Abstract
The interplay between comorbidities and viral infections is a critical factor that influences disease severity and outcomes. Diabetes Mellitus (DM) is one such comorbidity that significantly elevates the risk of severe viral infection from coronaviruses, namely, SARS-CoV-2. DM is characterized by either a lack of insulin production (type 1 diabetes) or insulin resistance (type 2 diabetes), both of which contribute to a state of hyperglycemia, or high blood sugar. Hyperglycemia significantly promotes chronic inflammation, metabolic dysfunction, and immune dysregulation, which put diabetics at an elevated risk of critical health outcomes. Additionally, diabetes is hypothesized to amplify viral titers during infection by promoting the expression of the viral entry receptor ACE2 and providing a favorable cellular energy environment for viral replication. This review focuses on explaining the mechanisms that link diabetics with more severe COVID-19 disease and exploring some of the mechanisms that contribute to the phenomenon where COVID-19 can promote new-onset diabetes. By highlighting the interconnections between diabetes and COVID-19, this review aims to emphasize the implications that the SARS-CoV-2 outbreak has had on metabolic health.
1. COVID-19
In late 2019, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spilled over into the human population from an unknown zoonotic source in Wuhan, China, and resulted in a global pandemic of coronavirus disease-2019 (COVID-19) [1,2,3]. Since then, more than 1.2 million people have died from COVID-19 in the United States alone (CDC COVID data tracker). SARS-CoV-2 is a positive-sense, single-stranded RNA virus that belongs to the genus Betacoronavirus (β-CoV). β-CoVs have garnered significant public interest that began with SARS-CoV (SARS) and then MERS-CoV (MERS) spilling over into human populations from zoonotic reservoirs and causing outbreaks [4]. The SARS-CoV epidemic caused an outbreak that began in the Guangdong province of China and spread to nearly 30 other countries in 2002–2003 [5]. Similarly, the MERS-CoV outbreaks occurred from 2012 to 2015 in the Middle East, and there was also a large outbreak in South Korea in 2015 that originated from a single person who had traveled to the Middle East [6,7]. Like SARS-CoV, the primary receptor for SARS-CoV-2 is the angiotensin-converting enzyme-2 (ACE2) receptor [8,9]. ACE2 is a highly expressed protein receptor in the upper and lower respiratory epithelium but is also expressed in other tissues, including the kidneys and intestines [10]. Although SARS-CoV-2 infections in humans primarily take place in the upper and lower respiratory tracts, there is strong evidence that infection occurs in the liver [11], intestinal tract [12], brain [13], and pancreas [14]. SARS-CoV-2 infections can cause oxygen and metabolic changes that contribute to damage to the heart, brain, kidney, and vascular tissues [15,16].
COVID-19 symptoms range from asymptomatic to severe [17,18]. Mild symptoms include fever, chills, body aches, and loss of smell, while severe symptoms include shortness of breath, chest pain, and intense coughing that can progress to acute respiratory distress syndrome (ARDS). If symptoms worsen, patients may require supplemental oxygen or intratracheal ventilation. Vascular dysfunction, such as blood clots and thrombosis, can also manifest as a result of COVID-19 [19]. The long-lasting impacts of COVID-19, termed long-COVID, are well-documented and may present as neurological and cardiovascular sequelae [20,21,22]. Acute and long-term COVID-19 complications can be especially devastating to individuals with pre-existing conditions that put them at a health disadvantage. This review will focus on the intersection of COVID-19 with diabetes, a very common comorbidity. Table 1 highlights some of the studies on COVID-19 patients discussed herein with respect to diabetes.

Table 1.
Clinical studies of COVID-19 patients and diabetes.
2. Diabetes Has Emerged as an Important Comorbidity That Markedly Increases the Severity of COVID-19
Diabetes incidence is on the rise worldwide, affecting both affluent and poor communities in emerging and developed nations [39,40]. According to the CDC National Center for Health Statistics, it was the seventh leading cause of death among Americans in 2023 [41], and almost 16% of the adult population was living with diabetes as of 2023 [42]. Diabetics cannot regulate their blood glucose levels because of insufficient insulin production (type 1 diabetes) or insulin insensitivity (type 2 diabetes). Hyperglycemia, or high blood glucose, is the main factor from which the downstream complications of diabetes stem [43,44]. Specifically, hyperglycemia primarily affects vascular tissue and can lead to sustained inflammation, organ failure, and nephropathy [43,44,45]. This constellation of disease manifestations that contribute to diabetes are recognized as key factors that significantly increase the risk of severe COVID-19 [23,24]. As such, diabetes is considered an immunocompromising disease [46].
2.1. Meta-Analysis of Diabetes Representation Among Persons with β-CoVs (Epidemiology)
Diabetes emerged as a prominent comorbidity associated with SARS and MERS [47,48]. During the first SARS epidemic in 2002–2003, diabetes was associated with mortality in Hong Kong [48]. In a study of hospitalized patients in Beijing during the 2003 SARS epidemic, the percentage of patients with a diabetes diagnosis was higher in the deceased group than the survivor group, indicating an association between diabetes and risk of death [49]. Additionally, infection with SARS-CoV caused an increase in fasting plasma glucose compared to uninfected control patients [50]. Data from the MERS epidemic shows that diabetes status was significantly associated with contracting MERS illness [51]. Moreover, diabetes was significantly associated with a higher mortality rate in MERS-CoV-infected individuals [52].
Similar conclusions have been drawn with respect to increased COVID-19 hospitalization rates for individuals with diabetes. In a cohort of 5700 COVID-19 patients from the metropolitan area of New York City, diabetes stood as the third most common comorbidity at roughly 33%, which was behind hypertension (57%) and obesity (42%) [25]. In a study in England, the risks of comorbidities in COVID-19 cases were assessed by surveying the health records of 61,440,470 individuals [26]. Analyses of the COVID-19-related deaths that occurred in-hospital between 1 March and 11 May of 2020 found that 31.4% of individuals had type 2 diabetes and 1.5% had type 1 diabetes. This is in contrast to the proportions in the entire sample group, where they found that 4.7% had type 2 diabetes and 0.4% had type 1 diabetes. These findings indicate that both type 1 and type 2 diabetes were associated with an increased risk of mortality from COVID-19.
There have been mixed findings on the protection afforded by previous SARS-CoV-2 infection or vaccination in diabetic patients. In a meta-analysis of the outcomes of 17 different published articles, vaccine efficacy had significantly waned in the diabetic population compared to healthy controls [27]. Unsurprisingly, vaccinated diabetics had a significantly higher odds ratio for SARS-CoV-2 breakthrough infection compared to healthy controls. This was supported in animal studies, where humoral responses induced by the BNT162b2 mRNA vaccine were reduced in an HFD diabetic model of C57BL/6J mice compared to healthy controls [53]. However, other studies in humans found that the humoral response in a vaccinated diabetic cohort was similar to that of a vaccinated non-diabetic control cohort [28].
2.2. Why Diabetes Contributes to Worse Outcomes
Hyperglycemia induces the generation of reactive oxidative stress (ROS) factors, which in turn promote inflammation [54]. Additionally, diabetes is often accompanied by obesity, which likewise promotes a high basal level of inflammation. Visceral adipose tissue promotes a significantly pro-inflammatory environment in the individual and leads to secretion of TNF-α, IL-6, IL-8, and adipokines that may amplify other immune responses [55,56,57]. This can be particularly deadly, especially for SARS-CoV-2, if it leads to a “cytokine storm”, which is an outsized, dysregulated immune response to the virus. The excess or overproduction of these immune factors can cause acute respiratory stress and organ damage in severe COVID-19. The blockade of IL-6 in particular has shown some success as a treatment option for severe and critical COVID-19 in clinical trials [58].
Diabetic panvascular disease (DPD) is another disorder that has a severe impact on the normal homeostatic function of multiple organs, including the heart, kidneys, and lungs [59]. DPD manifests as macro- and microvascular atherosclerosis, which contributes to endothelial damage in the aforementioned organs [60]. Hyperglycemia has been implicated as a leading factor in inducing lung injury in diabetics [61]. Preexisting injury in the lungs and other organs puts diabetic individuals at a disadvantage when fighting a viral infection.
2.3. Clinical Findings and Indicators Associated with Worse Outcomes (Pathogenesis)
The immunocompromising factors (i.e., vascular damage in the lung, and pro-inflammatory microenvironment) of diabetes are proposed to increase the risk of death and severe pathology from COVID-19 [29]. A study in a Swedish cohort found that type 2, but not type 1, diabetes was independently associated with an elevated risk of hospitalization for COVID-19 patients [30]. Elevated HbA1C levels in the type 2 diabetic cohort were associated with a higher risk of worse outcomes. In addition, a higher body mass index (BMI) index was independently associated with a higher risk of worsened outcomes. This would imply that uncontrolled diabetes and diabetes-caused obesity promote worsened disease.
A retrospective study in the USA analyzed the risk of death based on HbA1c levels among T2D COVID-19 patients and found that the risk of death increased with increasing HbA1c levels [31]. A Japanese study of SARS-CoV-2-infected individuals in 2020–2021 reported that the proportion of critical outcomes (supplemental oxygen, invasive ventilation, or death) was higher in the diagnosed and undiagnosed diabetic cohorts, 30% and 35%, respectively, compared to the non-diabetic group, where it was 5% [32]. Additionally, the proportion of undiagnosed and diagnosed diabetics who accumulated complications, such as bacterial infections, thromboembolism, or acute kidney injury, was also higher than recorded in the non-diabetic group. High HbA1c levels, a measurement of average blood glucose concentration, correlated to worse COVID-19 outcomes. Lastly, a study in Italy found a higher abundance of SARS-CoV-2-positive staining in postmortem cardiac tissue of diabetics [33].
3. The Incidence of Diabetes Is Higher in Individuals After COVID-19
SARS-CoV, the causative agent of the 2002 epidemic, was correlated with elevated plasma glucose levels. Two studies examined cohorts of individuals hospitalized with SARS-CoV and found that their fasting plasma glucose levels were elevated compared to the non-SARS-CoV-infected groups [49,50].
Similarly, COVID-19 has been reported to induce hyperglycemia and new-onset diabetes, where a systematic review of new-onset diabetes after COVID-19 in the USA, Germany, and England found a significant association [34]. SARS-CoV-2 infection was associated with a rise in the incidence of diabetes, hyperlipidemia, and hypertension [34]. Vaccinated cohorts did not have a rise in diabetes, hyperlipidemia, and hypertension [35]. A 551-patient cohort of COVID-19 in Italy additionally found a significant correlation between the patients and their hyperglycemic status. Patients who had recorded hyperglycemia upon hospital admission had persistent hyperglycemia for at least 6 months [36]. They reported individuals with no history of diabetes presenting with new-onset diabetes and insulin resistance after having COVID-19. A study of over 700 patients from Brazil who had been hospitalized with COVID-19 from March to August 2020 found that the proportion of hyperglycemia incidence was higher in the COVID-19-positive patients than the COVID-19-negative patients, regardless of diabetes diagnosis [11]. Not only does this phenomenon occur in adults, but it has been observed in pediatric populations as well, where a cohort of over 600,000 patients aged 10–19 revealed that the risk of new-onset diabetes was raised in pediatric patients within 6 months of COVID-19 diagnosis [37].
3.1. Potential Mechanisms Proposed for New-Onset Diabetes Post-COVID-19
SARS-CoV-2, like all viruses, relies on host cell machinery to replicate its genetic material and complete its life cycle. A hallmark of viruses is their ability to hijack cellular functions to promote their viral life cycle. A common target of viruses is the host’s central carbon metabolism pathways.
Central carbon metabolism refers to the interconnected biochemical cycles that produce energy from carbohydrates. Mammalian cells primarily rely on aerobic respiration to metabolize glucose into usable energy, ATP. In brief, glucose is shuttled into the cell and immediately enters the cycle of glycolysis and generates two new molecules of ATP and one molecule of acetyl-CoA, the input for the citric acid cycle. The process occurs in the cytosol independent of oxygen and can compensate for energy production in the cell when oxygen is present at sub-optimally low concentrations. Aerobic respiration is the most efficient method of glucose metabolism in the presence of oxygen. The citric acid cycle produces succinate, which undergoes oxidative phosphorylation (OXPHOS) in the cristae of the mitochondria and generates the bulk product of ATP (36 molecules). It is important to note that additional byproducts are made throughout these metabolic processes, including serine, glycine, and nucleotide precursors. The pentose phosphate pathway runs in tandem with glycolysis and is responsible for the production of ribose-5-phosphate, an important precursor to DNA and RNA.
It should come as no surprise that viruses commonly target host bioenergetic systems to promote their replication. The exact mechanism of how viruses hijack these systems varies, but one hypothesis proposes viruses modulate host pathways to increase the production of nucleotides and other substrates/macromolecules that are necessary for the assembly of mature virions [62]. For example, influenza A virus (IAV) infection induces glycolysis in vitro and in vivo [63,64]. An association between glycolysis activity and IAV titers was demonstrated, where inhibition of glycolysis prevented viral replication [64]. There are many more examples of viruses that increase glycolysis, including Zika virus [65], dengue virus [66], and human cytomegalovirus [67].
Another important group is the oncogenic viruses, such as human papillomavirus (HPV), hepatitis B virus (HBV), and Epstein–Barr virus (EBV), which activate host cell glycolysis in what is known as the ‘Warburg effect’ [68]. The Warburg effect is the phenomenon in which cancer cells have a heightened dependency on energy generation by glycolysis, unlike non-cancerous cells, which depend primarily on aerobic respiration and OXPHOS. Oncogenic viruses contribute to a cell becoming cancerous by activating oncogenes, but they simultaneously activate host cell glycolysis, mirroring the Warburg effect that can be observed in cancer cells [69]. The propensity for viruses to interfere with host cell metabolism could explain the new onset of diabetes after COVID-19.
3.2. SARS-CoV-2 Targets Mitochondrial Function
In A549 cells, a continuous lung adenocarcinoma cell line, transfection of plasmid expressing the nonstructural proteins (nsp) of SARS-CoV-2, ORF9b, ORF9c, and ORF10 interact directly with mitochondrial proteins [70]. The nsp produced by ORF3a downregulated the expression of proteins that have a role in maintaining mitochondrial function [70]. Expression of the nsp from ORF9b promoted fragmentation of mitochondria, and expression of the nsp from the ORF9c also altered cristae structure. The nsp from the expression of ORF10 promoted the fusion of mitochondria. The study was limited to only being shown in vitro, but it does provide a starting point to further investigate the role of the viral proteins in the alteration of host mitochondrial function.
Comparing RNA-Seq data across SARS-CoV-2-infected A549, A549-ACE2, Calu-3, and NHBE cells in addition to lung tissue from autopsies and bronchoalveolar lavage fluid (BALF) from patients suggested that SARS-CoV-2 downregulated mitochondrial ribosome protein genes in primary cell culture compared to IAV [71]. Mitochondrial Complex 1 gene and cellular respiration-related genes were downregulated in SARS-CoV-2-infected cell lines compared to cells infected with IAV, human parainfluenza viruses (HPIV), or respiratory syncytial virus (RSV). In nasopharyngeal samples from infected patients, mitochondrial OXPHOS gene expression was downregulated and correlated with the peak of viral RNA levels [72]. An inverse association was also observed with OXPHOS gene expression recovering as the viral RNA in the sample declined.
3.3. Disruption of Glucometabolic Control and Promotion of Glycolysis After SARS-CoV-2 Infection
SARS-CoV-2 infection has been shown to affect the protein expression of two major glucose importers, GLUT1 and GLUT4, (Figure 1) in the lung and cardiac tissue in a feline model [73]. The group infected with the SARS-CoV-2 Delta variant had increased GLUT1 expression in the heart and lung tissue at 4 days post-infection (dpi) and 12 dpi, and the GLUT4 insulin-sensitive glucose transporter was increased in heart and lung tissue at 4 dpi [73]. In infected Vero E6 TMPRSS2 cells, glucose and folate were depleted after SARS-CoV-2 infection, while lactate was increased, suggesting that glycolysis was increased [74]. Conversely, galactose supplementation did not promote SARS-CoV-2 replication. It is important to note that galactose is an input for the OXPHOS cycle, while glucose can be used for glycolysis.
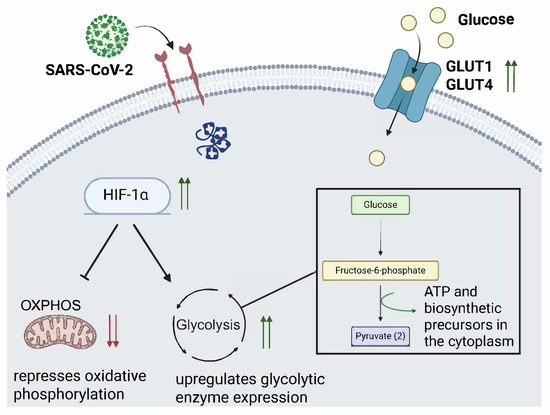
Figure 1.
SARS-CoV-2 infection promotes HIF-1α activity and glucose transporter expression. The illustration depicts a proposed model by which SARS-CoV-2 infection promotes glycolysis through upregulation of hypoxia inducible factor 1 alpha (HIF-1α) and glucose transporters (GLUT1 and GLUT4). Green upwards arrows indicate HIF-1α expression is increased, which drives glycolysis, shown by the black arrow. The bar-headed arrow emerging from HIF-1α indicates that oxidative phosphorylation activity is decreased by HIF-1α activity. GLUT1 and GLUT4 are pictured on the surface of the cell with two green arrows also indicating their increased expression from SARS-CoV-2 infection. The two red arrows indicate decreased expression.
During SARS-CoV-2 infection, the accessory protein ORF3a was shown to promote Hypoxia-inducible factor-1 (HIF1α) production in a Caco2 cell culture [75]. Additionally, HIF-1α expression was higher in peripheral blood mononuclear cells (PBMCs) from COVID-19 patients than in healthy patients. HIF-1α boosts glycolysis activity and can be leveraged by other viruses, such as RSV, to promote their replication [76].
It has also been reported that SARS-CoV-2 infection upregulates the HIF1α-dependent glycolysis axis in monocytes [77]. Monocyte dysfunction has been identified as a major driver of acute COVID-19 [78,79]. Inhibitors of PFKFB3 (3-PO), glycolysis (2-DG), and LDH-A (Oxamate) block SARS-CoV-2 replication in monocytes. However, an inhibitor of the mitochondrial pyruvate carrier was unable to do the same, implying that TCA cycle activity is not important for viral replication. IAV and RSV increased glycolytic activity in monocytes to a lesser extent than SARS-CoV-2 [77]. This would, in effect, induce aerobic glycolysis to sustain viral replication in human monocytes.
Using a model of SARS-CoV-2 infection in human pluripotent stem cell (hPSC)-derived organoids, which were differentiated into airway organoids, an inhibitor of glycolysis (GW6471) was able to block SARS-CoV-2 WA1 infection [80]. Chetomin and shRNA specific for HIF1 α also blocked SARS-CoV-2 infection. Metabolic profiling showed upregulation of D-glucose-6 phosphate, citric acid, fatty acids, and amino acids, suggesting that SARS-CoV-2 infection promoted glycolysis in the hPSC model. This phenomenon has been observed in other viral infections, including RSV upregulation of HIF1α [76].
SARS-CoV-2 could induce a shift from the OXPHOS cycle to glycolysis to support its proliferation. In doing so, the host cell’s glucometabolic control would be significantly skewed towards glycolysis, and as more cells and tissues are infected, so too would the glucometabolic dysregulation. The hypothesis is that widespread glycolysis upregulation would require a high amount of insulin to promote glucose uptake by the infected cells. Sustained high levels of insulin could promote insulin resistance in the host and contribute to new-onset type 2 diabetes diagnosis even after recovery.
3.4. COVID-19 Can Cause Direct and Indirect Damage to the Pancreas
Pancreatic dysfunction is often a key contributor to type 1 diabetes. The pancreas produces the insulin necessary to regulate the body’s blood glucose levels. However, in cases of severe autoimmunity or insensitivity from an unhealthy diet, it can fail to maintain acceptable blood glucose levels and lead to an individual’s diabetes diagnosis.
In a cohort of infected and infected + vaccinated non-human primates (NHP), the elder NHPs had a higher immunopositivity for the spike protein after infection compared to the adult NHPs, in addition to more severe lesions and amyloid deposits in the pancreas, hallmarks of type 2 diabetes cases [81]. Another group conducted a histological analysis of pancreas sections from COVID-19 autopsy patients and found more extensive microthrombi, fibrosis, and endothelialitis compared to the samples from the uninfected patients [14].
By infecting pancreatic islets collected from healthy donors with SARS-CoV-2, it was shown that they are susceptible to infection and subsequent β cell impairment and death [82]. Histological samples from COVID-19 patient autopsy tissue also found that the pancreatic beta cells stained positive for SARS-CoV-2 nucleoprotein (NP) in 7/9 patients confirmed COVID-19 positive by PCR. Another study conducted immunostaining on tissues from COVID-19 patient autopsy tissue and found SARS-CoV-2 infection in pancreatic β cells as well [14]. Infection of these cells would likely be accompanied by immune cell infiltration. There was one reported case of new-onset type 1 diabetes after COVID-19 in a 19-year-old patient who had severe β cell dysfunction for weeks after the infection cleared [83]. The authors concluded that the diabetes diagnosis was causally linked to the direct damage of the pancreas β cells as a result of the SARS-CoV-2 infection.
Other viruses have been speculated to cause new-onset type 1 diabetes in patients by directly damaging the pancreas [84], but larger cohort studies are needed to fully characterize the effect. Likewise, more studies are necessary to determine the extent to which SARS-CoV-2 can cause sufficient pancreatic damage to promote diabetes.
3.5. COVID-19 Vaccination Has Been Speculated to Induce Latent Autoimmune Diabetes in Adults (LADA)
LADA is a slow-onset version of diabetes that shares some characteristics with both type 1 and type 2 diabetes. Similar to type 2 diabetes, LADA is typically acquired in individuals over the age of 30, and as such, patients are often initially misdiagnosed as having type 2 diabetes. Unlike type 2 diabetes and similar to type 1 diabetes, it is immune-mediated, where autoimmune damage of an individual’s pancreatic β cells caused by a combination of CD8+ T cells, CD4+ T cells, B cells, macrophages, and dendritic cells impairs pancreas function [85]. Importantly, COVID-19 vaccination with the mRNA vaccine BNT162b2 can give rise to other autoimmune diseases, such as Graves disease [86]. It has also been speculated to drive the development of LADA in one documented case where a prediabetic individual who, soon after receiving the second dose of BNT162b2, had been admitted to a hospital for a diabetic ketoacidosis episode and tested positive for autoantibodies [38]. There would need to be larger-scale studies to establish a clear causal relationship between COVID-19 vaccination or infection and the induction of LADA, but it should be considered as a potential contributing factor to new-onset diabetes.
4. In Vivo Models Used to Study SARS-CoV-2 Infection in a Diabetic Host
Three main models of diabetes-induced mice have been used to study SARS-CoV-2 infection. They include the high-fat diet mice (HFD), db/db mice, and streptozotocin (STZ)-induced diabetic mice. Each approach simulates diabetic conditions, but they differ slightly in the pathogenic outcomes. Additionally, special attention should be given to whether the study used a mouse-adapted strain of SARS-CoV-2 or if it was propagated from a human isolate, as this can impact interpretations.
4.1. Db/Db (Leptin Deficient) Mouse Model
Db/db mice are deficient in the leptin receptor [87]. These mice are less capable of lowering blood glucose levels and have dysregulated metabolism. The C57BL/6J db/db animal model presents local swelling of the liver and worse pathological scores in the lung, liver, and pancreas compared to the uninfected and infected WT mice, indicating that the db/db model recapitulates severe diabetic complications [88]. Infecting C57BL/6J db/db mice with SARS-CoV-2 B.1.1.7 (Alpha Variant of Concern) showed that they were more susceptible to infection and severe pathological damage as compared to the wild-type (WT) mice [88]. The infected C57BL/6J db/db mice had worse pathological scores based on H&E staining of lung tissue, which included more intense pulmonary hemorrhage. Additionally, the SARS-CoV-2 Alpha-infected C57BL/6J db/db mice had higher viral gRNA recovered from lungs and throat swabs compared to the WT group.
The K18-hACE2 db/db mouse model, which is transgenic for the human ACE2 receptor, also suggested that infection with SARS-CoV-2 is enhanced in a diabetic host. Infecting these animals with the SARS-CoV-2 strain hCoV-19/China/CAS-B001/2020 (CAS-B001) showed that blood glucose levels were slightly higher in the diabetic group compared to the non-diabetic group [89]. Staining for SARS-CoV-2 N protein in the lung tissue showed that the diabetic mice supported more widespread virus replication with higher histopathological scores. Cathepsin L (CTSL) activity in db/db mice suggested that CTSL, a cofactor that promotes SARS-CoV-2 entry, is more active in the db/db mice [90]. This could explain why virus replication was elevated in these animals.
4.2. HFD Mouse Model
The HFD mouse model offers a more natural model of diabetes that has been caused by lifestyle. The mice are restricted to eating chow with a higher fat content and experience significant weight gain and become glucose-resistant after about three months on the diet. When diabetes was induced in C57BL/6 mice by keeping them on an HFD for 12–17 weeks, infecting them with MERS-CoV resulted in prolonged histopathological inflammation in the lung tissue compared to non-diabetic mice but had similar levels of virus [47]. Immunizing HFD animals with the mRNA vaccine BNT162b2 or a recombinant nanoparticle vaccine (NVX-CoV2373) and then infecting them at least two weeks later with a mouse-adapted strain of SARS-CoV-2 (MA/10) suggested virus clearance was delayed in the HFD group compared to the CD group. In the non-vaccinated animals, there was increased mortality, but not viral loads, in the HFD group compared to the control diet (CD) group after infection [53,91].
In contrast, K18-hACE2 mice with HFD-induced diabetes exhibited a similar course of infection to the CD group based on viral quantitation by qPCR at 3 dpi and histopathological analysis after infection with SARS-CoV-2 (hCoV-19/China/CAS-B001/2020) [89]. This implies that important differences between the model implementations, such as mouse and virus strain, can lead to data interpretations that vary widely.
4.3. Streptozotocin (STZ) Mouse Model
STZ induces diabetes in mice by damaging the pancreatic beta cells of the mouse and thereby reducing insulin production. Because the mouse cannot generate insulin to lower blood glucose levels after STZ treatment, it is considered a model of type 1 diabetes. There are limited studies that have been published on the effect of STZ injection on SARS-CoV-2 infection. However, one study used a model where hACE2 knock-in mice were injected with STZ, and then the animals were switched to an HFD for eight weeks to induce diabetes. Intravenously injecting the SARS-CoV-2 spike and nucleoprotein into the jugular significantly increased TLR signaling (TLR8, MyD88, and TRAF6) in the diabetic mice compared to control mice [92].
A study using STZ-treated mice as a diabetic model to study their reaction to S-protein immunization showed that STZ mice had a reduced IgG reaction at 7 weeks post-immunization compared to the non-diabetic mice [93]. However, mice were not subsequently challenged with the virus. Similar to what was found in db/db mice, induced diabetes in BALB/c mice using STZ and treating with insulin showed that CTSL, the host protease that exposed the Spike fusion peptide, mRNA expression was increased in diabetic mice [94].
5. In Vitro Models of SARS-CoV-2 Infection in Diabetic Conditions
While in vivo animal models of diabetes are useful for recapitulating human disease, in vitro models are important to highlight the cellular mechanisms at play. There are, however, fewer reports that use in vitro cell culture models of diabetes to study SARS-CoV-2 infection.
5.1. Continuous Cell Lines
It has been reported that ACE2 mRNA expression increases with increases in glucose concentration in a continuous lung epithelial cell line, Calu-3 cells [95]. BAY-876, a GLUT1 inhibitor, could prevent the increase in ACE2 mRNA expression, indicating that glucose influx into the cell leads to ACE2 mRNA upregulation. Similarly, one study showed that high glucose promoted mRNA expression of ACE2 in A549 cells [96], while another showed that D-galactose significantly increased the expression of cell receptors FURIN, TMPRSS2, and ACE2 [97].
Since ACE2 is a receptor for SARS-CoV-2 entry, these data suggested that persons with hyperglycemia may show greater susceptibility to SARS-CoV-2 infection. The increased expression of all three receptors is highly expected to increase susceptibility to SARS-CoV-2 infection. Indeed, there are in vitro studies that report that higher levels of glucose result in higher levels of virus production. In a study that infected monocytes with an isolate of SARS-CoV-2 from the second confirmed case in Brazil (GenBank: MT126808.1) and then supplemented the cells with increasing glucose concentrations (0, 2.5, 5.5, 11.1, and 22.2 mM), viral loads increased in association with glucose concentration after 24 h [77]. The increased viral load was attributed to the HIF-1α regulation of glycolysis.
High glucose also promoted infection with a SARS-CoV-2 pseudovirus in immortal human liver cells, Huh7, but not in CTSL knockout cells [90]. Greater pseudovirus infection in Huh7 cells grown in sera from diabetic donors has been observed, with high glucose correlating with higher CTSL activity, which may promote SARS-CoV-2 infection.
5.2. Primary Kidney and Brain Cell Lines
Using cells isolated from human donors to study SARS-CoV-2 infection increases the translatability of findings from other in vitro and in vivo models. In one study, human proximal tubular cells (HPTCs) isolated from the kidneys of diabetic individuals had higher basal respiration, ATP production, and ACE2 expression [98]. The cells also stained more positively for SARS-CoV-2 infection in addition to higher SARS-CoV-2 mRNA. Treating the cell samples with dichloroacetate (DCA), an inhibitor of pyruvate dehydrogenase kinase (PDK), decreased SARS-CoV-2 mRNA expression, suggesting that glycolysis activity is important for infection.
Culturing of human brain microvascular endothelial cells in either normal or high glucose medium and treating with SARS-CoV-2 spike protein suggested that cells grown in high glucose had higher concentrations of IL-6, TNF-α, and IL-1B compared to cells grown in a normal glucose environment [92].
5.3. Kidney Organoid
Organoids are growing in popularity due to greater sophistication as compared to 2D cell culture models and less intensive handling procedures compared to animal models. One group modeled SARS-CoV-2 infection in a kidney organoid model, which was subjected to high glucose oscillations (25mM and 5mM oscillated) to induce a diabetic state [98]. In their model, high glucose oscillations promoted the buildup of fibronectin and collagen fiber deposits, which mirrors some of the pathology consistent with chronic kidney disease and renal fibrosis, which can affect diabetics [99,100]. Hexokinase 2 (HK2) and LDHA genes, important enzymes in glycolysis, were upregulated [98]. Additionally, high glucose oscillation promoted gene and protein expression of ACE2. They found that SARS-CoV-2 infection was promoted by the high glucose oscillations compared to the low glucose control group at 1 dpi.
6. Discussion
Along with the elderly and immunocompromised populations, diabetics are considered at high risk of developing severe COVID-19. In general, clinical trials, animal studies, and cell-based models suggest that there is a strong bidirectional relationship between COVID-19 and diabetes. Clinical studies indicate that individuals with diabetes face more severe symptoms of COVID-19, with diabetes being one of the most overrepresented comorbidities among individuals hospitalized with COVID-19. SARS-CoV-2 has also been shown to replicate to higher titers in diabetic animal models and to associate with glucose concentration in cell culture models. Several animal models have revealed that vaccinated diabetic mice take longer to clear the virus after infection compared to control mice. Additionally, cell culture models have been used to demonstrate that hyperglycemia could promote SARS-CoV-2 infection and replication in the host cell. Additionally, there is a heightened proinflammatory response in diabetic mice.
Animal and cell culture models have provided ample evidence that SARS-CoV-2 infection dysregulates the metabolic cycle in the host at the cellular level and across tissues. This offers a strong foundation for the hypothesis that implicates COVID-19 as a contributor to new-onset diabetes in recovered patients. Special attention should continue to be placed on seeking out more efficient therapeutics, especially for the diabetic population, to counteract their overrepresentation in hospitalized cohorts. It should also be emphasized by medical professionals that those who overcome severe COVID-19 should monitor their blood glucose for a period after recovery to mitigate the incidence of new-onset diabetes. Additionally, there is an obvious gap in sophisticated in vitro models that can replicate the intracellular and intercellular hallmarks of diabetes, their susceptibility to SARS-CoV-2 infection, and the host–pathogen interactions of SARS-CoV-2 infection and diabetes.
Author Contributions
Conceptualization, J.H.N. and C.B.J.; methodology, J.H.N.; writing—original draft preparation, J.H.N.; writing—review and editing, J.H.N., A.M.S. and C.B.J.; data curation, J.H.N., A.M.S. and C.B.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
J.H.N. acknowledges the financial support from the Harriet Van Vleet Chair of Excellence in Virology to C.B.J, mentorship from C.B.J. and the collaboration from A.S.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Hui, D.S.; I Azhar, E.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; McHugh, T.D.; Memish, Z.A.; Drosten, C.; et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health 2014; The latest 2019 novel coronavirus outbreak in Wuhan, China. Int. J. Infect. Dis. 2020, 91, 264–266. [Google Scholar] [CrossRef] [PubMed]
- de Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Lam, W.; Zhong, N.; Tan, W. Overview on SARS in Asia and the World. Respirology 2003, 8, S2–S5. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Kang, J.-M.; Ha, Y.E.; Park, G.E.; Lee, J.Y.; Ko, J.-H.; Lee, J.Y.; Kim, J.M.; Kang, C.-I.; Jo, I.J.; et al. MERS-CoV outbreak following a single patient exposure in an emergency room in South Korea: An epidemiological outbreak study. Lancet 2016, 388, 994–1001. [Google Scholar] [CrossRef]
- Ramadan, N.; Shaib, H. Middle East respiratory syndrome coronavirus (MERS-CoV): A review. Germs 2019, 9, 35–42. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e899. [Google Scholar] [CrossRef]
- Li, M.-Y.; Li, L.; Zhang, Y.; Wang, X.-S. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues. Infect. Dis. Poverty 2020, 9, 45. [Google Scholar] [CrossRef]
- Barreto, E.A.; Cruz, A.S.; Veras, F.P.; Martins, R.; Bernardelli, R.S.; Paiva, I.M.; Lima, T.M.; Singh, Y.; Guimarães, R.C.; Damasceno, S.; et al. COVID-19-related hyperglycemia is associated with infection of hepatocytes and stimulation of gluconeogenesis. Proc. Natl. Acad. Sci. USA 2023, 120, e2217119120. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Tao, W.; Flavell, R.A.; Zhu, S. Potential intestinal infection and faecal–oral transmission of SARS-CoV-2. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 269–283. [Google Scholar] [CrossRef]
- Stein, S.R.; Ramelli, S.C.; Grazioli, A.; Chung, J.-Y.; Singh, M.; Yinda, C.K.; Winkler, C.W.; Sun, J.; Dickey, J.M.; Ylaya, K.; et al. SARS-CoV-2 infection and persistence in the human body and brain at autopsy. Nature 2022, 612, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.M.F.; Bhondeley, M.; Beatty, W.; Gaupp, D.D.; Doyle-Meyers, L.A.; Fischer, T.; Bandyopadhyay, I.; Blair, R.V.; Bohm, R.; Rappaport, J.; et al. SARS-CoV-2 infection of the pancreas promotes thrombofibrosis and is associated with new-onset diabetes. JCI Insight 2021, 6, e151551. [Google Scholar] [CrossRef]
- Poloni, T.E.; Moretti, M.; Medici, V.; Turturici, E.; Belli, G.; Cavriani, E.; Visonà, S.D.; Rossi, M.; Fantini, V.; Ferrari, R.R.; et al. COVID-19 Pathology in the Lung, Kidney, Heart and Brain: The Different Roles of T-Cells, Macrophages, and Microthrombosis. Cells 2022, 11, 3124. [Google Scholar] [CrossRef]
- Menter, T.; Haslbauer, J.D.; Nienhold, R.; Savic, S.; Hopfer, H.; Deigendesch, N.; Frank, S.; Turek, D.; Willi, N.; Pargger, H.; et al. Postmortem examination of COVID-19 patients reveals diffuse alveolar damage with severe capillary congestion and variegated findings in lungs and other organs suggesting vascular dysfunction. Histopathology 2020, 77, 198–209. [Google Scholar] [CrossRef]
- Baj, J.; Karakuła-Juchnowicz, H.; Teresiński, G.; Buszewicz, G.; Ciesielka, M.; Sitarz, R.; Forma, A.; Karakuła, K.; Flieger, W.; Portincasa, P.; et al. COVID-19: Specific and Non-Specific Clinical Manifestations and Symptoms: The Current State of Knowledge. J. Clin. Med. 2020, 9, 1753. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Sutanto, H.; Soegiarto, G. Risk of Thrombosis during and after a SARS-CoV-2 Infection: Pathogenesis, Diagnostic Approach, and Management. Hematol. Rep. 2023, 15, 225–243. [Google Scholar] [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Solomon, T. Neurological infection with SARS-CoV-2—The story so far. Nat. Rev. Neurol. 2021, 17, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Yong, S.J. Persistent Brainstem Dysfunction in Long-COVID: A Hypothesis. ACS Chem. Neurosci. 2021, 12, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, M.; Dong, Y.; Zhou, H.; Zhang, Z.; Tian, C.; Qin, R.; Wang, H.; Shen, Y.; Du, K.; et al. Diabetes is a risk factor for the progression and prognosis of COVID-19. Diabetes/Metab. Res. Rev. 2020, 36, e3319. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Gupta, R.; Ghosh, A.; Misra, A. Diabetes in COVID-19: Prevalence, pathophysiology, prognosis and practical considerations. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 303–310. [Google Scholar] [CrossRef]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; The Northwell COVID-19 Research Consortium. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef]
- Barron, E.; Bakhai, C.; Kar, P.; Weaver, A.; Bradley, D.; Ismail, H.; Knighton, P.; Holman, N.; Khunti, K.; Sattar, N.; et al. Associations of type 1 and type 2 diabetes with COVID-19-related mortality in England: A whole-population study. Lancet Diabetes Endocrinol. 2020, 8, 813–822. [Google Scholar] [CrossRef]
- van den Berg, J.M.; Remmelzwaal, S.; Blom, M.T.; van Hoek, B.A.C.E.; Swart, K.M.A.; Overbeek, J.A.; Burchell, G.L.; Herings, R.M.C.; Elders, P.J.M. Effectiveness of COVID-19 Vaccines in Adults with Diabetes Mellitus: A Systematic Review. Vaccines 2023, 11, 24. [Google Scholar] [CrossRef]
- Sourij, C.; Tripolt, N.J.; Aziz, F.; Aberer, F.; Forstner, P.; Obermayer, A.M.; Kojzar, H.; Kleinhappl, B.; Pferschy, P.N.; Mader, J.K.; et al. Humoral immune response to COVID-19 vaccination in diabetes is age-dependent but independent of type of diabetes and glycaemic control: The prospective COVAC-DM cohort study. Diabetes Obes. Metab. 2022, 24, 849–858. [Google Scholar] [CrossRef]
- Sardu, C.; Gargiulo, G.; Esposito, G.; Paolisso, G.; Marfella, R. Impact of diabetes mellitus on clinical outcomes in patients affected by COVID-19. Cardiovasc. Diabetol. 2020, 19, 76. [Google Scholar] [CrossRef]
- Rawshani, A.; Kjölhede, E.A.; Rawshani, A.; Sattar, N.; Eeg-Olofsson, K.; Adiels, M.; Ludvigsson, J.; Lindh, M.; Gisslén, M.; Hagberg, E.; et al. Severe COVID-19 in people with type 1 and type 2 diabetes in Sweden: A nationwide retrospective cohort study. Lancet Reg. Health Eur. 2021, 4, 100105. [Google Scholar] [CrossRef]
- Wong, R.; Hall, M.; Vaddavalli, R.; Anand, A.; Arora, N.; Bramante, C.T.; Garcia, V.; Johnson, S.; Saltz, M.; Tronieri, J.S.; et al. Glycemic Control and Clinical Outcomes in U.S. Patients With COVID-19: Data From the National COVID Cohort Collaborative (N3C) Database. Diabetes Care 2022, 45, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Chubachi, S.; Namkoong, H.; Asakura, T.; Tanaka, H.; Lee, H.; Azekawa, S.; Okada, Y.; Koike, R.; Kimura, A.; et al. Clinical significance of prediabetes, undiagnosed diabetes and diagnosed diabetes on critical outcomes in COVID-19: Integrative analysis from the Japan COVID-19 task force. Diabetes Obes. Metab. 2023, 25, 144–155. [Google Scholar] [CrossRef]
- D’Onofrio, N.; Scisciola, L.; Sardu, C.; Trotta, M.C.; De Feo, M.; Maiello, C.; Mascolo, P.; De Micco, F.; Turriziani, F.; Municinò, E.; et al. Glycated ACE2 receptor in diabetes: Open door for SARS-COV-2 entry in cardiomyocyte. Cardiovasc. Diabetol. 2021, 20, 99. [Google Scholar] [CrossRef]
- Zhang, T.; Mei, Q.; Zhang, Z.; Walline, J.H.; Liu, Y.; Zhu, H.; Zhang, S. Risk for newly diagnosed diabetes after COVID-19: A systematic review and meta-analysis. BMC Med. 2022, 20, 444. [Google Scholar] [CrossRef] [PubMed]
- Kwan, A.C.; Ebinger, J.E.; Botting, P.; Navarrette, J.; Claggett, B.; Cheng, S. Association of COVID-19 Vaccination With Risk for Incident Diabetes After COVID-19 Infection. JAMA Netw. Open 2023, 6, e2255965. [Google Scholar] [CrossRef]
- Montefusco, L.; Ben Nasr, M.; D’Addio, F.; Loretelli, C.; Rossi, A.; Pastore, I.; Daniele, G.; Abdelsalam, A.; Maestroni, A.; Dell’Acqua, M.; et al. Acute and long-term disruption of glycometabolic control after SARS-CoV-2 infection. Nat. Metab. 2021, 3, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.G.; Terebuh, P.; Kaelber, D.C.; Xu, R.; Davis, P.B. SARS-CoV-2 Infection and New-Onset Type 2 Diabetes Among Pediatric Patients, 2020 to 2022. JAMA Netw. Open 2024, 7, e2439444. [Google Scholar] [CrossRef]
- Bally, K.; Ji, B.; Soni, L. COVID-19 Vaccine-Induced Latent Autoimmune Diabetes in Adults. Cureus 2023, 15, e33762. [Google Scholar] [CrossRef]
- Unnikrishnan, R.; Pradeepa, R.; Joshi, S.R.; Mohan, V. Type 2 Diabetes: Demystifying the Global Epidemic. Diabetes 2017, 66, 1432–1442. [Google Scholar] [CrossRef]
- Abel, E.D.; Gloyn, A.L.; Evans-Molina, C.; Joseph, J.J.; Misra, S.; Pajvani, U.B.; Simcox, J.; Susztak, K.; Drucker, D.J. Diabetes mellitus2014;Progress and opportunities in the evolving epidemic. Cell 2024, 187, 3789–3820. [Google Scholar] [CrossRef]
- Murphy, S.L.K.; Kenneth, D.; Xu, J.; Arias, E. Mortality in the United States, 2023; National Center Health Statistics: Hyattsville, MA, USA, 2024; pp. 1–13. [Google Scholar]
- Gwira, J.A.; Fryar, C.D.; Gu, Q. Prevalence of Total, Diagnosed, and Undiagnosed Diabetes in Adults: United States, August 2021–August 2023; NCHS Data Brief: Hyattsville, MD, USA, 2024. [Google Scholar]
- The Diabetes Control and Complications Trial Research Group. The Effect of Intensive Treatment of Diabetes on the Development and Progression of Long-Term Complications in Insulin-Dependent Diabetes Mellitus. N. Engl. J. Med. 1993, 329, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Laakso, M. Hyperglycemia and cardiovascular disease in type 2 diabetes. Diabetes 1999, 48, 937–942. [Google Scholar] [CrossRef]
- Giri, B.; Dey, S.; Das, T.; Sarkar, M.; Banerjee, J.; Dash, S.K. Chronic hyperglycemia mediated physiological alteration and metabolic distortion leads to organ dysfunction, infection, cancer progression and other pathophysiological consequences: An update on glucose toxicity. Biomed. Pharmacother. 2018, 107, 306–328. [Google Scholar] [CrossRef] [PubMed]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Kulcsar, K.A.; Coleman, C.M.; Beck, S.E.; Frieman, M.B. Comorbid diabetes results in immune dysregulation and enhanced disease severity following MERS-CoV infection. JCI Insight 2019, 4, e131774. [Google Scholar] [CrossRef]
- Chan, J.W.M.; Ng, C.K.; Chan, Y.H.; Mok, T.Y.W.; Lee, S.; Chu, S.Y.Y.; Law, W.L.; Lee, M.P.; Li, P.C.K. Short term outcome and risk factors for adverse clinical outcomes in adults with severe acute respiratory syndrome (SARS). Thorax 2003, 58, 686–689. [Google Scholar] [CrossRef]
- Yang, J.K.; Feng, Y.; Yuan, M.Y.; Yuan, S.Y.; Fu, H.J.; Wu, B.Y.; Sun, G.Z.; Yang, G.R.; Zhang, X.L.; Wang, L.; et al. Plasma glucose levels and diabetes are independent predictors for mortality and morbidity in patients with SARS. Diabet. Med. 2006, 23, 623–628. [Google Scholar] [CrossRef]
- Yang, J.K.; Lin, S.S.; Ji, X.J.; Guo, L.M. Binding of SARS coronavirus to its receptor damages islets and causes acute diabetes. Acta Diabetol. 2010, 47, 193–199. [Google Scholar] [CrossRef]
- Alraddadi, B.M.; Watson, J.T.; Almarashi, A.; Abedi, G.R.; Turkistani, A.; Sadran, M.; Housa, A.; Almazroa, M.A.; Alraihan, N.; Banjar, A.; et al. Risk Factors for Primary Middle East Respiratory Syndrome Coronavirus Illness in Humans, Saudi Arabia, 2014. Emerg. Infect. Dis. 2016, 22, 49–55. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; Ali El Hadi Mohamed, R.; Alanazi, M.S.; Mohamed, N.; Alrasheed, M.M.; Abanmy, N.; Alhawassi, T. Prevalence of comorbidities in cases of Middle East respiratory syndrome coronavirus: A retrospective study. Epidemiol. Infect. 2018, 147, e35. [Google Scholar] [CrossRef]
- O’Meara, T.R.; Nanishi, E.; McGrath, M.E.; Barman, S.; Dong, D.; Dillen, C.; Menon, M.; Seo, H.-S.; Dhe-Paganon, S.; Ernst, R.K.; et al. Reduced SARS-CoV-2 mRNA vaccine immunogenicity and protection in mice with diet-induced obesity and insulin resistance. J. Allergy Clin. Immunol. 2023, 152, 1107–1120.e1106. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Berg, A.H.; Iyengar, P.; Lam, T.K.T.; Giacca, A.; Combs, T.P.; Rajala, M.W.; Du, X.; Rollman, B.; Li, W.; et al. The Hyperglycemia-induced Inflammatory Response in Adipocytes: The role of reactive oxyegen species. J. Biol. Chem. 2005, 280, 4617–4626. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.B. The complex role of adipokines in obesity, inflammation, and autoimmunity. Clin. Sci. 2021, 135, 731–752. [Google Scholar] [CrossRef]
- Leal, V.d.O.; Mafra, D. Adipokines in obesity. Clin. Chim. Acta 2013, 419, 87–94. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef]
- Angriman, F.; Ferreyro, B.L.; Burry, L.; Fan, E.; Ferguson, N.D.; Husain, S.; Keshavjee, S.H.; Lupia, E.; Munshi, L.; Renzi, S.; et al. Interleukin-6 receptor blockade in patients with COVID-19: Placing clinical trials into context. Lancet Respir. Med. 2021, 9, 655–664. [Google Scholar] [CrossRef]
- Rask-Madsen, C.; King, G.L. Vascular complications of diabetes: Mechanisms of injury and protective factors. Cell. Metab. 2013, 17, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Liu, S.; Gao, M.; Wang, W.; Chen, K.; Huang, L.; Liu, Y. Diabetic vascular diseases: Molecular mechanisms and therapeutic strategies. Signal Transduct. Target. Ther. 2023, 8, 152. [Google Scholar] [CrossRef]
- Mameli, C.; Ghezzi, M.; Mari, A.; Cammi, G.; Macedoni, M.; Redaelli, F.C.; Calcaterra, V.; Zuccotti, G.; D’Auria, E. The Diabetic Lung: Insights into Pulmonary Changes in Children and Adolescents with Type 1 Diabetes. Metabolites 2021, 11, 69. [Google Scholar] [CrossRef]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479–480, 609–618. [Google Scholar] [CrossRef]
- Smallwood, H.S.; Duan, S.; Morfouace, M.; Rezinciuc, S.; Shulkin, B.L.; Shelat, A.; Zink, E.E.; Milasta, S.; Bajracharya, R.; Oluwaseum, A.J.; et al. Targeting Metabolic Reprogramming by Influenza Infection for Therapeutic Intervention. Cell Rep. 2017, 19, 1640–1653. [Google Scholar] [CrossRef] [PubMed]
- Kohio, H.P.; Adamson, A.L. Glycolytic control of vacuolar-type ATPase activity: A mechanism to regulate influenza viral infection. Virology 2013, 444, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Thaker, S.K.; Chapa, T.; Garcia, G., Jr.; Gong, D.; Schmid, E.W.; Arumugaswami, V.; Sun, R.; Christofk, H.R. Differential Metabolic Reprogramming by Zika Virus Promotes Cell Death in Human versus Mosquito Cells. Cell Metab. 2019, 29, 1206–1216.e1204. [Google Scholar] [CrossRef]
- Fontaine, K.A.; Sanchez, E.L.; Camarda, R.; Lagunoff, M. Dengue Virus Induces and Requires Glycolysis for Optimal Replication. J. Virol. 2015, 89, 2358–2366. [Google Scholar] [CrossRef] [PubMed]
- Munger, J.; Bennett, B.D.; Parikh, A.; Feng, X.-J.; McArdle, J.; Rabitz, H.A.; Shenk, T.; Rabinowitz, J.D. Systems-level metabolic flux profiling identifies fatty acid synthesis as a target for antiviral therapy. Nat. Biotechnol. 2008, 26, 1179–1186. [Google Scholar] [CrossRef]
- Pouysségur, J.; Marchiq, I.; Parks, S.K.; Durivault, J.; Ždralević, M.; Vucetic, M. ‘Warburg effect’ controls tumor growth, bacterial, viral infections and immunity—Genetic deconstruction and therapeutic perspectives. Semin. Cancer Biol. 2022, 86, 334–346. [Google Scholar] [CrossRef]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef]
- López-Ayllón, B.D.; Marin, S.; Fernández, M.F.; García-García, T.; Fernández-Rodríguez, R.; de Lucas-Rius, A.; Redondo, N.; Mendoza-García, L.; Foguet, C.; Grigas, J.; et al. Metabolic and mitochondria alterations induced by SARS-CoV-2 accessory proteins ORF3a, ORF9b, ORF9c and ORF10. J. Med. Virol. 2024, 96, e29752. [Google Scholar] [CrossRef]
- Miller, B.; Silverstein, A.; Flores, M.; Cao, K.; Kumagai, H.; Mehta, H.H.; Yen, K.; Kim, S.-J.; Cohen, P. Host mitochondrial transcriptome response to SARS-CoV-2 in multiple cell models and clinical samples. Sci. Rep. 2021, 11, 3. [Google Scholar] [CrossRef]
- Guarnieri, J.W.; Dybas, J.M.; Fazelinia, H.; Kim, M.S.; Frere, J.; Zhang, Y.; Soto Albrecht, Y.; Murdock, D.G.; Angelin, A.; Singh, L.N.; et al. Core mitochondrial genes are down-regulated during SARS-CoV-2 infection of rodent and human hosts. Sci. Transl. Med. 2023, 15, eabq1533. [Google Scholar] [CrossRef]
- Rochowski, M.T.; Jayathilake, K.; Balcerak, J.-M.; Selvan, M.T.; Gunasekara, S.; Miller, C.; Rudd, J.M.; Lacombe, V.A. Impact of Delta SARS-CoV-2 Infection on Glucose Metabolism: Insights on Host Metabolism and Virus Crosstalk in a Feline Model. Viruses 2024, 16, 295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, R.; Kim, S.H.; Shah, H.; Zhang, S.; Liang, J.H.; Fang, Y.; Gentili, M.; Leary, C.N.O.; Elledge, S.J.; et al. SARS-CoV-2 hijacks folate and one-carbon metabolism for viral replication. Nat. Commun. 2021, 12, 1676. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Liu, W.; Li, X.; Zhao, P.; Shereen, M.A.; Zhu, C.; Huang, S.; Liu, S.; Yu, X.; Yue, M.; et al. HIF-1α promotes SARS-CoV-2 infection and aggravates inflammatory responses to COVID-19. Signal Transduct. Target. Ther. 2021, 6, 308. [Google Scholar] [CrossRef]
- Morris, D.R.; Qu, Y.; Agrawal, A.; Garofalo, R.P.; Casola, A. HIF-1α Modulates Core Metabolism and Virus Replication in Primary Airway Epithelial Cells Infected with Respiratory Syncytial Virus. Viruses 2020, 12, 1088. [Google Scholar] [CrossRef] [PubMed]
- Codo, A.C.; Davanzo, G.G.; Monteiro, L.d.B.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 437–446.e435. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. Natl. Sci. Rev. 2020, 7, 998–1002. [Google Scholar] [CrossRef]
- Park, J.; Dean, L.S.; Jiyarom, B.; Gangcuangco, L.M.; Shah, P.; Awamura, T.; Ching, L.L.; Nerurkar, V.R.; Chow, D.C.; Igno, F.; et al. Elevated circulating monocytes and monocyte activation in COVID-19 convalescent individuals. Front. Immunol. 2023, 14, 1151780. [Google Scholar] [CrossRef]
- Duan, X.; Tang, X.; Nair, M.S.; Zhang, T.; Qiu, Y.; Zhang, W.; Wang, P.; Huang, Y.; Xiang, J.; Wang, H.; et al. An airway organoid-based screen identifies a role for the HIF1α-glycolysis axis in SARS-CoV-2 infection. Cell Rep. 2021, 37, 109920. [Google Scholar] [CrossRef]
- Deng, W.; Bao, L.; Song, Z.; Zhang, L.; Yu, P.; Xu, Y.; Wang, J.; Zhao, W.; Zhang, X.; Han, Y.; et al. Infection with SARS-CoV-2 can cause pancreatic impairment. Signal Transduct. Target. Ther. 2024, 9, 98. [Google Scholar] [CrossRef]
- Wu, C.-T.; Lidsky, P.V.; Xiao, Y.; Lee, I.T.; Cheng, R.; Nakayama, T.; Jiang, S.; Demeter, J.; Bevacqua, R.J.; Chang, C.A.; et al. SARS-CoV-2 infects human pancreatic β cells and elicits β cell impairment. Cell Metab. 2021, 33, 1565–1576.e1565. [Google Scholar] [CrossRef]
- Hollstein, T.; Schulte, D.M.; Schulz, J.; Glück, A.; Ziegler, A.G.; Bonifacio, E.; Wendorff, M.; Franke, A.; Schreiber, S.; Bornstein, S.R.; et al. Autoantibody-negative insulin-dependent diabetes mellitus after SARS-CoV-2 infection: A case report. Nat. Metab. 2020, 2, 1021–1024. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Calvo, T.; Sabouri, S.; Anquetil, F.; von Herrath, M.G. The viral paradigm in type 1 diabetes: Who are the main suspects? Autoimmun. Rev. 2016, 15, 964–969. [Google Scholar] [CrossRef]
- Huang, J.; Pearson, J.A.; Wong, F.S.; Wen, L.; Zhou, Z. Innate immunity in latent autoimmune diabetes in adults. Diabetes/Metab. Res. Rev. 2022, 38, e3480. [Google Scholar] [CrossRef] [PubMed]
- Lui, D.T.W.; Lee, K.K.; Lee, C.H.; Lee, A.C.H.; Hung, I.F.N.; Tan, K.C.B. Development of Graves’ Disease After SARS-CoV-2 mRNA Vaccination: A Case Report and Literature Review. Front. Public Health 2021, 9, 778964. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; Moore, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef]
- Huang, Q.; An, R.; Wang, H.; Yang, Y.; Tang, C.; Wang, J.; Yu, W.; Zhou, Y.; Zhang, Y.; Wu, D.; et al. Aggravated pneumonia and diabetes in SARS-CoV-2 infected diabetic mice. Emerg. Microbes Infect. 2023, 12, 2203782. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, N.; Lu, X.; Zhou, M.; Yan, X.; Gu, W.; Yang, J.; Zhang, Q.; Zhang, C.; Gong, Y.; et al. Anti-infection effects of heparin on SARS-CoV-2 in a diabetic mouse model. Zool. Res. 2023, 44, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhao, M.-M.; Li, M.-J.; Li, X.-Y.; Jin, J.-M.; Feng, Y.-M.; Zhang, L.; Huang, W.J.; Yang, F.; Yang, J.-K. Hyperglycemia induced cathepsin L maturation linked to diabetic comorbidities and COVID-19 mortality. eLife 2024, 13, RP92826. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Ardanuy, J.; Hammond, H.; Logue, J.; Jackson, L.; Baracco, L.; McGrath, M.; Dillen, C.; Patel, N.; Smith, G.; et al. Diet-induced obesity and diabetes enhance mortality and reduce vaccine efficacy for SARS-CoV-2. J. Virol. 2023, 97, e0133623. [Google Scholar] [CrossRef]
- Burnett, F.N.; Coucha, M.; Bolduc, D.R.; Hermanns, V.C.; Heath, S.P.; Abdelghani, M.; Macias-Moriarity, L.Z.; Abdelsaid, M. SARS-CoV-2 Spike Protein Intensifies Cerebrovascular Complications in Diabetic hACE2 Mice through RAAS and TLR Signaling Activation. Int. J. Mol. Sci. 2023, 24, 6394. [Google Scholar] [CrossRef]
- Atef, Y.; Ito, T.; Masuda, A.; Kato, Y.; Nishimura, A.; Kanda, Y.; Kunisawa, J.; Kusakabe, T.; Nishida, M. Diabetic Mice Spleen Vulnerability Contributes to Decreased Persistence of Antibody Production after SARS-CoV-2 Vaccine. Int. J. Mol. Sci. 2024, 25, 10379. [Google Scholar] [CrossRef] [PubMed]
- Alkhawaldeh, O.; Jarrar, Y.; Gharaibeh, M.; Abudahab, S.; Abulebdah, D.; Jarrar, B. Alterations in the gene expression of SARS-COV-2 entry receptors and enzymes in lungs and hearts of controlled and uncontrolled diabetic mice. Fundam. Clin. Pharmacol. 2024, 38, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, Y.; Nakayama, S.; Yamamoto, A.; Kitazawa, T. High D-glucose levels induce ACE2 expression via GLUT1 in human airway epithelial cell line Calu-3. BMC Mol. Cell Biol. 2022, 23, 29. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Rodriguez, J.R.; Valdés Aguayo, J.J.; Garza-Veloz, I.; Martinez-Rendon, J.; Del Refugio Rocha Pizaña, M.; Cabral-Pacheco, G.A.; Juárez-Alcalá, V.; Martinez-Fierro, M.L. Sustained Hyperglycemia and Its Relationship with the Outcome of Hospitalized Patients with Severe COVID-19: Potential Role of ACE2 Upregulation. J. Pers. Med. 2022, 12, 805. [Google Scholar] [CrossRef]
- Baristaite, G.; Gurwitz, D. d-Galactose treatment increases and reduces mRNA expression in A549 human lung epithelial cells. Drug Dev. Res. 2022, 83, 622–627. [Google Scholar] [CrossRef]
- Garreta, E.; Prado, P.; Stanifer, M.L.; Monteil, V.; Marco, A.; Ullate-Agote, A.; Moya-Rull, D.; Vilas-Zornoza, A.; Tarantino, C.; Romero, J.P.; et al. A diabetic milieu increases ACE2 expression and cellular susceptibility to SARS-CoV-2 infections in human kidney organoids and patient cells. Cell Metab. 2022, 34, 857–873.e859. [Google Scholar] [CrossRef]
- Bülow, R.D.; Boor, P. Extracellular Matrix in Kidney Fibrosis: More Than Just a Scaffold. J. Histochem. Cytochem. 2019, 67, 643–661. [Google Scholar] [CrossRef]
- Yang, Y.-F.; Li, T.-C.; Li, C.-I.; Liu, C.-S.; Lin, W.-Y.; Yang, S.-Y.; Chiang, J.-H.; Huang, C.-C.; Sung, F.-C.; Lin, C.-C. Visit-to-Visit Glucose Variability Predicts the Development of End-Stage Renal Disease in Type 2 Diabetes: 10-Year Follow-Up of Taiwan Diabetes Study. Medicine 2015, 94, e1804. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).