final alignment comprised 265 nucleotide characters. The alignment was inspected manually to ensure sequences were in reading frame. Phylogenetic analysis was based on maximum likelihood with 1000 bootstrap replicates in MEGA v7.0.14 [36] using a TPM2u + F + G4 model determined by Bayesian Information Criterion calculations in W-IQ-TREE [37].
2.5. PCR for Rickettsia spp. and Piroplasms
For detection of Rickettsia spp., a pan-Rickettsia real-time PCR was used [38,39]. Positive samples were further subjected to Rickettsia species identification by amplification, sequencing in both directions and phylogenetic analysis of the 23S-5S intergenic spacer region (330 bp fragment) as described by Chitimia-Dobler, et al. [40]. Additional sequences from GenBank were chosen to cover the range of Rickettsia species that occur in Africa and Eurasia [41,42]. Sequence data were aligned using MAFFT (Q-INS-I, 200PAM/k = 2, Gap opening penalty 1.53) [35], and the final alignment comprised 403 nucleotide characters. The alignment was inspected manually to ensure sequences were in reading frame. Phylogenetic analysis was based on maximum likelihood with 1000 bootstrap replicates in MEGA v7.0.14 [36] using an HKY + F + G4 model determined by Bayesian Information Criterion calculations in W-IQ-TREE [37].
To identify whether the collected ticks were infected with piroplasms, the pools were tested for Theileria spp. and Babesia spp. DNA by amplifying a part of the 18S rDNA in a conventional PCR, using the primers BJ1 and BN2 [43], as described by Springer, et al. [44]. Obtained 18S rDNA amplicons were custom Sanger-sequenced (Microsynth Seqlab Sequencing Laboratories, Göttingen, Germany), or—in case of weak bands—ligated into the pCR™4-TOPO® TA vector and cloned into One Shot Top10 chemically competent Escherichia coli (TOPO® TA Cloning kit, Thermo Fisher Scientific GmbH, Dreieich, Germany). After plasmid extraction and purification (NucleoSpin Plasmid kit, Macherey-Nagel GmbH & Co. KG, Düren, Germany), the insert was custom Sanger-sequenced, as indicated above. Rickettsia spp. and piroplasms’ sequences generated in this study are available in GenBank under the accession numbers MW152276–MW152327 for Rickettsia spp., and MW131349–MW131365 for piroplasms. Minimum infection rates (MIRs) were calculated under the assumption of only one positive tick per pool (MIR = number of positive pools/total number of ticks).
3. Results
3.1. Identified Tick Species
In total, 2410 ticks, including 1301 from North Kordofan and 1109 from Kassala, were collected from cattle, sheep, goats, camels and horses. Based on morphological characteristics and 16S rDNA sequencing, 13 different tick species belonging to three genera were identified (i.e., Hyalomma (998/2410, 41.41%), Amblyomma (445/2410, 18.46%), and Rhipicephalus (967/2410, 40.12%)). Overall, Hyalomma impeltatum was most frequently identified (600/2410, 24.90%), followed by Rhipicephalus evertsi evertsi (454/2410, 18.84%), Amblyomma lepidum (387/2410, 16.06%), Rhipicephalus camicasi (301/2410, 12.49%), and Hyalomma anatolicum (185/2410, 7.68%). The remaining eight species were found in proportions ranging from 0.04% to 7.68%. Detailed data on tick species and the number of male, female and nymphal ticks of each species are presented in Table 1.

Table 1.
Tick species collected from livestock and other domestic animals in Sudan and their pathogen infection rates.
In North Kordofan, H. impeltatum was most abundant (587/1301, 45.11%), followed by A. lepidum (241/1301, 18.52%) and Rh. decoloratus (116/1301, 8.92%) (Figure 2A). In contrast, Rh. evertsi evertsi (373/1109, 33.63%), Rh. camicasi (234/1109, 21.10%) and H. anatolicum (176/1109, 15.87%) were the main identified tick species in Kassala. A. variegatum (58/1301, 4.46%) and H. truncatum (6/1301, 0.46%) were found only in North Kordofan, while Rh. afranicus was found only in Kassala (1/1109, 0.09%).
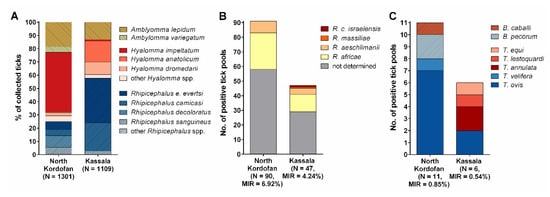
Figure 2.
Species composition of ticks and tick-borne pathogens detected in North Kordofan and Kassala, Sudan. (A) Species composition of ticks collected from livestock and other domestic animals from January to August 2017. (B) Rickettsia species composition among pan-Rickettsia real-time PCR-positive tick pools. (C) Piroplasm species composition among PCR-positive tick pools. Pathogenic species are indicated in shades of red, while apathogenic species are shown in shades of blue. MIR = minimum infection rate.
Phylogenetic analysis of the 16S rDNA sequences confirmed the morphology-based species determination for eight damaged or fully engorged Hyalomma ticks, namely H. anatolicum (n = 1), H. dromedarii (n = 2), and H. impeltatum (n = 5) (Figure 3). Moreover, the species of 18 Rhipicephalus ticks, including Rh. sanguineus s.l. tropical lineage (n = 7), Rh. afranicus (n = 1), Rh. camicasi (n = 7), Rh. evertsi evertsi (n = 1), Rh. decoloratus (n = 1) and Rh. microplus (n = 1), could only be identified by Sanger-sequencing (Figure 4).
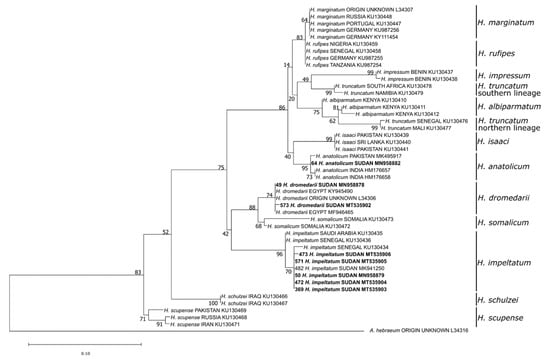
Figure 3.
Maximum likelihood phylogenetic analysis of 16S rDNA sequences of Hyalomma spp. ticks using Amblyomma hebraeum as an outgroup. Bolded tip labels refer to sequences generated in this study, and sample ID, species/lineage names, country of origin and GenBank accession numbers are indicated. Nodal values indicate bootstrap support using 1000 replicates.
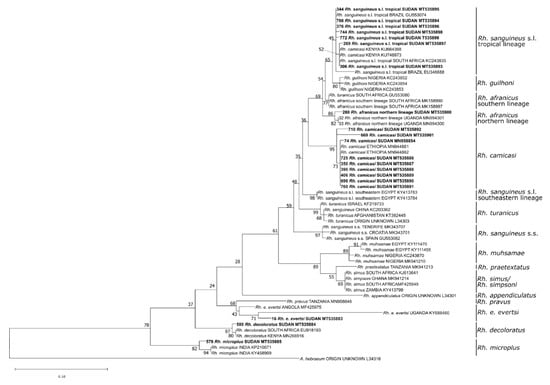
Figure 4.
Maximum likelihood phylogenetic analysis of 16S rDNA sequences of Rhipicephalus spp. ticks using Amblyomma hebraeum as an outgroup. Bolded tip labels refer to sequences generated in this study, and sample ID, species/lineage names, country of origin, and GenBank accession numbers are indicated. Nodal values indicate bootstrap support using 1000 replicates.
3.2. Prevalence of Tick-Borne Pathogens
3.2.1. Rickettsia Species
In total, 783 tick pools were tested for Rickettsia species by real-time PCR. Of these, 136 were Rickettsia-positive, resulting in an MIR of 5.64% (136/2410). Rickettsia DNA was detected in 11 out of 13 tick species (Table 1). Rickettsia species composition among the positive tick pools from North Kordofan and Kassala is shown in Figure 2B.
In Amblyomma spp., the MIR was 12.13% (54/445), and sequencing of the 23S-5S IGS region confirmed Rickettsia africae in 37 Amblyomma pools (Figure 5). In the remaining 17 samples, the Rickettsia DNA content was too low for species identification. The MIR in Hyalomma spp. was 4.4% (44/998). Twelve of the 44 samples were successfully sequenced, leading to the identification of Rickettsia aeschlimannii. Among the observed Rhipicephalus spp., the MIR was 3.93% (38/967). Unfortunately, the majority of Rickettsia-positive Rhipicephalus samples did not contain enough Rickettsia DNA for 23S-5S sequencing. Regardless, Rickettsia conorii israelensis was detected in two Rh. camicasi pools and R. aeschlimannii in one Rh. evertsi evertsi pool. The single Rh. afranicus specimen was also Rickettsia-positive, and subsequent sequencing identified Rickettsia massiliae.
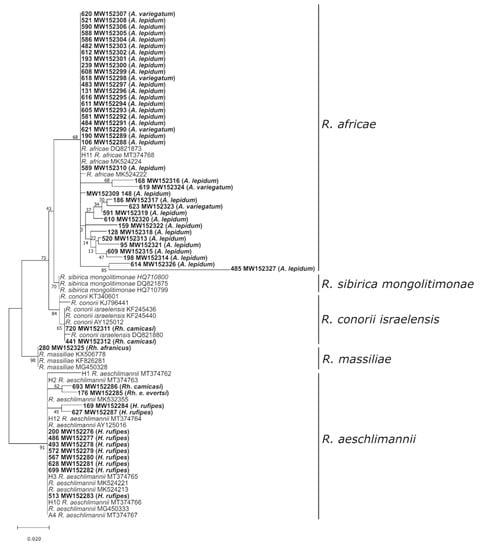
Figure 5.
Maximum likelihood phylogenetic analysis of 5S-23S intergenic spacer sequences obtained from Rickettsia species. Bolded terminals refer to sequences generated in this study, and sample ID, GenBank accession numbers, and tick host are indicated. Nodal values indicate bootstrap support using 1000 replicates.
3.2.2. Piroplasms
PCR for detection of piroplasms revealed overall MIRs of 0.58% (14/2410) and 0.12% (3/2410) for Theileria spp. and Babesia spp., respectively (Table 1). Piroplasm species composition among the positive tick pools from North Kordofan and Kassala is shown in Figure 2C.
Theileria spp. DNA was detected in H. anatolicum (MIR = 2.16%, 4/185), H. impeltatum (MIR = 0.67%, 4/600), Rh. evertsi evertsi (MIR = 0.88%, 4/454), Rh. camicasi (MIR = 0.33%, 1/301), and A. lepidum (MIR = 0.26%, 1/387). Five different Theileria spp. were identified by 18S rDNA sequencing, as shown in Table 2, including Theileria ovis, Theileria annulata, Theileria equi, Theileria lestoquardi, and Theileria velifera.
Table 2.
Babesia spp. and Theileria spp. identified in different tick species collected from livestock and other domestic animals in Sudan from January to August 2017.
Babesia spp. DNA was detected in H. impeltatum (MIR = 0.33%, 2/600) and A. variegatum (MIR 1.72%, 1/58). The 18S rDNA Babesia sequences from H. impeltatum showed 98–99% identity to Babesia pecorum while the sequences from A. variegatum showed 97% identity to B. caballi (Table 2).
4. Discussion
Ticks and TBDs constitute a global economic and health burden. In countries with a socioeconomic status similar to that of Sudan, a substantial proportion of livestock are owned by subsistence farmers, who are especially vulnerable to the impact of ticks and TBDs [45]. Hassan and Salih [17] stated that the natural population of ticks infesting livestock is changing in Sudan. Therefore, monitoring of the local tick fauna is necessary. In this study, we classified 2410 ticks collected from livestock and other domestic animals in two regions in Sudan into 13 different tick species belonging to the genera Hyalomma, Amblyomma, and Rhipicephalus. Tick screening for Rickettsia spp. and piroplasms revealed Rickettsia spp., like R. africae and R. aeschlimannii, as well as several piroplasms of veterinary relevance. Interestingly, we report Rhipicephalus afranicus, Rickettsia conorii israelensis, Rickettsia massiliae, and Babesia pecorum for the first time in Sudan. These findings are of high significance for the animal and public health sectors, particularly from a One Health point of view, as rickettsiosis is an important zoonotic disease.
With the exception of Rh. afranicus, which can experimentally transmit Babesia trautmanni to pigs [29], all of the other detected tick species have formerly been reported in Sudan [15,46,47,48]. Both sampling areas are characterized primarily by Sahelian dry savannah ecosystems; nevertheless, regional differences were noted. Although there was variation in the species composition of the examined host populations, limiting comparability between both regions, the majority of examined animals in both regions were sheep, goats and cattle. Therefore, it was remarkable that the tick fauna in North Kordofan was dominated by Hyalomma spp., while Rhipicephalus spp. were the most frequent ticks in Kassala. H. anatolicum, the main vector of T. annulata, which causes bovine tropical theileriosis, has undergone a south- and west-ward spread in Sudan since the 1980s, probably due to animal movement and environmental change [17]. Recently, H. anatolicum represented more than 50% of collected ticks in West Darfur, Al-Jazeera, and River Nile states [15]. This indicates that the distribution of H. anatolicum has reached the western border of Sudan. In the present study, it was the most frequently observed Hyalomma spp. in Kassala, but was also collected in North Kordofan, coinciding with the detection of bovine tropical theileriosis in North Kordofan [49].
In Kassala, Rh. evertsi evertsi and Rh. camicasi together accounted for more than 50% of collected ticks. Similarly, Rh. evertsi evertsi was frequently encountered on cattle in Gezira, central Sudan, and on different domestic animals in West Darfur and River Nile [15]. Both species were also reported by Jongejan, et al. [46] in the Blue and White Nile ecosystems. However, Rh. camicasi can be difficult to distinguish morphologically from other ticks of the Rh. sanguineus group, which may explain why this species has been less frequently reported in other studies [48,50].
Contrary to the findings of Shuaib, et al. [15], Elghali and Hassan [51] and Ahmed, et al. [50], A. lepidum was found in both states, North Kordofan and Kassala, and accounted for approximately 16% of all identified ticks. This tick has historically been abundant in the eastern part of the country, such as Kassala [17]. In recent decades, a westward (towards Kordofan and Darfur regions) spread of A. lepidum has been observed [17]. Indeed, the importance of A. lepidum lies in the fact that it is the main vector of Ehrlichia ruminantium, the causative agent of heartwater, which results in significant morbidities and mortalities in domestic ruminants [52].
One Rh. afranicus specimen, collected from a sheep, was identified by sequencing of the 16S rRNA gene. This taxon was historically confounded with Rhipicephalus turanicus, however, it was recently described as a distinct species [29]. It has further been confirmed in South Africa [29] and Uganda [53] to date. These Rh. afranicus populations may represent two distinct lineages within the species given molecular distances between southern (i.e., South Africa) and northern (i.e., Uganda, Sudan) regions [53]. Interestingly, this specimen carried R. massiliae DNA.
The most relevant tick-borne rickettsiae in Africa are R. africae, primarily transmitted by Amblyomma spp., R. aeschlimannii, mainly transmitted by Hyalomma spp., and R. conorii conorii, which is transmitted by Rhipicephalus ticks [54]. In this study, the detected MIR (12.13%) of Amblyomma ticks with Rickettsia spp. and the confirmation of R. africae in the majority of samples denote to a considerable risk of infection of humans with R. africae, the causative agent of African tick-bite fever. Similar infection rates of ticks with R. africae have been noted in Sudan before [15]. However, high rickettsial infection rates of up to 100% have been described in Amblyomma spp. in other regions of Africa, probably due to effective transovarial transmission [54]. Furthermore, MIRS of 4.4% and 3.9% were detected in Hyalomma and Rhipicephalus ticks, respectively. In previous studies from eastern Africa, these tick genera mostly showed a lower Rickettsia prevalence than Amblyomma spp., ranging from approximately 10 to 46% in Hyalomma spp. [55,56,57] and 0 to 1.1% in Rhipicephalus spp. [55,57]. Species identification was only possible in approximately one third of the positive Hyalomma pools and the pathogen was confirmed as R. aeschlimannii. In the same way, the low rickettsial DNA content did not allow for species identification in most of the Rhipicephalus samples. Probably, low rickettsial DNA content is indicative of the fact that the last blood meal of the tick contained rickettsiae, rather than indicating true infection of the tick. Nevertheless, R. conorii israelensis was identified in two Rh. camicasi pools and R. aeschlimannii in one Rh. evertsi evertsi pool. Rickettsia conorii israelensis is the causative agent of Israeli spotted fever and occurs mainly in the Mediterranean countries [54]. Nevertheless, it has been occasionally detected in Africa, e.g., in Tunisia [58], Nigeria [59] and Kenya [60]. Studies proved that Rh. sanguineus s.l. acts as a vector of R. conorii israelensis, while the competency of Rh. camicasi as a vector is yet to be confirmed [61].
Of note, most of the ticks collected in the present study infest humans only occasionally [62,63]. Nonetheless, this does not rule out the risk of Rickettsia spp. transmission to humans. Currently, there are no published data on human Rickettsia exposure in Sudan or on the incidence of African tick-bite fever or other rickettsioses. Regarding livestock, high seroprevalences have been observed in sheep (59.3%) and cattle (64.4%) [64]. Considering these high seroprevalence rates and the reported MIRs in this study, investigations into the epidemiology of rickettsiosis in humans are required, concentrating on at-risk populations, especially rural communities with frequent contact with livestock.
Furthermore, relevant tick-borne pathogens for domestic animal health were detected in this study. MIRs were 0.58% for Theileria spp. and 0.12% for Babesia species. Serological and molecular evidence for circulation of these piroplasms among livestock has been reported in Sudan before [13,65]. Molecular characterization by sequencing of the 18S rDNA revealed that the investigated ticks carried T. annulata. In Sudan, bovine tropical theileriosis has been recognized as one of the main limitations that slow the development of the dairy industry [66]. A westward spread of T. annulata with its main vector, H. anatolicum, has occurred in Sudan, and the pathogen is now also present in North Kordofan, where it was believed to be absent until 2015 [49]. In the present study, we detected H. anatolicum in North Kordofan, but all T. annulata-positive ticks (one H. anatolicum and one Rh. evertsi evertsi pool) were from Kassala. Therefore, further studies are needed to assess the risk of T. annulata transmission to cattle in North Kordofan.
Overall, the high diversity of pathogenic piroplasms detected in the present study indicates that tick control is relevant for all livestock species in Sudan. Besides T. annulata, T. lestoquardi that leads to malignant ovine theileriosis, as well as T. equi and B. caballi, the etiological agents of equine piroplasmosis were also detected, in addition to the apathogenic species T. ovis and T. velifera [67]. Regarding equine piroplasmosis, T. equi was detected in H. anatolicum and B. caballi in A. variegatum. While H. anatolicum is a relevant vector for T. equi, the detection of B. caballi in A. variegatum may indicate that this tick had simply ingested infected blood, as Amblyomma ticks are not known to act as vectors of Babesia spp. [68].
In addition, B. pecorum was detected in two H. impeltatum pools. This finding suggests that this parasite is globally widespread, since it has been reported in wild animals in South Africa and Spain and in sheep in China [69,70,71]. For transmission of this parasite, H. anatolicum showed vector competency in China, whereas in Spain, H. lusitanicum was suggested to be the vector of B. pecorum [69,71]. It is unlikely that B. pecorum is of any relevance for domestic animal health, as experimentally infected non-immunosuppressed sheep and calves did not show any clinical signs [69,71].
5. Conclusions
The present study demonstrated a diverse tick fauna on livestock and other domestic animals in Sudan, with Hyalomma spp. predominating in North Kordofan and Rhipicephalus spp. in Kassala. In addition, the newly described species Rh. afranicus was detected. The high Rickettsia infection rates indicate a non-negligible risk for humans, especially in pastoral communities and rural areas. The presence of R. conorii israelensis in Sudan was documented for the first time. The detection of the highly pathogenic livestock piroplasms (T. annulata, T. lestoquardi, T. equi and B. caballi) is an indicator of the need for control programs to reduce the potential economic losses due to ticks and TBDs, as well as for further studies to provide a full picture of their epidemiology.
Author Contributions
Conceptualization, L.C.-D., Y.A.S., C.S. and G.D.; investigation, L.C.-D., A.S., M.H.I., M.I.-E.E.-E., A.Y.O., I.A.Y., M.A.A., A.O.B., S.E.-T.M.-N., S.S., and R.R.; formal analysis, D.K.B.; visualization, A.S., A.Y.O. and D.K.B.; writing—original draft preparation, A.S. and Y.A.S.; writing—review and editing, L.C.-D., D.K.B., C.S., G.D. All authors have read and agreed to the published version of the manuscript.
Funding
The project was funded in part by the institutional grant STAN 53-2012-11 of the Bundeswehr Institute of Microbiology. A.Y.O. is part of PANDORA-ID-NET Consortium (EDCTP Reg/Grant RIA2016E-1609) funded by the European and Developing Countries Clinical Trials Partnership (EDCTP2) programme, which is supported under Horizon 2020, the European Union’s Framework Programme for Research and Innovation. This publication was supported by Deutsche Forschungsgemeinschaft and University of Veterinary Medicine Hannover, Foundation within the funding programme Open Access Publishing.
Acknowledgments
The authors are thankful to all of colleagues in the study area who helped during sampling and to the Director General of the Ministry of Animal Resources and Fisheries (MARF), Khartoum, Sudan, for giving permission to send the samples abroad for laboratory analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Uilenberg, G. Veterinary significance of ticks and tick-borne diseases. In Tick Vector Biology; Springer: Berlin/Heidelberg, Germany, 1992; pp. 23–33. [Google Scholar]
- Obregón Alvarez, D.; Corona-González, B.; Rodríguez-Mallón, A.; Rodríguez Gonzalez, I.; Alfonso, P.; Noda Ramos, A.A.; Díaz-Sánchez, A.A.; González Navarrete, M.; Rodríguez Fernández, R.; Méndez Mellor, L.; et al. Ticks and tick-borne diseases in Cuba, half a century of scientific research. Pathogens 2020, 9, 616. [Google Scholar] [CrossRef]
- de la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef]
- Lew-Tabor, A.E.; Rodriguez Valle, M. A review of reverse vaccinology approaches for the development of vaccines against ticks and tick borne diseases. Ticks Tick Borne Dis. 2016, 7, 573–585. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Climate change, biodiversity, ticks and tick-borne diseases: The butterfly effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef]
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J. Vital signs: Trends in reported vectorborne disease cases—United States and Territories, 2004–2016. Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vicente, S.; Tagliafierro, T.; Coleman, J.L.; Benach, J.L.; Tokarz, R. Polymicrobial nature of tick-borne diseases. mBio 2019, 10, e02055-19. [Google Scholar] [CrossRef] [PubMed]
- Muhanguzi, D.; Byaruhanga, J.; Amanyire, W.; Ndekezi, C.; Ochwo, S.; Nkamwesiga, J.; Mwiine, F.N.; Tweyongyere, R.; Fourie, J.; Madder, M.; et al. Invasive cattle ticks in East Africa: Morphological and molecular confirmation of the presence of Rhipicephalus microplus in south-eastern Uganda. Parasites Vectors 2020, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Makenov, M.; Toure, A.; Korneev, M.; Sacko, N.; Porshakov, A.; Yakovlev, S.; Radyuk, E.; Zakharov, K.; Shipovalov, A.; Boumbaly, S.; et al. Rhipicephalus microplus and its vector-borne haemoparasites in Guinea: Further species expansion in West Africa. bioRxiv 2020. [Google Scholar] [CrossRef]
- Walker, A.R.; Bouattour, A.; Camicas, J.-L.; Estrada-Pena, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Endinburgh, UK, 2014. [Google Scholar]
- Lorusso, V.; Wijnveld, M.; Majekodunmi, A.O.; Dongkum, C.; Fajinmi, A.; Dogo, A.G.; Thrusfield, M.; Mugenyi, A.; Vaumourin, E.; Igweh, A.C.; et al. Tick-borne pathogens of zoonotic and veterinary importance in Nigerian cattle. Parasites Vectors 2016, 9, 217. [Google Scholar] [CrossRef]
- Raboloko, O.O.; Ramabu, S.S.; Guerrini, L.; Jori, F. Seroprevalence of selected tick borne pathogens and diversity and abundance of ixodid ticks (Acari: Ixodidae) at the wildlife-livestock interface in Northern Botswana. Front. Vet. Sci. 2020, 7, 187. [Google Scholar] [CrossRef]
- Shuaib, Y.A.; Osman, H.M.; Hussein, M.O.; Bakhiet, M.A.; Omer, R.A.; Al-Nahas, A.; Suliman, S.E.; Abdalla, M.A.; Ismail, A.A. Seroprevalence of Babesia bigemina antibodies in cattle in North Kordofan state, the Sudan. ARC J. Anim. Vet. Sci. 2015, 1, 1–11. [Google Scholar]
- Chitimia-Dobler, L.; Issa, M.H.; Ezalden, M.E.; Yagoub, I.A.; Abdalla, M.A.; Bakhiet, A.O.; Schaper, S.; Rieß, R.; Vollmar, P.; Grumbach, A.; et al. Crimean-Congo haemorrhagic fever virus in Hyalomma impeltatum ticks from North Kordofan, the Sudan. Int. J. Infect. Dis. 2019, 89, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Shuaib, Y.A.; Elhag, A.M.-A.W.; Brima, Y.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.-T.; Lemhöfer, G.; Bestehorn, M.; Poppert, S.; Schaper, S.; et al. Ixodid tick species and two tick-borne pathogens in three areas in the Sudan. Parasitol. Res. 2020, 119, 385–394. [Google Scholar] [CrossRef] [PubMed]
- MARF. Number of Animals in the Sudan; Ministry of Animal Resources and Fisheries (MARF), Statistics: Khartoum, Sudan, 2012.
- Hassan, S.; Salih, D. An overview of factors responsible for geographic distribution pattern of ixodid ticks in the Sudan. Sokoto J. Vet. Sci. 2013, 11, 1–9. [Google Scholar] [CrossRef][Green Version]
- Salih, D.A.; El Hussein, A.M.; Seitzer, U.; Ahmed, J.S. Epidemiological studies on tick-borne diseases of cattle in Central Equatoria State, Southern Sudan. Parasitol. Res. 2007, 101, 1035–1044. [Google Scholar] [CrossRef]
- Salim, B.; Bakheit, M.A.; Kamau, J.; Sugimoto, C. Current status of equine piroplasmosis in the Sudan. Infect. Genet. Evol. 2013, 16, 191–199. [Google Scholar] [CrossRef]
- Nakao, M.; Qiu, Y.; Salim, B.; Hassan, S.M.; Sugimoto, C. Molecular detection of Rickettsia africae in Amblyomma variegatum collected from Sudan. Vector Borne Zoonotic Dis. 2015, 15, 323–325. [Google Scholar] [CrossRef]
- Maina, A.N.; Jiang, J.; Omulo, S.A.; Cutler, S.J.; Ade, F.; Ogola, E.; Feikin, D.R.; Njenga, M.K.; Cleaveland, S.; Mpoke, S.; et al. High prevalence of Rickettsia africae variants in Amblyomma variegatum ticks from domestic mammals in rural western Kenya: Implications for human health. Vector Borne Zoonotic Dis. 2014, 14, 693–702. [Google Scholar] [CrossRef]
- Apanaskevich, D.; Horak, I. The genus Hyalomma Koch, 1844: II Taxonomic status of H. (Euhyalomma) anatolicum Koch, 1844 and H. (E.) excavatum Koch, 1844 (Acari: Ixodidae) with re-description of all stages. Acarina 2005, 13, 181–197. [Google Scholar]
- Apanaskevich, D.A.; Horak, I.G. The genus Hyalomma Koch, 1844: V Re-evaluation of the taxonomic rank of taxa comprising the H. (Euhyalomma) marginatum Koch complex of species (Acari: Ixodidae) with redescription of all parasitic stages and notes on biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Horak, I.G. The genus Hyalomma Koch, 1844. IX. Redescription of all parasitic stages of H. (Euhyalomma) impeltatum Schulze & Schlottke, 1930 and H. (E.) somalicum Tonelli Rondelli, 1935 (Acari: Ixodidae). Syst. Parasitol. 2009, 73, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The genus Hyalomma: VII. Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef]
- Voltzit, O.; Keirans, J. A review of African Amblyomma species (Acari, Ixodida, Ixodidae). Acarina 2003, 11, 135–214. [Google Scholar]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar] [CrossRef]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Bakkes, D.K.; Chitimia-Dobler, L.; Matloa, D.; Oosthuysen, M.; Mumcuoglu, K.Y.; Mans, B.J.; Matthee, C.A. Integrative taxonomy and species delimitation of Rhipicephalus turanicus (Acari: Ixodida: Ixodidae). Int. J. Parasitol. 2020, 50, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Black, W.C.; Piesman, J. Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Langguth, J.; Pfeffer, M.; Kattner, S.; Küpper, T.; Friese, D.; Dobler, G.; Guglielmone, A.A.; Nava, S. Genetic analysis of Rhipicephalus sanguineus sensu lato ticks parasites of dogs in Africa north of the Sahara based on mitochondrial DNA sequences. Vet. Parasitol. 2017, 239, 1–6. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Latrofa, M.; Annoscia, G.; Giannelli, A.; Parisi, A.; Otranto, D. Morphological and genetic diversity of Rhipicephalus sanguineus sensu lato from the New and Old Worlds. Parasites Vectors 2013, 6, 213. [Google Scholar] [CrossRef]
- Nava, S.; Beati, L.; Venzal, J.M.; Labruna, M.B.; Szabó, M.P.J.; Petney, T.; Saracho-Bottero, M.N.; Tarragona, E.L.; Dantas-Torres, F.; Silva, M.M.S.; et al. Rhipicephalus sanguineus (Latreille, 1806): Neotype designation, morphological re-description of all parasitic stages and molecular characterization. Ticks Tick. Borne. Dis. 2018, 9, 1573–1585. [Google Scholar] [CrossRef]
- Sands, A.F.; Apanaskevich, D.A.; Matthee, S.; Horak, I.G.; Harrison, A.; Karim, S.; Mohammad, M.K.; Mumcuoglu, K.Y.; Rajakaruna, R.S.; Santos-Silva, M.M.; et al. Effects of tectonics and large scale climatic changes on the evolutionary history of Hyalomma ticks. Mol. Phylogenet. Evol. 2017, 114, 153–165. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, B.; Chamberlain, J.; Logue, C.H.; Cook, N.; Bruce, C.; Dowall, S.D.; Hewson, R. Development of a real-time RT-PCR assay for the detection of Crimean-Congo hemorrhagic fever virus. Vector Borne Zoonotic Dis. 2012, 12, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, R.; Essbauer, S.; Dobler, G. Diagnostics of tick-borne rickettsioses in Germany: A modern concept for a neglected disease. Int. J. Med. Microbiol. 2008, 298, 368–374. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Riess, R.; Kahl, O.; Wolfel, S.; Dobler, G.; Nava, S.; Estrada-Pena, A. Ixodes inopinatus—Occurring also outside the Mediterranean region. Ticks Tick Borne Dis. 2018, 9, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, L.; Chelo, I.M.; Bacellar, F.; Zé-Zé, L. Rickettsiae phylogeny: A multigenic approach. Microbiology 2007, 153, 160–168. [Google Scholar] [CrossRef]
- Weinert, L.A.; Werren, J.H.; Aebi, A.; Stone, G.N.; Jiggins, F.M. Evolution and diversity of Rickettsia bacteria. BMC Biol. 2009, 7, 6. [Google Scholar] [CrossRef]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- Springer, A.; Höltershinken, M.; Lienhart, F.; Ermel, S.; Rehage, J.; Hülskötter, K.; Lehmbecker, A.; Wohlsein, P.; Barutzki, D.; Gietl, C.; et al. Emergence and epidemiology of bovine babesiosis due to Babesia divergens on a northern German beef production farm. Front. Vet. Sci. 2020, 7, 649. [Google Scholar] [CrossRef]
- Adehan, S.B.; Adakal, H.; Gbinwoua, D.; Yokossi, D.; Zoungrana, S.; Toe, P.; Ouedraogo, M.; Gbaguidi, A.M.; Adoligbé, C.; Fandohan, A.B.; et al. West African cattle farmers’ perception of tick-borne diseases. EcoHealth 2018, 15, 437–449. [Google Scholar] [CrossRef]
- Jongejan, F.; Zivkovic, D.; Pegram, R.G.; Tatchell, R.J.; Fison, T.; Latif, A.A.; Paine, G. Ticks (Acari:Ixodidae) of the Blue and White Nile ecosystems in the Sudan with particular reference to the Rhipicephalus sanguineus group. Exp. Appl. Acarol. 1987, 3, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Karrar, G.; Kaiser, M.N.; Hoogstraal, H. Ecology and host-relationships of ticks (Ixodoidea) infesting domestic animals in Kassala Province, Sudan, with special reference to Amblyomma lepidum Dönitz. Bull. Entomol. Res. 1963, 54, 509–522. [Google Scholar] [CrossRef]
- Salih, D.A.; Hassan, S.M.; El Hussein, A.M.; Jongejan, F. Preliminary survey of ticks (Acari: Ixodidae) on cattle in northern Sudan. Onderstepoort J. Vet. Res. 2004, 71, 319–326. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mohammed-Ahmed, G.M.; Hassan, S.M.; El Hussein, A.M.; Salih, D.A. Molecular, serological and parasitological survey of Theileria annulata in North Kordofan State, Sudan. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.M.; El-Hussein, A.M.; El-Khider, A.O. Some observations on ticks (Acari: Ixodidae) infesting sheep in River Nile Province of Northern Sudan. Onderstepoort J. Vet. Res. 2005, 72, 239–243. [Google Scholar] [CrossRef][Green Version]
- Elghali, A.; Hassan, S.M. Ticks (Acari: Ixodidae) infesting camels (Camelus dromedarius) in Northern Sudan. Onderstepoort J. Vet. Res. 2009, 76, 177–185. [Google Scholar] [CrossRef]
- Marcelino, I.; Holzmuller, P.; Stachurski, F.; Rodrigues, V.; Vachiéry, N. Ehrlichia ruminantium: The causal agent of heartwater. In Rickettsiales: Biology, Molecular Biology, Epidemiology, and Vaccine Development; Thomas, S., Ed.; Springer: Cham, Switzerland, 2016; pp. 241–280. [Google Scholar] [CrossRef]
- Balinandi, S.; Chitimia-Dobler, L.; Grandi, G.; Nakayiki, T.; Kabasa, W.; Bbira, J.; Lutwama, J.J.; Bakkes, D.K.; Malmberg, M.; Mugisha, L. Morphological and molecular identification of ixodid tick species (Acari: Ixodidae) infesting cattle in Uganda. Parasitol. Res. 2020, 119, 2411–2420. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Kumsa, B.; Socolovschi, C.; Raoult, D.; Parola, P. Spotted fever group rickettsiae in ixodid ticks in Oromia, Ethiopia. Ticks Tick Borne Dis. 2015, 6, 8–15. [Google Scholar] [CrossRef]
- Morita, C.; El Hussein, A.R.; Matsuda, E.; Abdel Gabbar, K.M.; Muramatsu, Y.; Abdel Rahman, M.B.; Eleragi, A.M.; Hassan, S.M.; Chitambo, A.M.; Ueno, H. Spotted fever group rickettsiae from ticks captured in Sudan. Jpn. J. Infect. Dis. 2004, 57, 107–109. [Google Scholar]
- Mura, A.; Socolovschi, C.; Ginesta, J.; Lafrance, B.; Magnan, S.; Rolain, J.-M.; Davoust, B.; Raoult, D.; Parola, P. Molecular detection of spotted fever group rickettsiae in ticks from Ethiopia and Chad. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Znazen, A.; Khrouf, F.; Elleuch, N.; Lahiani, D.; Marrekchi, C.; M’Ghirbi, Y.; Ben Jemaa, M.; Bouattour, A.; Hammami, A. Multispacer typing of Rickettsia isolates from humans and ticks in Tunisia revealing new genotypes. Parasites Vectors 2013, 6, 367. [Google Scholar] [CrossRef] [PubMed]
- Kamani, J.; Baneth, G.; Mumcuoglu, K.Y.; Waziri, N.E.; Eyal, O.; Guthmann, Y.; Harrus, S. Molecular detection and characterization of tick-borne pathogens in dogs and ticks from Nigeria. PLoS Negl. Trop. Dis. 2013, 7, e2108. [Google Scholar] [CrossRef] [PubMed]
- Mutai, B.K.; Wainaina, J.M.; Magiri, C.G.; Nganga, J.K.; Ithondeka, P.M.; Njagi, O.N.; Jiang, J.; Richards, A.L.; Waitumbi, J.N. Zoonotic surveillance for rickettsiae in domestic animals in Kenya. Vector Borne Zoonotic Dis. 2013, 13, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Zemtsova, G.; Killmaster, L.F.; Mumcuoglu, K.Y.; Levin, M.L. Co-feeding as a route for transmission of Rickettsia conorii israelensis between Rhipicephalus sanguineus ticks. Exp. Appl. Acarol. 2010, 52, 383–392. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Jongejan, F. Ticks feeding on humans: A review of records on human-biting Ixodoidea with special reference to pathogen transmission. Exp. Appl. Acarol. 1999, 23, 685–715. [Google Scholar] [CrossRef]
- Horak, I.G.; Fourie, L.J.; Heyne, H.; Walker, J.B.; Needham, G.R. Ixodid ticks feeding on humans in South Africa: With notes on preferred hosts, geographic distribution, seasonal occurrence and transmission of pathogens. Exp. Appl. Acarol. 2002, 27, 113–136. [Google Scholar] [CrossRef]
- Eisawi, N.M.; Hassan, D.A.; Hussien, M.O.; Musa, A.B.; El Hussein, A.R.M. Seroprevalence of spotted fever group (SFG) rickettsiae infection in domestic ruminants in Khartoum State, Sudan. Vet. Med. Sci. 2017, 3, 91–98. [Google Scholar] [CrossRef]
- Taha, K.M.; Salih, D.A.; Ali, A.M.; Omer, R.A.; El Hussein, A.M. Naturally occurring infections of cattle with Theileria lestoquardi and sheep with Theileria annulata in the Sudan. Vet. Parasitol. 2013, 191, 143–145. [Google Scholar] [CrossRef]
- El Hussein, A.M.; Hassan, S.M.; Salih, D.A. Current situation of tropical theileriosis in the Sudan. Parasitol. Res. 2012, 111, 503–508. [Google Scholar] [CrossRef]
- Mans, B.J.; Pienaar, R.; Latif, A.A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Estrada-Peña, A.; Zintl, A. Vectors of Babesiosis. Ann. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.; Ma, M.; Moreau, E.; Liu, J.; Lu, B.; Bai, Q.; Luo, J.; Jorgensen, W.; Chauvin, A.; Yin, H. A new ovine Babesia species transmitted by Hyalomma anatolicum anatolicum. Exp. Parasitol. 2009, 122, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Oosthuizen, M.C.; Allsopp, B.A.; Troskie, M.; Collins, N.E.; Penzhorn, B.L. Identification of novel Babesia and Theileria species in South African giraffe (Giraffa camelopardalis, Linnaeus, 1758) and roan antelope (Hippotragus equinus, Desmarest 1804). Vet. Parasitol. 2009, 163, 39–46. [Google Scholar] [CrossRef]
- Jouglin, M.; Fernández-de-Mera, I.G.; de la Cotte, N.; Ruiz-Fons, F.; Gortázar, C.; Moreau, E.; Bastian, S.; de la Fuente, J.; Malandrin, L. Isolation and characterization of Babesia pecorum sp. nov. from farmed red deer (Cervus elaphus). Vet. Res. 2014, 45, 78. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).