
Rotavirus Reverse Genetics Systems and Oral Vaccine Delivery Vectors for Mucosal Vaccination

 ,
,  ,
,
Abstract
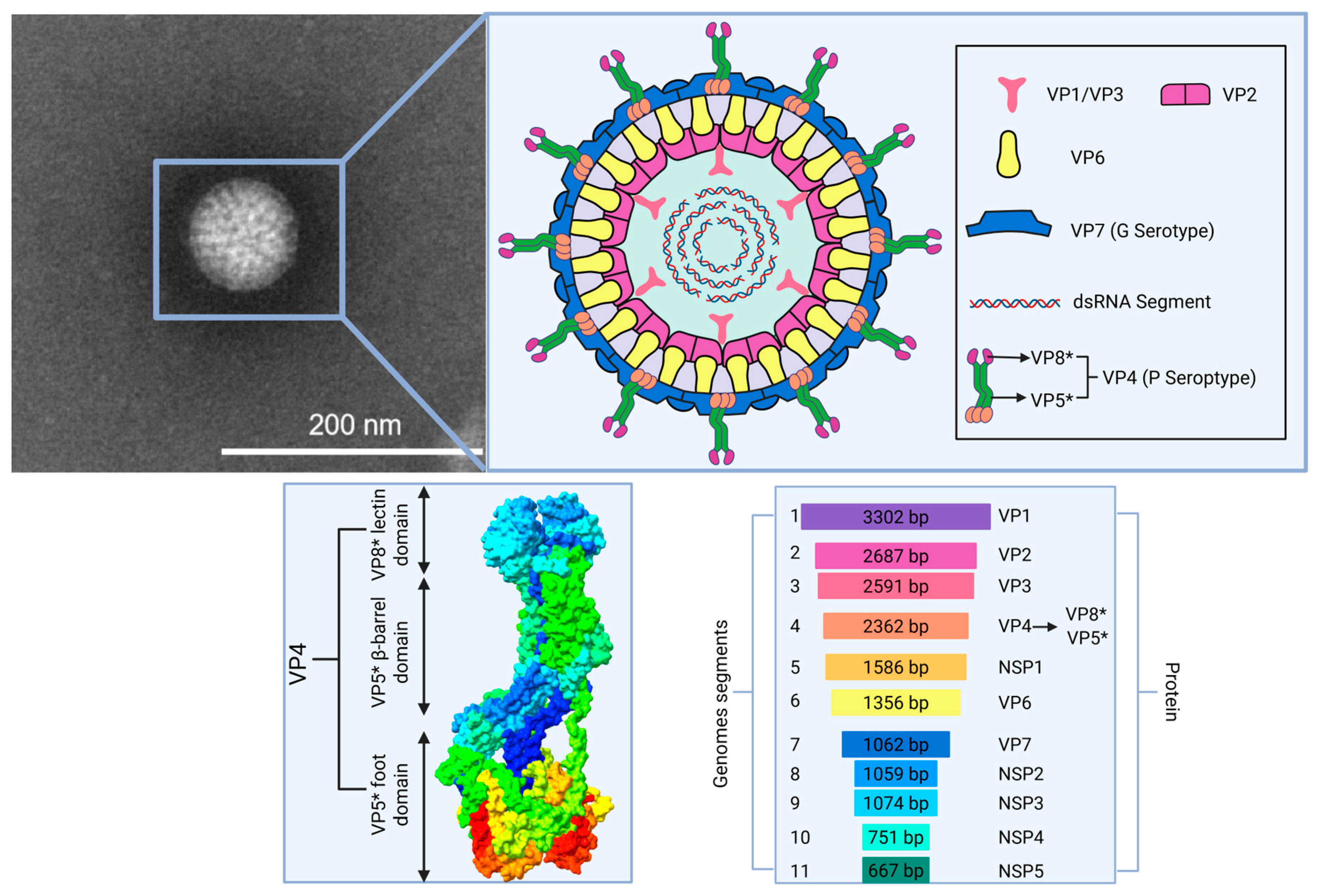
1. Introduction
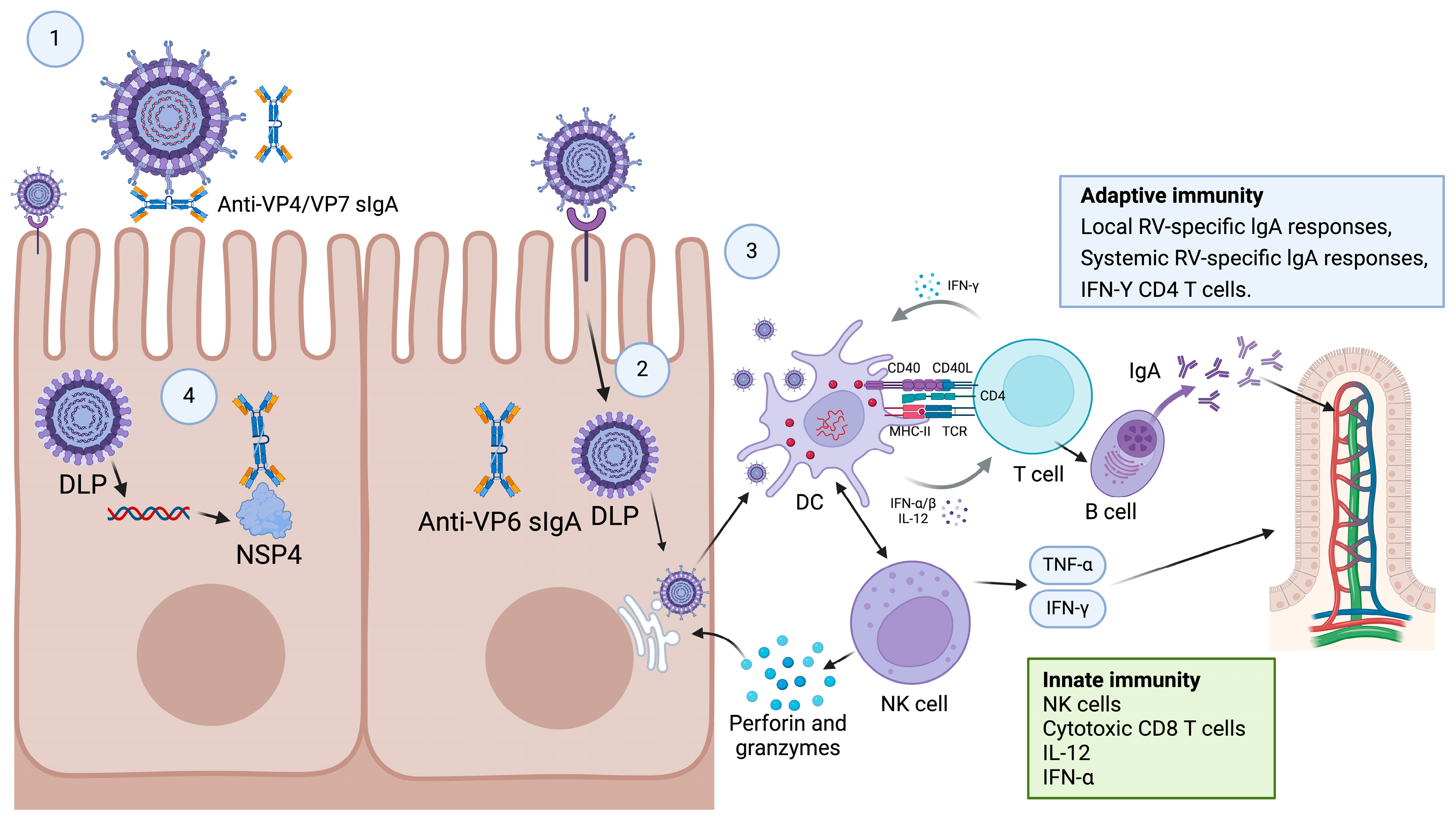
2. Colonization of the Intestine and Stimulation of Intestinal Immunity by RVs
3. Live-Attenuated RV as a Gene Delivery Vector Involving Natural Gene Reassortment
4. Reverse Genetic System of RV
4.1. RV Reverse Genetic System Involving Helper Viruses and Pressure Screening
4.2. Entirely Plasmid-Based Reverse Genetics System for RVs
5. Application of RV as a Vector
6. Challenges of RV as a Transduction Vector
7. Advantages of RVs as Vectors
8. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef]
- Seki, N.; Kimizuka, T.; Gondo, M.; Yamaguchi, G.; Sugiura, Y.; Akiyama, M.; Yakabe, K.; Uchiyama, J.; Higashi, S.; Haneda, T.; et al. (D)-Tryptophan suppresses enteric pathogen and pathobionts and prevents colitis by modulating microbial tryptophan metabolism. iScience 2022, 25, 104838. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef]
- Garrett, W.S.; Lord, G.M.; Punit, S.; Lugo-Villarino, G.; Mazmanian, S.K.; Ito, S.; Glickman, J.N.; Glimcher, L.H. Communicable ulcerative colitis induced by T-bet deficiency in the innate immune system. Cell 2007, 131, 33–45. [Google Scholar] [CrossRef]
- Yamashiro, R.; Sakudo, A.; Nagatsu, M. Efficient recovery and enrichment of infectious rotavirus using separation with antibody-integrated graphite-encapsulated magnetic nanobeads produced by argon/ammonia gas plasma technology. Int. J. Nanomed. 2019, 14, 1865–1876. [Google Scholar] [CrossRef]
- WHO Diarrhoeal Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/diarrhoeal-disease (accessed on 7 March 2024).
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef]
- Elliott, E.J. Acute gastroenteritis in children. BMJ (Clin. Res. Ed.) 2007, 334, 35–40. [Google Scholar] [CrossRef]
- Zhang, S.X.; Yang, C.L.; Gu, W.P.; Ai, L.; Serrano, E.; Yang, P.; Zhou, X.; Li, S.Z.; Lv, S.; Dang, Z.S.; et al. Case-control study of diarrheal disease etiology in individuals over 5 years in southwest China. Gut Pathog. 2016, 8, 58. [Google Scholar] [CrossRef]
- Cossart, P.; Sansonetti, P.J. Bacterial invasion: The paradigms of enteroinvasive pathogens. Science 2004, 304, 242–248. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Shi, C.Z.; Chen, H.Q.; Liang, Y.; Xia, Y.; Yang, Y.Z.; Yang, J.; Zhang, J.D.; Wang, S.H.; Liu, J.; Qin, H.L. Combined probiotic bacteria promotes intestinal epithelial barrier function in interleukin-10-gene-deficient mice. World J. Gastroenterol. 2014, 20, 4636–4647. [Google Scholar] [CrossRef]
- Yinda, C.K.; Vanhulle, E.; Conceição-Neto, N.; Beller, L.; Deboutte, W.; Shi, C.; Ghogomu, S.M.; Maes, P.; Van Ranst, M.; Matthijnssens, J. Gut Virome Analysis of Cameroonians Reveals High Diversity of Enteric Viruses, Including Potential Interspecies Transmitted Viruses. mSphere 2019, 4, e00585-18. [Google Scholar] [CrossRef]
- Niu, X.; Kong, F.; Xu, J.; Liu, M.; Wang, Q. Mutations in Porcine Epidemic Diarrhea Virus nsp1 Cause Increased Viral Sensitivity to Host Interferon Responses and Attenuation In Vivo. J. Virol. 2022, 96, e00469-22. [Google Scholar] [CrossRef]
- Xu, Y.; Yuen, P.W.; Lam, J.K. Intranasal DNA Vaccine for Protection against Respiratory Infectious Diseases: The Delivery Perspectives. Pharmaceutics 2014, 6, 378–415. [Google Scholar] [CrossRef]
- Song, D.S.; Oh, J.S.; Kang, B.K.; Yang, J.S.; Moon, H.J.; Yoo, H.S.; Jang, Y.S.; Park, B.K. Oral efficacy of Vero cell attenuated porcine epidemic diarrhea virus DR13 strain. Res. Vet. Sci. 2007, 82, 134–140. [Google Scholar] [CrossRef]
- Pabst, O. New concepts in the generation and functions of IgA. Nat. Rev. Immunol. 2012, 12, 821–832. [Google Scholar] [CrossRef]
- Lim, C.H.; Voedisch, S.; Wahl, B.; Rouf, S.F.; Geffers, R.; Rhen, M.; Pabst, O. Independent bottlenecks characterize colonization of systemic compartments and gut lymphoid tissue by Salmonella. PLoS Pathog. 2014, 10, e1004270. [Google Scholar] [CrossRef]
- Sano, K.; Bhavsar, D.; Singh, G.; Floda, D.; Srivastava, K.; Gleason, C.; Carreño, J.M.; Simon, V.; Krammer, F. SARS-CoV-2 vaccination induces mucosal antibody responses in previously infected individuals. Nat. Commun. 2022, 13, 5135. [Google Scholar] [CrossRef]
- Wahid, R.; Fresnay, S.; Levine, M.M.; Sztein, M.B. Cross-reactive multifunctional CD4+ T cell responses against Salmonella enterica serovars Typhi, Paratyphi A and Paratyphi B in humans following immunization with live oral typhoid vaccine Ty21a. Clin. Immunol. 2016, 173, 87–95. [Google Scholar] [CrossRef]
- Mohan, T.; Kim, J.; Berman, Z.; Wang, S.; Compans, R.W.; Wang, B.Z. Co-delivery of GPI-anchored CCL28 and influenza HA in chimeric virus-like particles induces cross-protective immunity against H3N2 viruses. J. Control. Release 2016, 233, 208–219. [Google Scholar] [CrossRef]
- Huang, X.N.; Ma, Y.Y.; Wang, Y.X.; Niu, C.; Liu, Z.M.; Yao, X.; Jiang, X.X.; Pan, R.H.; Jia, S.; Li, D.D.; et al. Oral Probiotic Vaccine Expressing Koi Herpesvirus (KHV) ORF81 Protein Delivered by Chitosan-Alginate Capsules Is a Promising Strategy for Mass Oral Vaccination of Carps against KHV Infection. J. Virol. 2021, 95, e00415-21. [Google Scholar] [CrossRef]
- Romero-Maraccini, O.C.; Sadik, N.J.; Rosado-Lausell, S.L.; Pugh, C.R.; Niu, X.Z.; Croué, J.P.; Nguyen, T.H. Sunlight-induced inactivation of human Wa and porcine OSU rotaviruses in the presence of exogenous photosensitizers. Environ. Sci. Technol. 2013, 47, 11004–11012. [Google Scholar] [CrossRef]
- Komoto, S.; Sasaki, J.; Taniguchi, K. Reverse genetics system for introduction of site-specific mutations into the double-stranded RNA genome of infectious rotavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 4646–4651. [Google Scholar] [CrossRef]
- Kanai, Y.; Komoto, S.; Kawagishi, T.; Nouda, R.; Nagasawa, N.; Onishi, M.; Matsuura, Y.; Taniguchi, K.; Kobayashi, T. Entirely plasmid-based reverse genetics system for rotaviruses. Proc. Natl. Acad. Sci. USA 2017, 114, 2349–2354. [Google Scholar] [CrossRef]
- Agbemabiese, C.A.; Philip, A.A.; Patton, J.T. Recovery of Recombinant Rotaviruses by Reverse Genetics. Methods Mol. Biol. 2023, 2733, 249–263. [Google Scholar] [CrossRef]
- Trask, S.D.; Taraporewala, Z.F.; Boehme, K.W.; Dermody, T.S.; Patton, J.T. Dual selection mechanisms drive efficient single-gene reverse genetics for rotavirus. Proc. Natl. Acad. Sci. USA 2010, 107, 18652–18657. [Google Scholar] [CrossRef]
- Troupin, C.; Dehée, A.; Schnuriger, A.; Vende, P.; Poncet, D.; Garbarg-Chenon, A. Rearranged genomic RNA segments offer a new approach to the reverse genetics of rotaviruses. J. Virol. 2010, 84, 6711–6719. [Google Scholar] [CrossRef]
- Johne, R.; Reetz, J.; Kaufer, B.B.; Trojnar, E. Generation of an Avian-Mammalian Rotavirus Reassortant by Using a Helper Virus-Dependent Reverse Genetics System. J. Virol. 2016, 90, 1439–1443. [Google Scholar] [CrossRef]
- Komoto, S.; Fukuda, S.; Ide, T.; Ito, N.; Sugiyama, M.; Yoshikawa, T.; Murata, T.; Taniguchi, K. Generation of Recombinant Rotaviruses Expressing Fluorescent Proteins by Using an Optimized Reverse Genetics System. J. Virol. 2018, 92, e00588-18. [Google Scholar] [CrossRef]
- Sánchez-Tacuba, L.; Feng, N.; Meade, N.J.; Mellits, K.H.; Jaïs, P.H.; Yasukawa, L.L.; Resch, T.K.; Jiang, B.; López, S.; Ding, S.; et al. An Optimized Reverse Genetics System Suitable for Efficient Recovery of Simian, Human, and Murine-Like Rotaviruses. J. Virol. 2020, 94, e01294-20. [Google Scholar] [CrossRef]
- Kanai, Y.; Onishi, M.; Kawagishi, T.; Pannacha, P.; Nurdin, J.A.; Nouda, R.; Yamasaki, M.; Lusiany, T.; Khamrin, P.; Okitsu, S.; et al. Reverse Genetics Approach for Developing Rotavirus Vaccine Candidates Carrying VP4 and VP7 Genes Cloned from Clinical Isolates of Human Rotavirus. J. Virol. 2020, 95, e01374-20. [Google Scholar] [CrossRef]
- Philip, A.A.; Patton, J.T. Rotavirus as an Expression Platform of Domains of the SARS-CoV-2 Spike Protein. Vaccines 2021, 9, 449. [Google Scholar] [CrossRef]
- Diebold, O.; Gonzalez, V.; Venditti, L.; Sharp, C.; Blake, R.A.; Tan, W.S.; Stevens, J.; Caddy, S.; Digard, P.; Borodavka, A.; et al. Using Species a Rotavirus Reverse Genetics to Engineer Chimeric Viruses Expressing SARS-CoV-2 Spike Epitopes. J. Virol. 2022, 96, e00488-22. [Google Scholar] [CrossRef]
- Kawagishi, T.; Sánchez-Tacuba, L.; Feng, N.; Costantini, V.P.; Tan, M.; Jiang, X.; Green, K.Y.; Vinjé, J.; Ding, S.; Greenberg, H.B. Mucosal and systemic neutralizing antibodies to norovirus induced in infant mice orally inoculated with recombinant rotaviruses. Proc. Natl. Acad. Sci. USA 2023, 120, e2214421120. [Google Scholar] [CrossRef]
- Qin, S.K.; Li, K.H.; Liu, B.J.; Cao, C.; Yu, D.B.; Jiang, Z.G.; Wang, J.; Han, Y.X.; Wang, F.; Qi, Y.L.; et al. Efficient and robust reverse genetics system for bovine rotavirus generation and its application for antiviral screening. Virol. Sin. 2024, 39, 917–928. [Google Scholar] [CrossRef]
- Kawamura, Y.; Komoto, S.; Fukuda, S.; Kugita, M.; Tang, S.; Patel, A.; Pieknik, J.R.; Nagao, S.; Taniguchi, K.; Krause, P.R.; et al. Development of recombinant rotavirus carrying herpes simplex virus 2 glycoprotein D gene based on reverse genetics technology. Microbiol. Immunol. 2024, 68, 56–64. [Google Scholar] [CrossRef]
- Oishi, T.; Hasegawa, S.; Nakano, T.; Sudo, S.; Kuwajima, H.; Tokuriki, S.; Tamura, T. Changes in vaccine coverage and incidence of acute gastroenteritis and severe rotavirus gastroenteritis in children <5 years in Shibata City, Niigata Prefecture, Japan. Hum. Vaccines Immunother. 2024, 20, 2322202. [Google Scholar] [CrossRef]
- Elkady, G.; Chen, Y.; Hu, C.; Chen, J.; Chen, X.; Guo, A. MicroRNA Profile of MA-104 Cell Line Associated with the Pathogenesis of Bovine Rotavirus Strain Circulated in Chinese Calves. Front. Microbiol. 2022, 13, 854348. [Google Scholar] [CrossRef]
- Qiu, Y.; Yang, J.; Wang, W.; Zhao, W.; Peng, F.; Xiang, Y.; Chen, G.; Chen, T.; Chai, C.; Zheng, S.; et al. HMGB1-promoted and TLR2/4-dependent NK cell maturation and activation take part in rotavirus-induced murine biliary atresia. PLoS Pathog. 2014, 10, e1004011. [Google Scholar] [CrossRef]
- Pesavento, J.B.; Crawford, S.E.; Roberts, E.; Estes, M.K.; Prasad, B.V. pH-induced conformational change of the rotavirus VP4 spike: Implications for cell entry and antibody neutralization. J. Virol. 2005, 79, 8572–8580. [Google Scholar] [CrossRef]
- Araud, E.; DiCaprio, E.; Yang, Z.; Li, X.; Lou, F.; Hughes, J.H.; Chen, H.; Li, J. High-Pressure Inactivation of Rotaviruses: Role of Treatment Temperature and Strain Diversity in Virus Inactivation. Appl. Environ. Microbiol. 2015, 81, 6669–6678. [Google Scholar] [CrossRef]
- Sánchez-San Martín, C.; López, T.; Arias, C.F.; López, S. Characterization of rotavirus cell entry. J. Virol. 2004, 78, 2310–2318. [Google Scholar] [CrossRef]
- Gonzalez-Hernandez, M.B.; Liu, T.; Payne, H.C.; Stencel-Baerenwald, J.E.; Ikizler, M.; Yagita, H.; Dermody, T.S.; Williams, I.R.; Wobus, C.E. Efficient norovirus and reovirus replication in the mouse intestine requires microfold (M) cells. J. Virol. 2014, 88, 6934–6943. [Google Scholar] [CrossRef]
- Santiana, M.; Ghosh, S.; Ho, B.A.; Rajasekaran, V.; Du, W.L.; Mutsafi, Y.; De Jésus-Diaz, D.A.; Sosnovtsev, S.V.; Levenson, E.A.; Parra, G.I.; et al. Vesicle-Cloaked Virus Clusters Are Optimal Units for Inter-Organismal Viral Transmission. Cell Host Microbe 2018, 24, 208–220.e8. [Google Scholar] [CrossRef]
- Azevedo, M.S.; Yuan, L.; Jeong, K.I.; Gonzalez, A.; Nguyen, T.V.; Pouly, S.; Gochnauer, M.; Zhang, W.; Azevedo, A.; Saif, L.J. Viremia and nasal and rectal shedding of rotavirus in gnotobiotic pigs inoculated with Wa human rotavirus. J. Virol. 2005, 79, 5428–5436. [Google Scholar] [CrossRef]
- Nyblade, C.; Zhou, P.; Frazier, M.; Frazier, A.; Hensley, C.; Fantasia-Davis, A.; Shahrudin, S.; Hoffer, M.; Agbemabiese, C.A.; LaRue, L.; et al. Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs. Viruses 2023, 15, 1864. [Google Scholar] [CrossRef]
- Ghosh, S.; Kumar, M.; Santiana, M.; Mishra, A.; Zhang, M.; Labayo, H.; Chibly, A.M.; Nakamura, H.; Tanaka, T.; Henderson, W.; et al. Enteric viruses replicate in salivary glands and infect through saliva. Nature 2022, 607, 345–350. [Google Scholar] [CrossRef]
- Miao, Q.; Pan, Y.; Gong, L.; Guo, L.; Wu, L.; Jing, Z.; Zhang, G.; Tian, J.; Feng, L. Full genome characterization of a human-porcine reassortment G12P[7] rotavirus and its pathogenicity in piglets. Transbound. Emerg. Dis. 2022, 69, 3506–3517. [Google Scholar] [CrossRef]
- Kim, H.H.; Park, J.G.; Matthijnssens, J.; Kim, H.J.; Kwon, H.J.; Son, K.Y.; Ryu, E.H.; Kim, D.S.; Lee, W.S.; Kang, M.I.; et al. Pathogenicity of porcine G9P[23] and G9P[7] rotaviruses in piglets. Vet. Microbiol. 2013, 166, 123–137. [Google Scholar] [CrossRef]
- Crawford, S.E.; Patel, D.G.; Cheng, E.; Berkova, Z.; Hyser, J.M.; Ciarlet, M.; Finegold, M.J.; Conner, M.E.; Estes, M.K. Rotavirus viremia and extraintestinal viral infection in the neonatal rat model. J. Virol. 2006, 80, 4820–4832. [Google Scholar] [CrossRef]
- Amimo, J.O.; Raev, S.A.; Chepngeno, J.; Mainga, A.O.; Guo, Y.S.; Saif, L.; Vlasova, A.N. Rotavirus Interactions with Host Intestinal Epithelial Cells. Front. Immunol. 2021, 12, 793841. [Google Scholar] [CrossRef]
- Criglar, J.M.; Anish, R.; Hu, L.; Crawford, S.E.; Sankaran, B.; Prasad, B.V.V.; Estes, M.K. Phosphorylation cascade regulates the formation and maturation of rotaviral replication factories. Proc. Natl. Acad. Sci. USA 2018, 115, E12015–E12023. [Google Scholar] [CrossRef]
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus infection. Nat. Rev. Dis. Primers 2017, 3, 17083. [Google Scholar] [CrossRef]
- Velázquez, F.R.; Matson, D.O.; Calva, J.J.; Guerrero, L.; Morrow, A.L.; Carter-Campbell, S.; Glass, R.I.; Estes, M.K.; Pickering, L.K.; Ruiz-Palacios, G.M. Rotavirus infection in infants as protection against subsequent infections. N. Engl. J. Med. 1996, 335, 1022–1028. [Google Scholar] [CrossRef]
- Hou, G.; Son, J.; Gomez Castro, M.F.; Kawagishi, T.; Ren, X.; Roth, A.N.; Antia, A.; Zeng, Q.; DeVeaux, A.L.; Feng, N.; et al. Innate immune sensing of rotavirus by intestinal epithelial cells leads to diarrhea. Cell Host Microbe 2025, 33, 408–419.E8. [Google Scholar] [CrossRef]
- Lockhart, A.; Mucida, D.; Parsa, R. Immunity to enteric viruses. Immunity 2022, 55, 800–818. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Amimo, J.O.; Saif, L.J. Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies. Viruses Basel 2017, 9, 48. [Google Scholar] [CrossRef]
- Desselberger, U.; Huppertz, H.I. Immune responses to rotavirus infection and vaccination and associated correlates of protection. J. Infect. Dis. 2011, 203, 188–195. [Google Scholar] [CrossRef]
- Ramig, R.F. Genetics of the rotaviruses. Annu. Rev. Microbiol. 1997, 51, 225–255. [Google Scholar] [CrossRef]
- Clark, H.F.; Offit, P.A.; Ellis, R.W.; Eiden, J.J.; Krah, D.; Shaw, A.R.; Pichichero, M.; Treanor, J.J.; Borian, F.E.; Bell, L.M.; et al. The development of multivalent bovine rotavirus (strain WC3) reassortant vaccine for infants. J. Infect. Dis. 1996, 174 (Suppl. 1), S73–S80. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Zhang, H.; Fang, H. Public health impact and cost-effectiveness of rotavirus vaccination in China: Comparison between private market provision and national immunization programs. Hum. Vaccines Immunother. 2022, 18, 2090162. [Google Scholar] [CrossRef]
- Ghonaim, A.H.; Rouby, S.R.; Nageeb, W.M.; Elgendy, A.A.; Xu, R.; Jiang, C.; Ghonaim, N.H.; He, Q.; Li, W. Insights into recent advancements in human and animal rotavirus vaccines: Exploring new frontiers. Virol. Sin. 2025, 40, 1–14. [Google Scholar] [CrossRef]
- Wang, J.; Wen, X.; Ran, X. Advances in the development of human rotavirus vaccine. Chin. J. Zoonosess 2021, 37, 278–284. [Google Scholar]
- Chang, J.T.; Li, X.; Liu, H.J.; Yu, L. Ovine rotavirus strain LLR-85-based bovine rotavirus candidate vaccines: Construction, characterization and immunogenicity evaluation. Vet. Microbiol. 2010, 146, 35–43. [Google Scholar] [CrossRef]
- Ávila-Pérez, G.; Nogales, A.; Martín, V.; Almazán, F.; Martínez-Sobrido, L. Reverse Genetic Approaches for the Generation of Recombinant Zika Virus. Viruses 2018, 10, 597. [Google Scholar] [CrossRef]
- Ishii, K.; Ueda, Y.; Matsuo, K.; Matsuura, Y.; Kitamura, T.; Kato, K.; Izumi, Y.; Someya, K.; Ohsu, T.; Honda, M.; et al. Structural analysis of vaccinia virus DIs strain: Application as a new replication-deficient viral vector. Virology 2002, 302, 433–444. [Google Scholar] [CrossRef]
- Komoto, S.; Kugita, M.; Sasaki, J.; Taniguchi, K. Generation of recombinant rotavirus with an antigenic mosaic of cross-reactive neutralization epitopes on VP4. J. Virol. 2008, 82, 6753–6757. [Google Scholar] [CrossRef]
- Navarro, A.; Trask, S.D.; Patton, J.T. Generation of genetically stable recombinant rotaviruses containing novel genome rearrangements and heterologous sequences by reverse genetics. J. Virol. 2013, 87, 6211–6220. [Google Scholar] [CrossRef]
- Criglar, J.; Greenberg, H.B.; Estes, M.K.; Ramig, R.F. Reconciliation of rotavirus temperature-sensitive mutant collections and assignment of reassortment groups D, J, and K to genome segments. J. Virol. 2011, 85, 5048–5060. [Google Scholar] [CrossRef]
- Komoto, S.; Fukuda, S.; Murata, T.; Taniguchi, K. Human Rotavirus Reverse Genetics Systems to Study Viral Replication and Pathogenesis. Viruses 2021, 13, 1791. [Google Scholar] [CrossRef]
- Kobayashi, T.; Antar, A.A.; Boehme, K.W.; Danthi, P.; Eby, E.A.; Guglielmi, K.M.; Holm, G.H.; Johnson, E.M.; Maginnis, M.S.; Naik, S.; et al. A plasmid-based reverse genetics system for animal double-stranded RNA viruses. Cell Host Microbe 2007, 1, 147–157. [Google Scholar] [CrossRef]
- Boyce, M.; Celma, C.C.P.; Roy, P. Development of reverse genetics systems for bluetongue virus: Recovery of infectious virus from synthetic RNA transcripts. J. Virol. 2008, 82, 8339–8348. [Google Scholar] [CrossRef]
- Guo, Y.Z.; Pretorius, J.M.; Xu, Q.Y.; Wu, D.L.; Bu, Z.G.; Theron, J.; Sun, E.C. Development and optimization of a DNA-based reverse genetics systems for epizootic hemorrhagic disease virus. Arch. Virol. 2020, 165, 1079–1087. [Google Scholar] [CrossRef]
- Kaname, Y.; Celma, C.C.P.; Kanai, Y.; Roy, P. Recovery Of African horse sickness virus from synthetic RNA. J. Gen. Virol. 2013, 94, 2259–2265. [Google Scholar] [CrossRef]
- Kanai, Y.; Kobayashi, T. Rotavirus reverse genetics systems: Development and application. Virus Res. 2021, 295, 198296. [Google Scholar] [CrossRef]
- Campagna, M.; Eichwald, C.; Vascotto, F.; Burrone, O.R. RNA interference of rotavirus segment 11 mRNA reveals the essential role of NSP5 in the virus replicative cycle. J. Gen. Virol. 2005, 86, 1481–1487. [Google Scholar] [CrossRef]
- Eichwald, C.; De Lorenzo, G.; Schraner, E.M.; Papa, G.; Bollati, M.; Swuec, P.; de Rosa, M.; Milani, M.; Mastrangelo, E.; Ackermann, M.; et al. Identification of a Small Molecule That Compromises the Structural Integrity of Viroplasms and Rotavirus Double-Layered Particles. J. Virol. 2018, 92, e01943-17. [Google Scholar] [CrossRef]
- Papa, G.; Venditti, L.; Arnoldi, F.; Schraner, E.M.; Potgieter, C.; Borodavka, A.; Eichwald, C.; Burrone, O.R. Recombinant Rotaviruses Rescued by Reverse Genetics Reveal the Role of NSP5 Hyperphosphorylation in the Assembly of Viral Factories. J. Virol. 2019, 94, e01110-19. [Google Scholar] [CrossRef]
- Smee, D.F.; Sidwell, R.W.; Clark, S.M.; Barnett, B.B.; Spendlove, R.S. Inhibition of rotaviruses by selected antiviral substances: Mechanisms of viral inhibition and in vivo activity. Antimicrob. Agents Chemother. 1982, 21, 66–73. [Google Scholar] [CrossRef]
- Kanai, Y.; Kawagishi, T.; Nouda, R.; Onishi, M.; Pannacha, P.; Nurdin, J.A.; Nomura, K.; Matsuura, Y.; Kobayashi, T. Development of Stable Rotavirus Reporter Expression Systems. J. Virol. 2019, 93, e01774-18. [Google Scholar] [CrossRef]
- Hatazawa, R.; Fukuda, S.; Kumamoto, K.; Matsushita, F.; Nagao, S.; Murata, T.; Taniguchi, K.; Matsui, T.; Komoto, S. Strategy for generation of replication-competent recombinant rotaviruses expressing multiple foreign genes. J. Gen. Virol. 2021, 102, e001587. [Google Scholar] [CrossRef]
- Philip, A.A.; Herrin, B.E.; Garcia, M.L.; Abad, A.T.; Katen, S.P.; Patton, J.T. Collection of Recombinant Rotaviruses Expressing Fluorescent Reporter Proteins. Microbiol. Resour. Announc. 2019, 8, e00523-19. [Google Scholar] [CrossRef]
- Philip, A.A.; Patton, J.T. Expression of Separate Heterologous Proteins from the Rotavirus NSP3 Genome Segment Using a Translational 2A Stop-Restart Element. J. Virol. 2020, 94, e00959-20. [Google Scholar] [CrossRef]
- Zhu, Y.; Sánchez-Tacuba, L.; Hou, G.; Kawagishi, T.; Feng, N.; Greenberg, H.B.; Ding, S. A recombinant murine-like rotavirus with Nano-Luciferase expression reveals tissue tropism, replication dynamics, and virus transmission. Front. Immunol. 2022, 13, 911024. [Google Scholar] [CrossRef]
- Philip, A.A.; Patton, J.T. Generation of Recombinant Rotaviruses Expressing Human Norovirus Capsid Proteins. J. Virol. 2022, 96, e01262-22. [Google Scholar] [CrossRef]
- Kotaki, T.; Kanai, Y.; Onishi, M.; Minami, S.; Chen, Z.; Nouda, R.; Nurdin, J.A.; Yamasaki, M.; Kobayashi, T. Generation of single-round infectious rotavirus with a mutation in the intermediate capsid protein VP6. J. Virol. 2024, 98, e00762-24. [Google Scholar] [CrossRef]
- Sun, C.; Gardner, C.L.; Watson, A.M.; Ryman, K.D.; Klimstra, W.B. Stable, high-level expression of reporter proteins from improved alphavirus expression vectors to track replication and dissemination during encephalitic and arthritogenic disease. J. Virol. 2014, 88, 2035–2046. [Google Scholar] [CrossRef]
- Fukuyama, S.; Katsura, H.; Zhao, D.M.; Ozawa, M.; Ando, T.; Shoemaker, J.E.; Ishikawa, I.; Yamada, S.; Neumann, G.; Watanabe, S.; et al. Multi-spectral fluorescent reporter influenza viruses (Color-flu) as powerful tools for studies. Nat. Commun. 2015, 6, 6600. [Google Scholar] [CrossRef]
- Philip, A.A.; Hu, S.; Dai, J.; Patton, J.T. Recombinant rotavirus expressing the glycosylated S1 protein of SARS-CoV-2. J. Gen. Virol. 2023, 104, 001899. [Google Scholar] [CrossRef]
- Kim, S.H.; Jang, Y.S. The development of mucosal vaccines for both mucosal and systemic immune induction and the roles played by adjuvants. Clin. Exp. Vaccine Res. 2017, 6, 15–21. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Q.H.; Xing, X.X.; Sun, D.B. A Mini-Review on the Common Antiviral Drug Targets of Coronavirus. Microorganisms 2024, 12, 600. [Google Scholar] [CrossRef]
- Kang, G.G.D.; Lakhkar, A.; Bhamare, C.; Dharmadhikari, A.; Narwadkar, J.; Kanujia, A.; Desai, S.; Gunale, B.; Poonawalla, C.S.; Kulkarni, P.S. Post-marketing safety surveillance of the rotavirus vaccine in India. Vaccine X 2023, 15, 100362. [Google Scholar] [CrossRef]
- Clark, A.; Tate, J.; Parashar, U.; Jit, M.; Hasso-Agopsowicz, M.; Henschke, N.; Lopman, B.; Van Zandvoort, K.; Pecenka, C.; Fine, P.; et al. Mortality reduction benefits and intussusception risks of rotavirus vaccination in 135 low-income and middle-income countries: A modelling analysis of current and alternative schedules. Lancet. Glob. Health 2019, 7, e1541–e1552. [Google Scholar] [CrossRef]
- Verkerke, H.; Sobuz, S.; Ma, J.Z.; Petri, S.E.; Reichman, D.; Qadri, F.; Rahman, M.; Haque, R.; Petri, W.A.J. Malnutrition Is Associated with Protection from Rotavirus Diarrhea: Evidence from a Longitudinal Birth Cohort Study in Bangladesh. J. Clin. Microbiol. 2016, 54, 2568–2574. [Google Scholar] [CrossRef]
- Du, Y.; Chen, C.; Zhang, X.; Yan, D.; Jiang, D.; Liu, X.; Yang, M.; Ding, C.; Lan, L.; Hecht, R.; et al. Global burden and trends of rotavirus infection-associated deaths from 1990 to 2019: An observational trend study. Virol. J. 2022, 19, 166. [Google Scholar] [CrossRef]
- Tissera, M.S.; Cowley, D.; Bogdanovic-Sakran, N.; Hutton, M.L.; Lyras, D.; Kirkwood, C.D.; Buttery, J.P. Options for improving effectiveness of rotavirus vaccines in developing countries. Hum. Vaccines Immunother. 2017, 13, 921–927. [Google Scholar] [CrossRef]
- Writing Group for Expert Consensus on Rotavirus Gastroenteritis. Expert consensus on immunoprophylaxis of childhood rotavirus gastroenteritis (2024 version). Chin. J. Prev. Med. 2024, 58, 1–33. [Google Scholar] [CrossRef]
- Pawar, S.D.; Kode, S.S.; Keng, S.S.; Tare, D.S.; Diop, O.M.; Abraham, P.; Sharma, D.K.; Sangal, L.; Yadav, P.D.; Potdar, V.A. Replication of SARS-CoV-2 in cell lines used in public health surveillance programmes with special emphasis on biosafety. Indian J. Med. Res. 2022, 155, 129–135. [Google Scholar] [CrossRef]
- Pathogen Regulation Directorate. Human Rotavirus. Available online: https://www.ehs.com/resources/sds-resources/free-safety-data-sheet-index/human-rotavirus/ (accessed on 1 September 2010).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Strategy | Inactivated Vaccines | Subunit Formulations | Oral Live-Attenuated Vaccines |
|---|---|---|---|
| Immune response | Humoral (Th2-biased) | Humoral (Th2-biased) | Mucosal IgA antibody + Systemic Th1/Th17 |
| Duration | Short duration | Short duration | Long-term |
| Cross-Protection | Limited (<30% serotype coverage) | None | Broad (>80% heterotypic immunity) |
| Safety concerns | Few | Few | Slight |
| Production | Easier | Easier | Challenging |
| Storage | Refrigeration | Refrigeration | Refrigeration |
| Year | Virus | Cell Line | Helper Virus | Helper Protein | Vector | Heterologous Proteins | Reference |
|---|---|---|---|---|---|---|---|
| 2006 | KU | COS-7 | KU | KU VP4 antibody | / | SA11 VP4 | [25] |
| 2010 | KU | COS-7 | tsE | NSP2 g8D siRNA | / | SA11 NSP2 | [28] |
| 2010 | RF | COS-7 | RF | / | / | Human NSP3 | [29] |
| 2015 | SA11 | BSR5/T7 | tsA | VP4 siRNA | / | Chicken VP4 | [30] |
| 2017 | SA11 | BHK-T7 | / | FAST, D1R, D12L | / | / | [26] |
| 2018 | SA11 | BHK-T7 | / | NSP2, NSP5 | / | / | [31] |
| 2020 | SA11 | BHK-T7 | / | C3P3-G1 | / | / | [32] |
| 2020 | SA11 | BHK-T7 | / | C3P3-G1 | / | Human VP4, VP7, VP6 | [33] |
| 2021 | SA11 | BHK-T7 | / | C3P3-G1 | NSP3 | SARS-CoV-2 S1 | [34] |
| 2022 | SA11 | BHK-T7 | / | NSP2, NSP5, D1R, D12L | VP4 | SARS-CoV-2 S1 | [35] |
| 2022 | RF | BHK-T7 | / | NSP2, NSP5, D1R, D12L | NSP3 | SARS-CoV-2 S1 | [35] |
| 2023 | SA11 | BHK-T7 | C3P3-G1 | NSP3 | HuNoV VP1 | [36] | |
| 2024 | BLR | BHK-T7 M1 | / | / | / | [37] | |
| 2024 | C73 | BHK-T7 M1 | / | / | / | / | [37] |
| 2024 | HM26 | BHK-T7 M1 | / | / | / | [37] | |
| 2024 | SA11 | BHK-T7 | C3P3-G1 | NSP1 | HSV-2 gD | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Qin, S.; Li, K.; Yin, X.; Sun, D.; Chang, J. Rotavirus Reverse Genetics Systems and Oral Vaccine Delivery Vectors for Mucosal Vaccination. Microorganisms 2025, 13, 1579. https://doi.org/10.3390/microorganisms13071579
Wang J, Qin S, Li K, Yin X, Sun D, Chang J. Rotavirus Reverse Genetics Systems and Oral Vaccine Delivery Vectors for Mucosal Vaccination. Microorganisms. 2025; 13(7):1579. https://doi.org/10.3390/microorganisms13071579
Chicago/Turabian StyleWang, Jun, Songkang Qin, Kuanhao Li, Xin Yin, Dongbo Sun, and Jitao Chang. 2025. "Rotavirus Reverse Genetics Systems and Oral Vaccine Delivery Vectors for Mucosal Vaccination" Microorganisms 13, no. 7: 1579. https://doi.org/10.3390/microorganisms13071579
APA StyleWang, J., Qin, S., Li, K., Yin, X., Sun, D., & Chang, J. (2025). Rotavirus Reverse Genetics Systems and Oral Vaccine Delivery Vectors for Mucosal Vaccination. Microorganisms, 13(7), 1579. https://doi.org/10.3390/microorganisms13071579