
Genomic Diversity of the tet(X)-Positive Myroides Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Bacterial Isolation
2.2. Antibiotic Resistance Evaluation
2.3. Whole Genome Sequencing (WGS)
2.4. Bioinformatics Analyses
2.5. Phenotypic Experiments of Novel Myroides Species
2.6. Cloning Expression
2.7. Transfer of tet(X) Genes
3. Results
3.1. Sporadic Detection of tet(X)-Positive MDR Myroides spp.
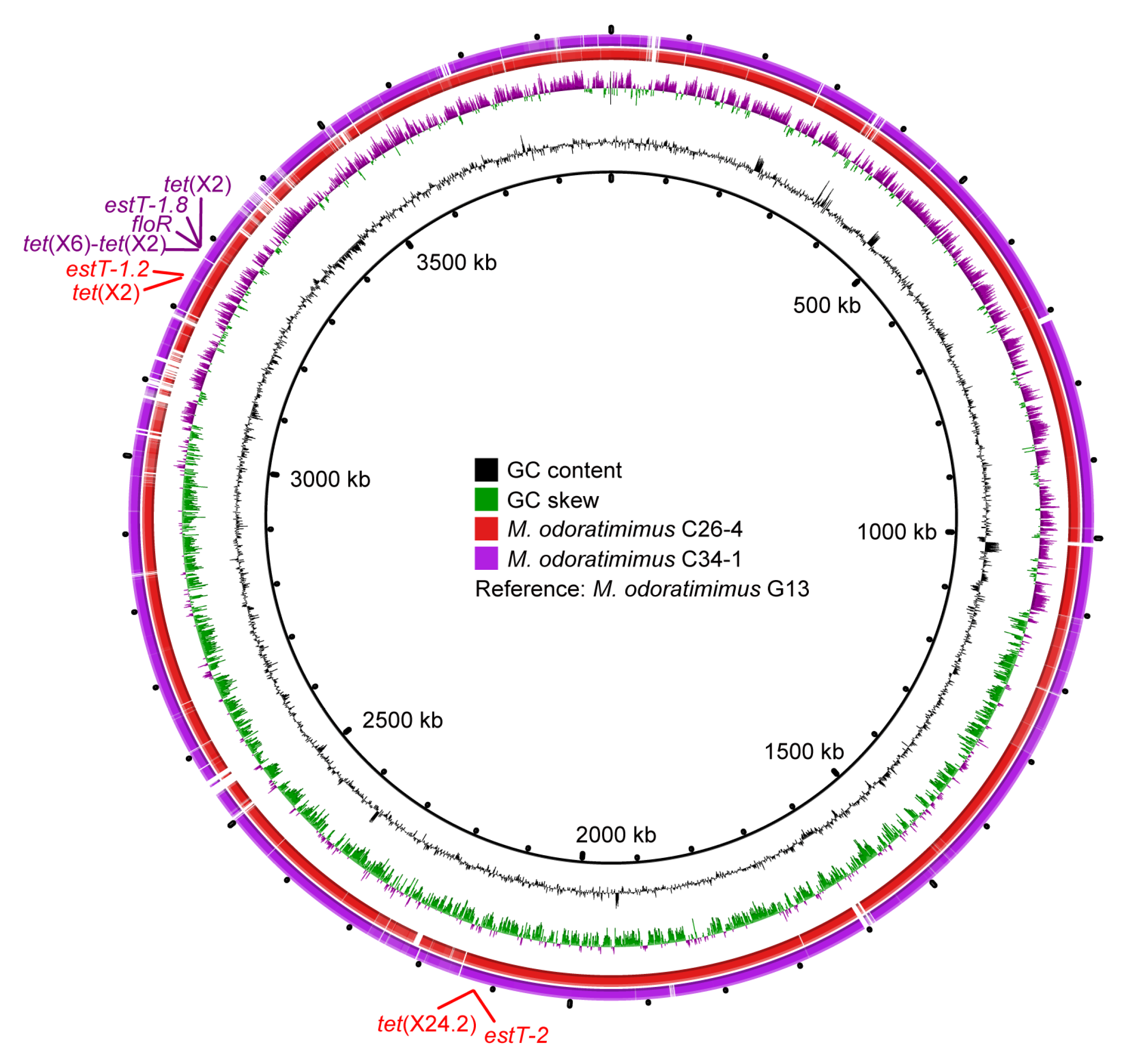
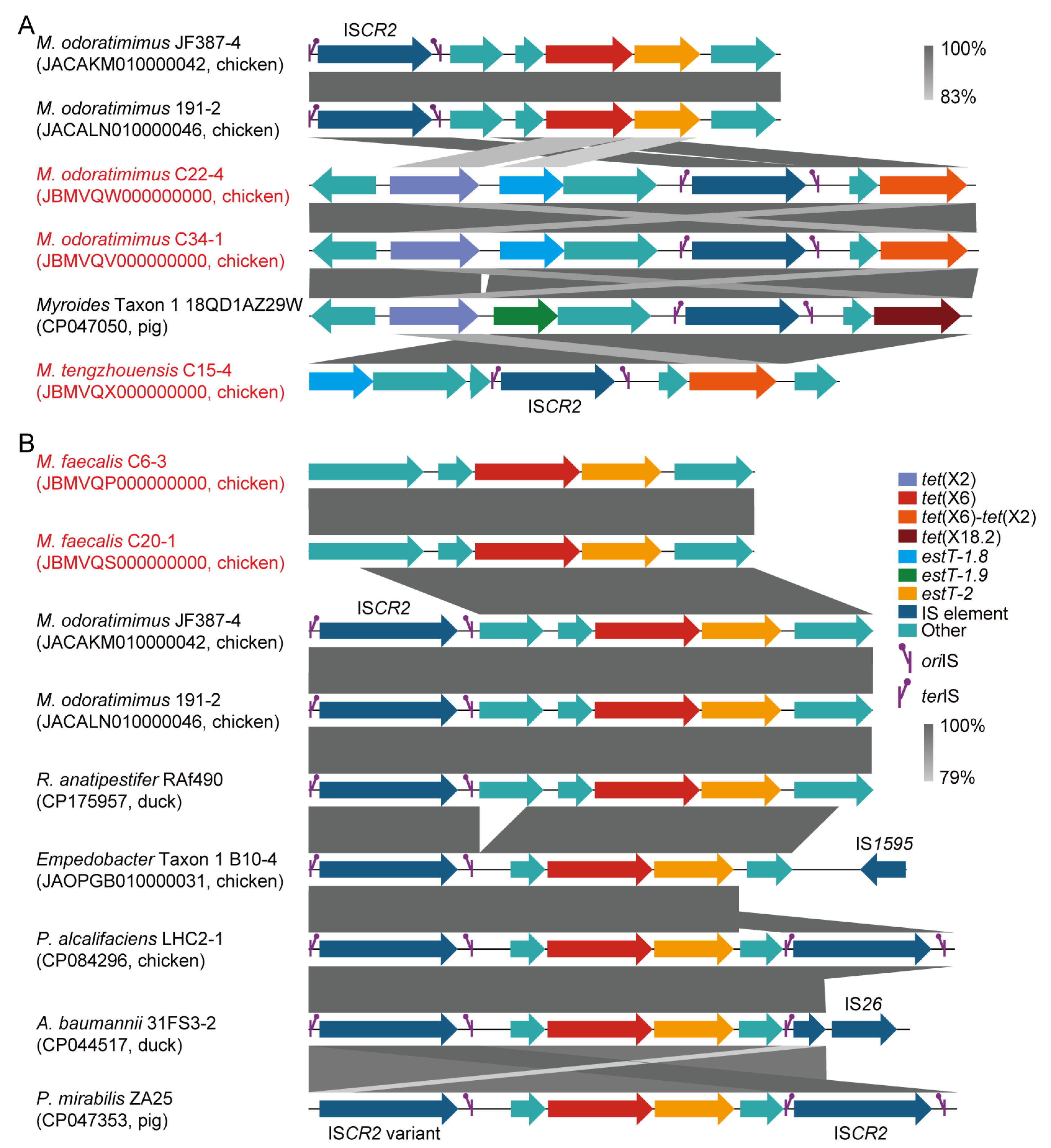
3.2. Polymorphism of tet(X) and estT
3.3. Novel Myroides Species
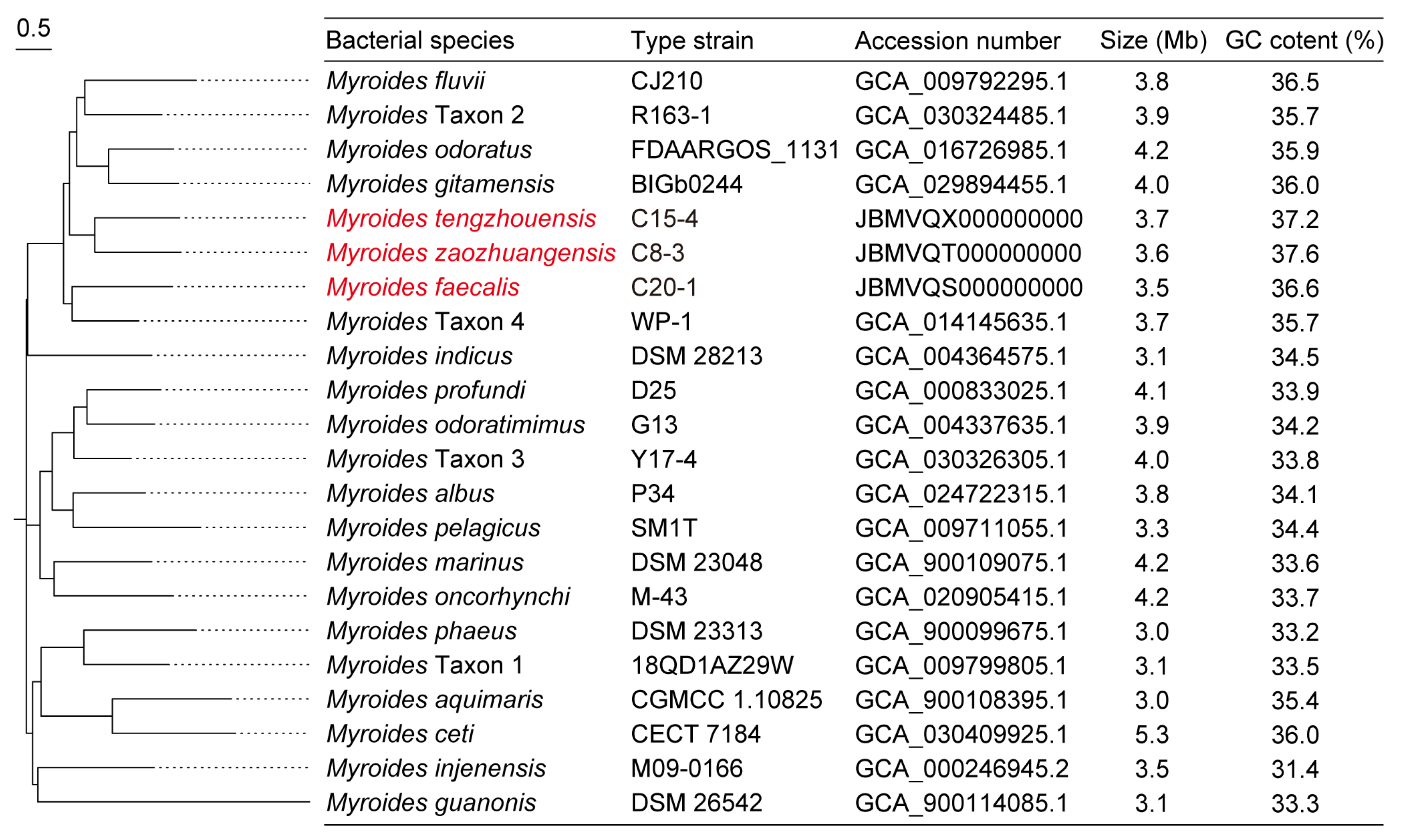
3.3.1. Description of Myroides zaozhuangensis sp. nov.
3.3.2. Description of Myroides tengzhouensis sp. nov.
3.3.3. Description of Myroides faecalis sp. nov.
3.4. Phylogeny of Myroides spp.
3.5. ISCR2-Mediated Transposons of tet(X) and estT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDR | Multidrug-resistant |
| MICs | Minimum inhibitory concentrations |
| LB | Luria–Bertani |
| MH | Mueller–Hinton |
| WGS | Whole genome sequencing |
| ANI | Average nucleotide identity |
| SNP | Single-nucleotide polymorphism |
| isDDH | in silico DNA-DNA hybridization |
| ARGs | Antibiotic resistance genes |
| ERIC-PCR | Enterobacterial repetitive intergenic consensus PCR |
| IS | Insertion sequence |
References
- LPSN. Available online: https://lpsn.dsmz.de/genus/myroides (accessed on 10 May 2025).
- Gunzer, F.; Rudolph, W.W.; Bunk, B.; Schober, I.; Peters, S.; Muller, T.; Oberheitmann, B.; Schrottner, P. Whole-genome sequencing of a large collection of Myroides odoratimimus and Myroides odoratus isolates and antimicrobial susceptibility studies. Emerg. Microbes Infect. 2018, 7, 61. [Google Scholar] [CrossRef]
- Aworh, M.K.; Colín-Castro, C.A.; Ortiz-Álvarez, J.M.; Hernández-Pérez, C.F.; Hernández-Durán, M.; García-Hernández, M.d.L.; Martínez-Zavaleta, M.G.; Becerra-Lobato, N.; Cervantes-Hernández, M.I.; Rosas-Alquicira, G.; et al. Myroides species, pathogenic spectrum and clinical microbiology sight in Mexican isolates. PLoS ONE 2024, 19, e0310262. [Google Scholar] [CrossRef]
- Kurt, A.F.; Mete, B.; Houssein, F.M.; Tok, Y.; Kuskucu, M.A.; Yucebag, E.; Urkmez, S.; Tabak, F.; Aygun, G. A pan-resistant Myroides odoratimimus catheter-related bacteremia in a COVID-19 patient and review of the literature. Acta Microbiol. Immunol. Hung. 2022, 69, 164–170. [Google Scholar] [CrossRef]
- Sahu, C.; Patel, S.S.; Chaudhary, R.; Bhartiya, C.; Bhatnagar, N. A Retrospective Study on UTI by Myroides Species: An Emerging Drug Resistant Nosocomial Pathogen. Indian J. Crit. Care M 2024, 28, 399–403. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, M.; Fu, J.; Zhong, C.; Zong, G.; Cao, G. Identification of a mobilizable, multidrug-resistant genomic island in Myroides odoratimimus isolated from Tibetan pasture. Sci. Total Environ. 2020, 723, 137970. [Google Scholar] [CrossRef]
- Aygar, I.S.; Aydogan, C.N.; Ozcan, H.; Unat, I.; Fatsa, T.; Tekin, K.; Yalci, A.; Hosbul, T.; Sahiner, F.; Gumral, R. Myroides odoratimimus: A New Threat with Persistent Infections, Multidrug Resistance, and the Potential for Hospital Outbreaks. Jpn. J. Infect. Dis. 2023, 76, 335–342. [Google Scholar] [CrossRef]
- Yartasi, E.; Durmaz, R.; Ari, O.; Mumcuoglu, I.; Dinc, B. Molecular characterization of the multi-drug resistant Myroides odoratimimus isolates: A whole genome sequence-based study to confirm carbapenem resistance. Int. Microbiol. 2023, 27, 1169–1180. [Google Scholar] [CrossRef]
- Seifert, H.; Blondeau, J.; Lucassen, K.; Utt, E.A. Global update on the in vitro activity of tigecycline and comparators against isolates of Acinetobacter baumannii and rates of resistant phenotypes (2016–2018). J. Glob. Antimicrob. Res. 2022, 31, 82–89. [Google Scholar] [CrossRef]
- Nguyen, F.; Starosta, A.L.; Arenz, S.; Sohmen, D.; Donhofer, A.; Wilson, D.N. Tetracycline antibiotics and resistance mechanisms. Biol. Chem. 2014, 395, 559–575. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Huband, M.D.; Streit, J.M.; Flamm, R.K.; Sader, H.S. Surveillance of tigecycline activity tested against clinical isolates from a global (North America, Europe, Latin America and Asia-Pacific) collection (2016). Int. J. Antimicrob. Agents 2018, 51, 848–853. [Google Scholar] [CrossRef]
- Chen, C.; Cui, C.Y.; Wu, X.T.; Fang, L.X.; He, Q.; He, B.; Long, T.F.; Liao, X.P.; Chen, L.; Liu, Y.H.; et al. Spread of tet(X5) and tet(X6) genes in multidrug-resistant Acinetobacter baumannii strains of animal origin. Vet. Microbiol. 2021, 253, 108954. [Google Scholar] [CrossRef]
- He, T.; Wang, R.; Liu, D.J.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.B.; Ji, Q.J.; Wei, R.C.; Liu, Z.H.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Sun, J.; Chen, C.; Cui, C.Y.; Zhang, Y.; Liu, X.; Cui, Z.H.; Ma, X.Y.; Feng, Y.; Fang, L.X.; Lian, X.L.; et al. Plasmid-encoded tet(X) genes that confer high-level tigecycline resistance in Escherichia coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef]
- Wang, L.; Liu, D.; Lv, Y.; Cui, L.; Li, Y.; Li, T.; Song, H.; Hao, Y.; Shen, J.; Wang, Y.; et al. Novel Plasmid-Mediated tet(X5) Gene Conferring Resistance to Tigecycline, Eravacycline, and Omadacycline in a Clinical Acinetobacter baumannii Isolate. Antimicrob. Agents Chemother. 2019, 64, e01326-19. [Google Scholar] [CrossRef]
- He, D.; Wang, L.; Zhao, S.; Liu, L.; Liu, J.; Hu, G.; Pan, Y. A novel tigecycline resistance gene, tet(X6), on an SXT/R391 integrative and conjugative element in a Proteus genomospecies 6 isolate of retail meat origin. J. Antimicrob. Chemother. 2020, 75, 1159–1164. [Google Scholar] [CrossRef]
- Li, R.; Peng, K.; Xiao, X.; Wang, Y.; Wang, Z. Characterization of novel ISAba1-bounded tet(X15)-bearing composite transposon Tn6866 in Acinetobacter variabilis. J. Antimicrob. Chemother. 2021, 76, 2481–2483. [Google Scholar] [CrossRef]
- Chen, C.; Lv, Y.; Wu, T.; Liu, J.; Guo, Y.; Huang, J. Concurrence of Inactivation Enzyme-Encoding Genes tet(X), blaEBR, and estT in Empedobacter Species from Chickens and Surrounding Environments. Foods 2024, 13, 3201. [Google Scholar] [CrossRef]
- Li, R.; Jiang, Y.; Peng, K.; Wang, Y.; Wang, M.; Liu, Y.; Wang, Z. Phenotypic and genomic analysis reveals Riemerella anatipestifer as the potential reservoir of tet(X) variants. J. Antimicrob. Chemother. 2022, 77, 374–380. [Google Scholar] [CrossRef]
- Jin, H.; Jia, Q.; Jin, X.; Zhu, X.; Wang, M.-G.; Sun, R.-Y.; Cui, C. Identification of novel Tet(X6)-Tet(X2) recombinant variant in Elizabethkingia meningoseptica from a bullfrog farm and downstream river in China. Front. Microbiol. 2024, 15, 1453801. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Peng, K.; Wang, Q.; Liu, R.; Wang, Z.; Li, R. Characterisation of a Novel Tigecycline Resistance Gene tet(X22) and its Coexistence with blaNDM-1 in a Pseudomonas caeni Isolate. Int. J. Antimicrob. Agents 2023, 62, 106961. [Google Scholar] [CrossRef]
- Liu, D.; Zhai, W.; Song, H.; Fu, Y.; Schwarz, S.; He, T.; Bai, L.; Wang, Y.; Walsh, T.R.; Shen, J. Identification of the novel tigecycline resistance gene tet(X6) and its variants in Myroides, Acinetobacter and Proteus of food animal origin. J. Antimicrob. Chemother. 2020, 75, 1428–1431. [Google Scholar] [CrossRef]
- Dong, N.; Zeng, Y.; Cai, C.; Sun, C.; Lu, J.; Liu, C.; Zhou, H.; Sun, Q.; Shu, L.; Wang, H.; et al. Prevalence, transmission, and molecular epidemiology of tet(X)-positive bacteria among humans, animals, and environmental niches in China: An epidemiological, and genomic-based study. Sci. Total Environ. 2022, 818, 151767. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Wu, J.W.; Chen, Y.Y.; Quyen, T.L.T.; Liao, W.C.; Li, S.W.; Chen, Y.C.; Pan, Y.J. An Outbreak of tet(X6)-Carrying Tigecycline-Resistant Acinetobacter baumannii Isolates with a New Capsular Type at a Hospital in Taiwan. Antibiotics 2021, 10, 1239. [Google Scholar] [CrossRef]
- Miyoshi, T.; Iwatsuki, T.; Naganuma, T. Phylogenetic characterization of 16S rRNA gene clones from deep-groundwater microorganisms that pass through 0.2-micrometer-pore-size filters. Appl. Environ. Microb. 2005, 71, 1084–1088. [Google Scholar] [CrossRef]
- CLSI. CLSI Performance Standards for Antimicrobial Susceptibility Testing Guideline M100-Ed32; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Al Atrouni, A.; Joly-Guillou, M.L.; Hamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/datasets/genome/ (accessed on 10 January 2025).
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, Y.; Fan, G.; Sun, D.; Zhang, X.; Yu, Z.; Wang, J.; Wu, L.; Shi, W.; Ma, J. IPGA: A handy integrated prokaryotes genome and pan-genome analysis web service. iMeta 2022, 1, e55. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antimicrobial Resistance or Virulence Genes. Available online: https://github.com/tseemann/abricate (accessed on 27 February 2025).
- Xu, S.; Dai, Z.; Guo, P.; Fu, X.; Liu, S.; Zhou, L.; Tang, W.; Feng, T.; Chen, M.; Zhan, L.; et al. ggtreeExtra: Compact Visualization of Richly Annotated Phylogenetic Data. Mol. Biol. Evol. 2021, 38, 4039–4042. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.M.; Schwarz, S. Resistance gene naming and numbering: Is it a new gene or not? J. Antimicrob. Chemother. 2016, 71, 569–571. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Ram, H.; Kumar, A.; Thomas, L.; Dastager, S.G.; Mawlankar, R.; Singh, V.P. Myroides indicus sp. nov., isolated from garden soil. Int. J. Syst. Evol. Microbiol. 2015, 65, 4008–4012. [Google Scholar] [CrossRef]
- Chen, C.; Cui, C.Y.; Yu, J.J.; He, Q.; Wu, X.T.; He, Y.Z.; Cui, Z.H.; Li, C.; Jia, Q.L.; Shen, X.G.; et al. Genetic diversity and characteristics of high-level tigecycline resistance Tet(X) in Acinetobacter species. Genome Med. 2020, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, H.J.; Towner, K.J.; Dijkshoorn, L.; Gernersmidt, P.; Maher, M.; Seifert, H.; Vaneechoutte, M. Multicenter Study Using Standardized Protocols and Reagents for Evaluation of Reproducibility of PCR-Based Fingerprinting of Acinetobacter spp. J. Clin. Microbiol. 1997, 35, 3071–3077. [Google Scholar] [CrossRef]
- Cui, C.Y.; Chen, C.; Liu, B.T.; He, Q.; Wu, X.T.; Sun, R.Y.; Zhang, Y.; Cui, Z.H.; Guo, W.Y.; Jia, Q.L.; et al. Co-occurrence of Plasmid-Mediated Tigecycline and Carbapenem Resistance in Acinetobacter spp. from Waterfowls and Their Neighboring Environment. Antimicrob. Agents Chemother. 2020, 64, e02502-19. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.Y.; He, Q.; Jia, Q.L.; Li, C.; Chen, C.; Wu, X.T.; Zhang, X.J.; Lin, Z.Y.; Zheng, Z.J.; Liao, X.P.; et al. Evolutionary Trajectory of the Tet(X) Family: Critical Residue Changes towards High-Level Tigecycline Resistance. mSystems 2021, 6, e00050-21. [Google Scholar] [CrossRef]
- Cheng, Q.; Cheung, Y.; Liu, C.; Chan, E.W.C.; Wong, K.Y.; Zhang, R.; Chen, S. Functional and phylogenetic analysis of TetX variants to design a new classification system. Commun. Biol. 2022, 5, 522. [Google Scholar] [CrossRef]
- Ming, D.; Chen, Q.Q.; Chen, X.T. Analysis of resistance genes in pan-resistant Myroides odoratimimus clinical strain PR63039 using whole genome sequencing. Microb. Pathog. 2017, 112, 164–170. [Google Scholar] [CrossRef]
- Speer, B.S.; Bedzyk, L.; Salyers, A.A. Evidence that a novel tetracycline resistance gene found on two Bacteroides transposons encodes an NADP-requiring oxidoreductase. J. Bacteriol. 1991, 173, 176–183. [Google Scholar] [CrossRef]
- Linkevicius, M.; Sandegren, L.; Andersson, D.I. Potential of Tetracycline Resistance Proteins To Evolve Tigecycline Resistance. Antimicrob. Agents Chemother. 2016, 60, 789–796. [Google Scholar] [CrossRef]
- Blake, K.S.; Xue, Y.-P.; Gillespie, V.J.; Fishbein, S.R.S.; Tolia, N.H.; Wencewicz, T.A.; Dantas, G. The tetracycline resistome is shaped by selection for specific resistance mechanisms by each antibiotic generation. Nat. Commun. 2025, 16, 1452. [Google Scholar] [CrossRef]
- Dhindwal, P.; Thompson, C.; Kos, D.; Planedin, K.; Jain, R.; Jelinski, M.; Ruzzini, A. A neglected and emerging antimicrobial resistance gene encodes for a serine-dependent macrolide esterase. Proc. Natl. Acad. Sci. USA 2023, 120, e2219827120. [Google Scholar] [CrossRef] [PubMed]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR elements: Novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. R 2006, 70, 296–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhan, Z.; Shi, C. International Spread of Tet(X4)-Producing Escherichia coli Isolates. Foods 2022, 11, 2010. [Google Scholar] [CrossRef]
- Chen, C.; Chen, L.; Zhang, Y.; Cui, C.Y.; Wu, X.T.; He, Q.; Liao, X.P.; Liu, Y.H.; Sun, J. Detection of chromosome-mediated tet(X4)-carrying Aeromonas caviae in a sewage sample from a chicken farm. J. Antimicrob. Chemother. 2019, 74, 3628–3630. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Strains (MICs, mg/L) 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| C8-3 | C6-3 | C34-1 | C26-4 | C22-4 | C21-3 | C18-2 | C20-1 | C15-4 | |
| TC | 32 | 128 | 64 | 128 | 64 | 64 | 64 | 64 | 32 |
| TGC | 4 | 8 | 16 | 16 | 16 | 16 | 16 | 8 | 8 |
| SAM | 2/1 | 2/1 | 32/16 | 16/8 | 32/16 | 16/8 | 32/16 | 2/1 | 16/8 |
| CAZ | 32 | 32 | 64 | 128 | 64 | 128 | 128 | 32 | 64 |
| MEM | 0.25 | 0.5 | 0.5 | 4 | 2 | 2 | 2 | 0.5 | 4 |
| CTX | 16 | 16 | 128 | 128 | 128 | 128 | 128 | 16 | 64 |
| AMK | >256 | 64 | 256 | >256 | >256 | >256 | >256 | 256 | >256 |
| GEN | 128 | 64 | >256 | 128 | >256 | >256 | 256 | 128 | 256 |
| CS | >256 | 256 | >256 | >256 | >256 | >256 | >256 | 256 | 256 |
| CIP | 32 | 64 | >64 | >64 | >64 | >64 | >64 | >64 | 8 |
| GAT | 8 | >16 | >16 | 8 | >16 | 8 | 8 | >16 | 8 |
| FFC | 32 | 32 | 128 | 64 | 32 | 32 | 64 | 32 | 32 |
| SXT | 16/304 | 8/152 | 16/304 | >16/304 | 16/304 | >16/304 | 16/304 | 8/152 | 16/304 |
| TYL | 512 | 128 | 256 | 512 | 256 | 512 | 512 | 256 | 512 |
| TIL | 64 | 32 | 1 | 64 | 8 | 64 | 64 | 32 | 1 |
| TIP | 32 | 32 | 8 | 16 | 8 | 8 | 32 | 32 | 4 |
| Clones | MICs (mg/L) 1 | ||||||
|---|---|---|---|---|---|---|---|
| TC | DOX | MIN | TGC | TYL | TIL | TIP | |
| E. coli JM109 + pBAD24: tet(X2) | 16 | 4 | 0.5 | 0.25 | - | - | - |
| E. coli JM109 + pBAD24: tet(X6) | 128 | 16 | 16 | 8 | - | - | - |
| E. coli JM109 + pBAD24: tet(X6)-tet(X2) | 128 | 16 | 16 | 4 | - | - | - |
| E. coli JM109 + pBAD24: tet(X24.2) | 128 | 32 | 16 | 8 | - | - | - |
| E. coli JM109 + pBAD24: estT-1.2 | - | - | - | - | 512 | 64 | 1 |
| E. coli JM109 + pBAD24: estT-1.8 | - | - | - | - | 512 | 64 | 2 |
| E. coli JM109 + pBAD24: estT-2 | - | - | - | - | 256 | 64 | 1 |
| E. coli JM109 + pBAD24 | 2 | 0.5 | 0.25 | 0.125 | 128 | 16 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Wu, T.; Liu, J.; Lv, Y. Genomic Diversity of the tet(X)-Positive Myroides Species. Microorganisms 2025, 13, 1180. https://doi.org/10.3390/microorganisms13061180
Chen C, Wu T, Liu J, Lv Y. Genomic Diversity of the tet(X)-Positive Myroides Species. Microorganisms. 2025; 13(6):1180. https://doi.org/10.3390/microorganisms13061180
Chicago/Turabian StyleChen, Chong, Taotao Wu, Jing Liu, and Yilin Lv. 2025. "Genomic Diversity of the tet(X)-Positive Myroides Species" Microorganisms 13, no. 6: 1180. https://doi.org/10.3390/microorganisms13061180
APA StyleChen, C., Wu, T., Liu, J., & Lv, Y. (2025). Genomic Diversity of the tet(X)-Positive Myroides Species. Microorganisms, 13(6), 1180. https://doi.org/10.3390/microorganisms13061180