Abstract
Despite the increased reporting of Carbapenem-resistant hypervirulent Klebsiella pneumoniae (CR-hvKp) in Egypt, there is a paucity of information regarding the molecular characteristics of such strains. Herein, we present the genome sequence of two CR-hvKp strains, K22 and K45, which were isolated from VAP (ventilator-associated-pneumonia) patients admitted to pediatric ICU at Assiut University Children’s Hospital, Egypt. K22 and K45 isolates were subjected to antimicrobial susceptibility testing and whole-genome sequencing. Genomic analysis was performed to characterize each strain, determining their plasmids, antimicrobial resistance (AMR) genes, and virulence determinants. K22 possessed an extensive drug resistance phenotype (XDR), whilst K45 exhibited a multidrug resistance phenotype (MDR), with genome sequencing revealing the presence of a diverse array of AMR genes. Both strains were resistant to the carbapenem antibiotic imipenem, carrying the OXA-48 carbapenemase, with K22 additionally possessing an NDM-1 carbapenemase. Each strain was considered high-risk, with K22 and K45 respectively belonging to sequence types ST383 and ST14 and possessing virulence genes implicated in hypervirulence (e.g., iucABCD-iutA and rmpA). Importantly, both strains carried multiple plasmid replicons, including an AMR/virulence IncHI1B/FIB hybrid plasmid and MDR IncL/M plasmids. This report highlights the critical role of plasmids in the evolution of virulent K. pneumoniae strains and suggests the circulation of an IncHI1B/FIB hybrid plasmid, simultaneously disseminating AMR and hypervirulence, amongst K. pneumoniae strains within Assiut University Children’s Hospital.
1. Introduction
Klebsiella pneumoniae is a Gram-negative, encapsulated bacterium that colonizes the human gut and frequently causes healthcare-associated infections [1,2]. Hypervirulent K. pneumoniae (hvKp) is a distinct pathotype first recognized in the 1980s, that evolved through the acquisition of potent virulence determinants. HvKp signaled the onset of a concerning public health threat owing to its ability to produce invasive infections even in healthy immunocompetent adults [1]. Nevertheless, hospital-acquired hvKp infections has been increasingly reported worldwide [3,4,5]. Generally, community-acquired hvKp infections result in pyogenic liver abscess, bacteremia, and pneumonia, while health care-associated hvKp infections tend to lead to ventilator-associated pneumonia (VAP), catheter-associated urinary tract infection, and surgical site infections [2]. Furthermore, hvKp strains can colonize healthy individuals, with a propensity for dissemination to otherwise healthy individuals [6].
Initially, hvKp strains were susceptible to many different classes of antibiotics. Along with the reported worldwide dissemination of antibiotic resistance, multidrug resistance (MDR) hvKp strains were increasingly reported, namely through horizontal gene transfer between classical-Kp (cKp) and hvKp strains [7]. MDR-hvKp strains can carry virulence and resistance genes on separate plasmids or harbor large hybrid plasmids, encoding both determinants [8]. Alarmingly, carbapenem-resistant hvKp strains (CR-hvKp) are now emerging, causing severe infections, coupled with restricted antibiotic options, leading to elevated morbidity and mortality [4,9]. It is postulated that CR-hvKp can evolve either from hvKp strains through acquiring carbapenemases-encoding plasmids or from carbapenem-resistant cKp by acquiring hypervirulence genes. Worryingly, the presence of a hybrid plasmid carrying both carbapenemase and virulence genes can cause rapid evolution of CR-hvKp from carbapenem susceptible cKP through a single conjugation event [10].
Recently, CR-hvKp has been reported in many regions of Egypt [11,12]. Despite this, there are only a few studies investigating the genomic characteristics of CR-hvKp [11,13,14]. This limited number of studies may be attributed to the relatively recent recognition of CR-hvKp in Egypt, the cost of genomic characterization, and the lack of routine screening for virulence factors in hospitals. Our previous study demonstrated the predominance of MDR-K. pneumoniae as the causative agent of VAP in a pediatric ICU (intensive care unit) [12]. A subsequent investigation, involving 53 hvKP isolated from VAP patients admitted to different ICUs at Assiut University Hospitals, revealed that similar PFGE hvKp clones were isolated from diverse ICUs at different times suggesting the likelihood of long-term colonization, as well as the dissemination of hvKp in the hospital environment. On the other hand, different PFGE hvKp clones were found to carry similar virulence and resistance genes, suggesting the presence of endemic plasmids circulating in the hospital environment [11,12]. Thus, an in-depth analysis of plasmids encoding carbapenemase and virulence determinants may provide insight into the evolution of MDR hvKp strains in Egypt and, importantly, provide aid in their control.
Accordingly, in this study, we analyzed the phenotypic and genomic characteristics (i.e., the plasmids, virulence-associated genes, and AMR genes) of two CR-hvKp strains, K22 and K45. Both strains were isolated at the pediatric ICU of Assiut University Hospital, Egypt, in 2015 from patients suffering from VAP. The detection of CR-hvKp, underlines the threat posed by such pathogens in ICU settings and supports the need for strengthened surveillance and more effective screening procedures to identify and isolate hvKp strains.
2. Materials and Methods
2.1. Bacterial Isolates
The current study is a retrospective study analyzing two clinical CR-hvKP strains (K22 and K45) isolated from the pediatric ICU at Assiut University Hospital, Egypt. Both strains were retrieved from endotracheal aspirates of patients suffering from VAP who were enrolled in a previous study [12]. Strains were categorized as hvKP based on the detection of virulence genes iucA, rmpA, rmpA2, iroN, and iroB [1,2]. They were identified as carbapenem-resistant by determining the minimum inhibitory concentrations (MICs) of imipenem using an E-test. The MIC for ciprofloxacin was also done using an E-test (BioMérieux, Solna, Sweden) and was interpreted according to the Clinical and Laboratory Standard Institute (CLSI) [15].
2.2. Antimicrobial Susceptibility Testing
Antimicrobial susceptibility testing was determined through the Kirby–Bauer disk diffusion method [16] using antibiotic disks (ThermoFisher Scientific, Waltham, MA, USA) representing different classes of antibiotics, including penicillin derivatives [amoxicillin (AML10 μg), amoxicillin/clavulanic acid (AMC 20/10 μg), piperacillins (PI 100 μg)], cephalosporins [cefazolin (CZ 30 μg), cefpodoxime (CPD 30 μg), cefoperazone (CPZ 75 μg), ceftriaxone (CTR 30 μg)], aminoglycosides [gentamicin (GE 10 μg), amikacin (AK 30 μg)], tetracycline (TE 30 μg), chloramphenicol (C 30 μg), and trimethoprim sulfonamide (SXT 1.25/23.75 μg). The results were interpreted according to the guidelines stated by CLSI 2023 [15].
2.3. String Test
The string test was performed for the detection of the hypermucoviscous phenotype using a standard bacteriologic loop (Fisher Scientific, Loughborough, UK) to generate a viscous string from a single bacterial colony. Strains with a string >5 mm were considered positive for the string test [17].
2.4. Whole-Genome Sequencing of Clinical CR-hvKP Strains
The genome sequencing of the two CR-hvKP strains was performed using Illumina sequencing at Microbes NG (https://microbesng.com/ (accessed on 25 April 2025)). Bacterial cultures were preserved in a DNA/RNA Shield cryopreservative (Zymo Research, Irvine, CA, USA) and 45 µL of cell suspension was lysed with 120 μL of TE buffer, containing lysozyme (MPBio, Santa Ana, CA, USA), metapolyzyme (Sigma Aldrich, St. Louis, MO, USA) and RNase A (ITW Reagents, Barcelona, Spain), and incubated for 25 min at 37 °C. Proteinase K (VWR Chemicals, OH, USA) (final concentration 0.1 mg/mL) and SDS (Sigma-Aldrich, St. Louis, MO, USA) (final concentration 0.5% v/v) were added and incubated for 5 min at 65 °C. Genomic DNA was purified using an equal volume of SPRI beads and resuspended in EB buffer (10 mM Tris-HCl, pH 8.0). Extracted DNA was then quantified with the Quant-iT dsDNA HS (ThermoFisher Scientific, Waltham, MA, USA) assay in an Eppendorf AF2200 plate reader (Eppendorf UK Ltd, Hamburg, UK) and diluted as appropriate.
Genomic DNA libraries were prepared using the Nextera XT Library Prep Kit (Illumina, San Diego, CA, USA), following the manufacturer’s instructions. DNA quantification and library preparations were carried out on a Hamilton Microlab STAR automated liquid handling system (Hamilton Bonaduz AG, Bonaduz, Switzerland). Sequencing was performed with the Illumina NovaSeq 600 (Illumina, San Diego, CA, USA), using a 250 bp paired-end protocol. Adapter trimming of Illumina reads was done using Trimmomatic 0.30, with a sliding window quality cutoff of Q15 [18], producing 1,226,225 trimmed reads for K22 and 786,858 for K45. Genome assembly was achieved using Spades version 3.7 [19] and contigs were annotated using Prokka 1.11 [20]. This Whole Genome Shotgun project has been deposited at the National Center for Biotechnology Information (NCBI) GenBank as BioProject PRJNA1079765, with the sequence data for clinical CR-hvKP (K22 and K45) under accession numbers JBAMJT000000000 and JBAMJS000000000, respectively.
2.5. Genomic Analysis
Draft genomes were visualized using Artemis [21], and comparisons between genomes were examined using the Proksee website (https://proksee.ca/about (accessed on 25 April 2025)) [22], the Basic Local Alignment Search Tool (BLAST) at NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 25 April 2025)) and the Artemis Comparison Tool (ACT) [23]. Representations of genome organization were drawn using the Proksee Server [22] and ACT [23]. Sequence types were determined using MLST 2.0 [24], and the presence of plasmids was determined by detecting plasmid replicons using PlasmidFinder 2.1 [25] with the online software from the Center for Genomic Epidemiology (CGE) (http://www.genomicepidemiology.org/ (accessed on 25 April 2025)). Antibiotic resistance genes were detected using ResFinder 3.2 at CGE [26] and using the ResFinder (22 March 2024) and PointFinder (8 March 2024) databases at the standard setting of 90% and 60% for threshold and length ID, respectively. Resistance gene identity was further corroborated by interrogating the Prokka annotation of each isolate [20], BLAST analysis at NCBI and the Comprehensive Antibiotic Resistance Database (CARD) using Proksee [22,27]. Kaptive Web (https://kaptive-web.erc.monash.edu/ (accessed on 25 April 2025)) was used for the determination of capsule type and O-antigen locus type [28]. Furthermore, heavy metal-resistance genes were detected using the BIGSdb-Kp database at the Institut Pasteur (https://bigsdb.web.pasteur.fr/index.html (accessed on 25 April 2025)). The Virulence Factor Database (http://www.mgc.ac.cn/VFs/main.htm (accessed on 25 April 2025)) was used to screen for virulence genes [29]. Insertion sequences and bacteriophage were identified using ISfinder (https://www-is.biotoul.fr/index.php (accessed on 25 April 2025)) [30] and PHASTER (https://phaster.ca/ (accessed on 25 April 2025)) [31], respectively.
3. Results
3.1. Bacterial Isolation and AMR
The two clinical CR-hvKP isolates, K22 and K45, were isolated from children with VAP admitted to the pediatric ICU at Assiut University Children’s Hospital in 2015 [12]. Phenotypic analysis indicated that these two isolates were highly resistant to almost all tested antibiotics with both isolates resistant to the front-line carbapenem antibiotic, imipenem (Table 1). Thus, K22 is classified as possessing extensive drug resistance phenotype (XDR), while K45 exhibited an MDR phenotype [32]. However, it is of note that both isolates were still susceptible to tetracycline and chloramphenicol, with K45 additionally susceptible to fluoroquinolones. Furthermore, both strains were negative for the string test, which is used to define the hypermucoviscous phenotype (Table 1) [17].

Table 1.
Characterization and antimicrobial susceptibility testing of clinical CR-hvKP isolates.
3.2. Genomic Characterization of CR-hvKP Isolates
To understand more about each isolate, the genomes of K22 and K45 were sequenced using Illumina short-read sequencing, which revealed that they possessed respective genome sizes of 5,862,489 bp and 5,741,936 bp, with both carrying multiple plasmid replicons (Table 2). Comparison of each strain’s genome with the chromosome of the archetype K. pneumoniae reference strain, MGH 78587 (CP000647.1) [33], revealed that both differed from MGH 78578 in their capsule and O-antigen locus, along with other chromosomal positions, which included bacteriophage, sugar utilization operons and secretions systems (Figure 1). Both isolates possessed an O1v1 O-antigen loci, whilst K22 was capsular type K30 and K45 capsular type K2 (Table 2). Note that K2 capsule is often associated with hypervirulence in K. pneumoniae [34]. Sequence typing indicated that K22 was ST383 and K45 was ST14, both of which have been designated as high-risk K. pneumoniae clonal complexes and isolated in Egypt before [11,14,35].
Table 2.
Genetic characteristics of CR-hvKP clinical isolates from Assiut, Egypt.
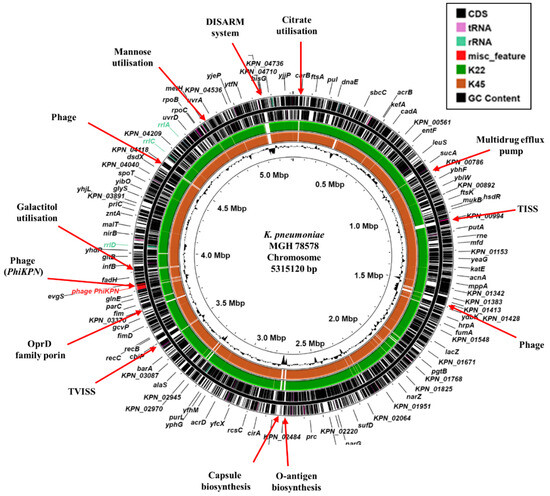
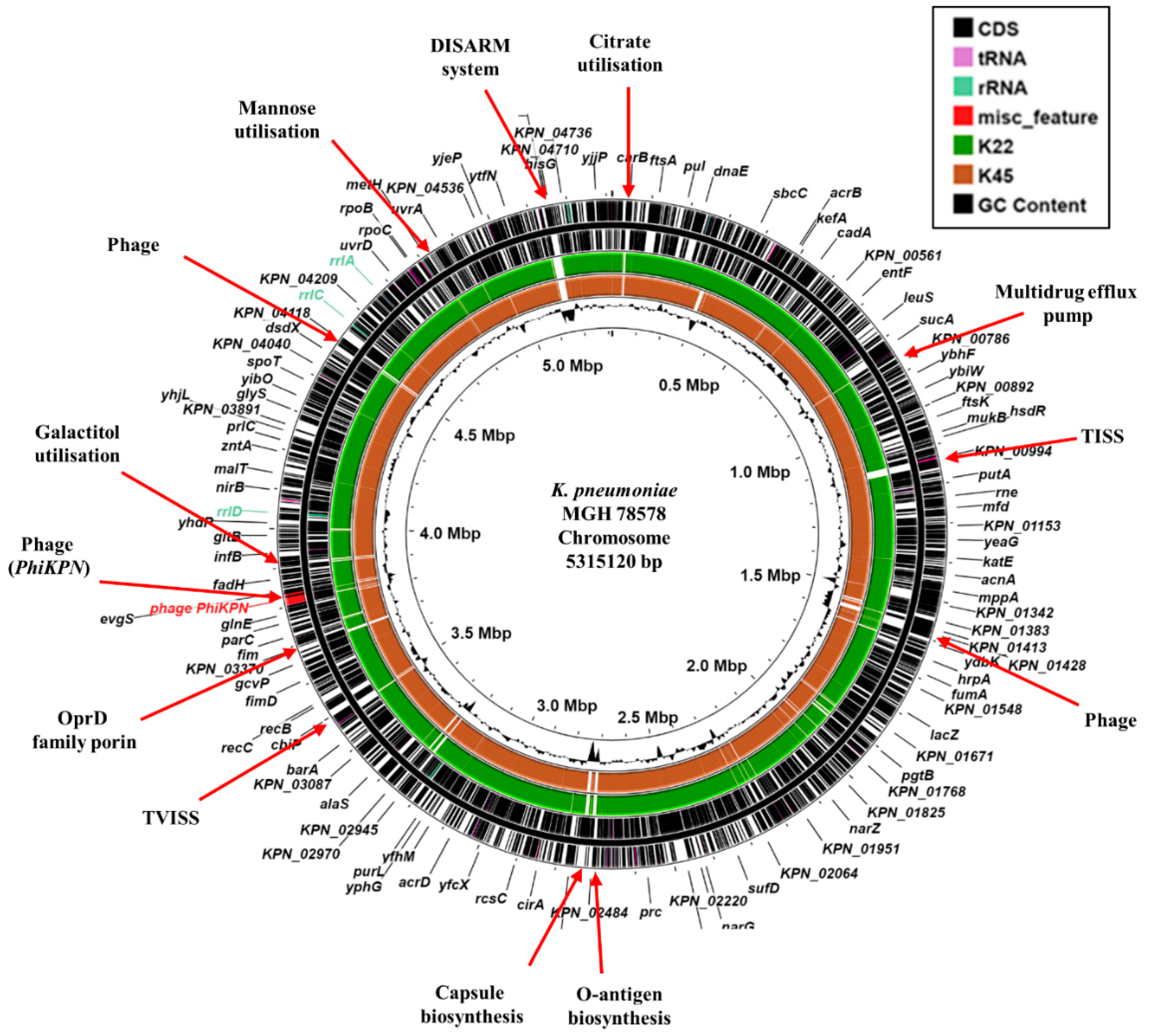
Figure 1.
Comparison of the K. pneumoniae MGH 78578 chromosome with the draft genomes of K. pneumoniae isolates K22 and K45. The figure shows the comparison of the K. pneumoniae MGH 78578 chromosome with the draft genomes of K. pneumoniae K22 and K45, using ProkSee [22]. The two outer rings display the genes/CDS of the K. pneumoniae MGH 78578 chromosome (CP000647.1) with selected features indicated [33]. The green and brown rings depict the BLAST results when the contigs from the draft genome sequences of K22 and K45 are compared with the MGH 78578 chromosome with shaded regions indicating homology. The innermost ring displays the GC content for the K. pneumoniae MGH 78578 chromosome. Abbreviations: TISS, Type I secretion system; TVISS, Type VI secretion system; DISARM, Defense Island System Associated with Restriction-Modification [36].
3.3. Antimicrobial Resistance Genes
Analysis of the antimicrobial resistance genes carried via K22 and K45 agreed with their antimicrobial susceptibility profiles (Table 1), with each strain carrying genes associated with resistance to aminoglycoside, β-lactam, trimethoprim sulfonamide, macrolide, and fosfomycin antibiotics (Table 3). Importantly, both strains carried the carbapenemase gene blaOXA-48, whilst K22 also possessed blaNDM-1, explaining the resistance of both strains to imipenem (Table 1). K22 also possessed the aac(6′)-Ib-cr gene, which affords protection against both aminoglycoside and fluoroquinolone antibiotics, explaining this isolate’s resistance to ciprofloxacin (Table 1). In addition to acquired AMR genes, each strain also carried various point mutations in chromosomal genes such as gyrA, acrR, and parC, which can influence resistance to fluoroquinolones, and alternations in the genes encoding outer membrane porins ompK36 and ompK37, which are associated with carbapenem resistance (Table 3) [26].
Table 3.
Antimicrobial resistance, heavy metal resistance and virulence-associated genes detected in the K22 and K45 CR-hvKP clinical isolates.
3.4. Virulence Determinants
Whole-genome sequence analysis also identified many virulence-associated genes within our isolates (Table 3). These included genes involved in adhesion and biofilm formation, such as those encoding the E. coli common pilus (ecpRABCDE), mannose-sensitive type I fimbriae (fimABCDFGH), and Klebsiella mannose-insensitive type III fimbriae (mrkABCDF, mrkHIJ). Both isolates carried many genes for siderophore production and uptake, which included those for enterobactin production (fepA-entD, entF-fes, fepDGC, entS, fepB, entCEBAH), the aerobactin gene cluster and receptor (iucABCD, iutA), and the salmochelin siderophore esterase and receptor (iroE, iroN). Additionally, K45 also carried the kfuABD iron uptake genes and both strains carried the rmpA and rmpA2 transcription regulators genes, which are commonly found in hvKP strains. It is of note that, in both strains, rmpA2 was a pseudogene, due to a frame shift within the open reading frame.
3.5. Plasmid-Associated Antimicrobial Resistance and Virulence Genes
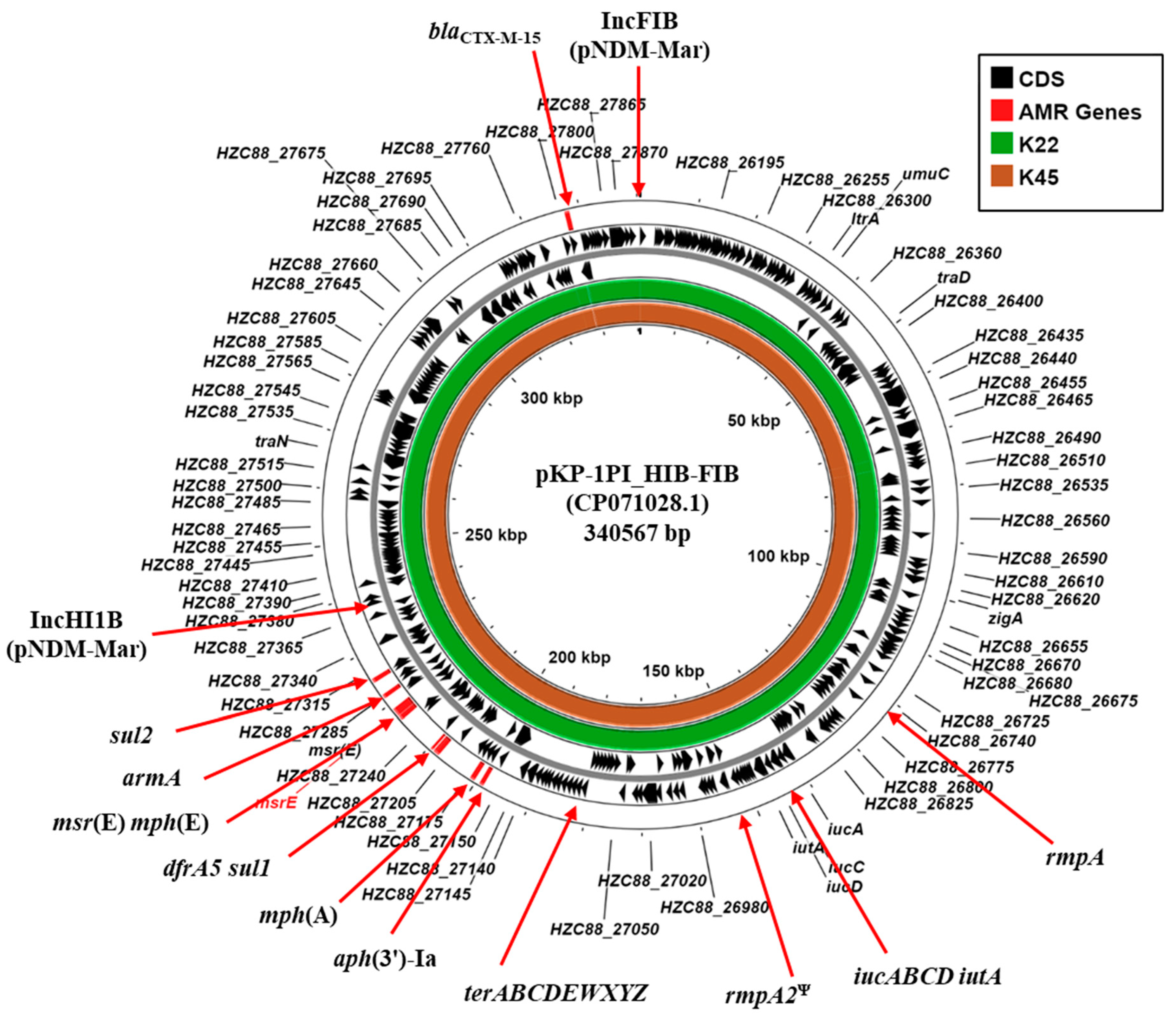
Typical of clinically isolated CR-hvKP strains, K22 and K45, carried multiple plasmid replicons (Table 2). Of particular note is that both K22 and K45 carry identical IncFIB (pNDM-Mar) and IncHI1B (pNDM-Mar) replicons (Supplementary Figure S1). BLASTn analysis of contigs carrying the IncFIB (K22 contig 15: 63,461 bp; K45 contig 25: 62,414 bp) and the IncHI1B replicons (K22 contig 23: 68,431 bp; K25 contig 12: 90,554 bp) indicated that they possess a high degree of similarity to plasmid pKP-1PI_HIB-FIB (CP071028.1) (Figure 2; Supplementary Figure S1). Plasmid pKP-1PI_HIB-FIB (also known as pVIR-147Tu [37]) was isolated in Tuscany, Italy, during an outbreak of an NDM-1-producing K. pneumoniae in 2018. The plasmid was found to be a multi-replicon plasmid formed through the fusion of IncFIB and IncHI1B plasmid backbones that was capable of conjugative transfer. Additionally, pKP-1PI_HIB-FIB carries multiple AMR genes (aph(3′)-Ia, armA, dfrA5, mphE, msrE, mphA, sul1, and sul2,), hypervirulence-associated genes (rmpA, iucABCD-iutA, and a rmpA2 pseudogene), tellurium tolerance genes (terABCDEWXYZ) and various transfer (tra) genes (e.g., traD and traN) (Figure 2; Supplementary Figure S1) [37]. As these genes are similarly organized in the draft genomes of K22 and K45 (Supplementary Figure S1), our results suggest that both strains harbor a similar conjugative IncHI1B/FIB hybrid virulence plasmid, which we term pK22-Vir and pK45-Vir.
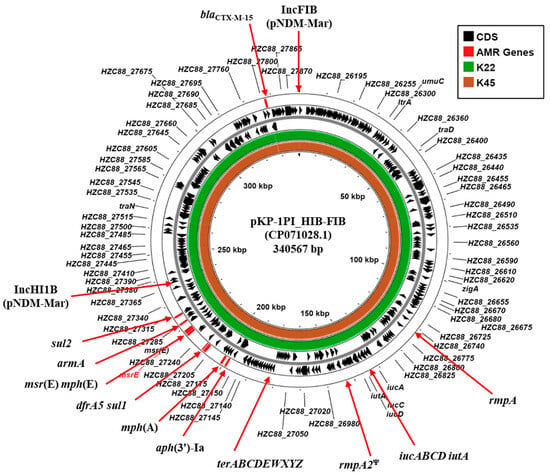
Figure 2.
Comparison of the K. pneumoniae plasmid pKP-1PI_HIB-FIB with the draft genomes of K. pneumoniae isolates K22 and K45. The figure shows the comparison of K. pneumoniae plasmid pKP-1PI_HIB-FIB (also known as pVIR-147Tu [37]) with contigs from the draft genomes of K. pneumoniae K22 and K45, using ProkSee [22]. The genes/CDS of pKP-1PI_HIB-FIB (CP071028.1) is displayed with selected features indicated [37]. The green and brown rings depict the BLAST results when the contigs from the draft genome sequences of K22 and K45 are compared with pKP-1PI_HIB-FIB. Ψ pseudogene.
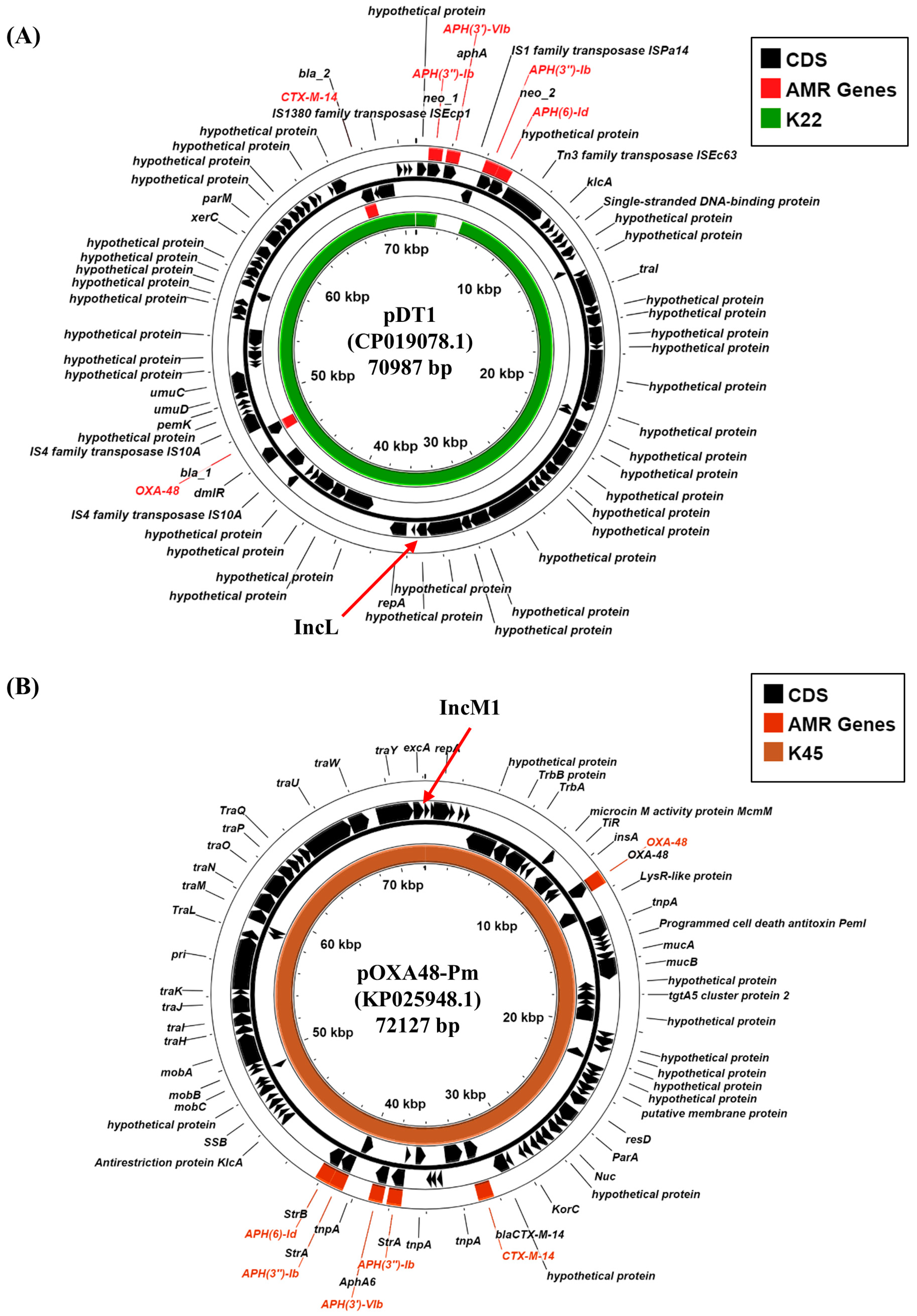
The dissemination of carbapenemase genes in Gram-negative bacteria has been primarily through the transfer of conjugative plasmids. Therefore, we sought to determine which K22 and K45 plasmid replicons harbor these resistance determinants. Our analysis indicates that K. pneumoniae K22 carries an IncL replicon (K22 contig 30: 41,120 bp), whilst K45 a related IncM1 replicon (K45 contig 14: 66,715 bp), both of which have been implicated in the carriage of specific carbapenemase genes (Table 2; Supplementary Figure S2) [38]. Blastn analysis of the K22 IncL-containing contig, which also carries the aph(3″)-Ib and aph(6)-Id AMR genes, indicated that it was identical to plasmid pDT1 (CP019078.1), carried via K. pneumoniae strain DT1, isolated in Hamburg, Germany in 2015 during an ICU outbreak (Supplementary Figure S3A) [39]. Like K22, strain DT1 was sequence type ST383, and pDT1 carried blaOXA-48 and the extended-spectrum beta-lactamase (ESBL) gene, blaCTX-M-14 (Figure 3A). An analysis of the IncM1 replicon from K45, which carries aph(3′)-VIb, aph(3″)-Ib, aph(6)-Id, and blaCTX-M-14b AMR genes, indicated that it was identical to plasmid pOXA48-Pm (KP025948.1), a broad host range plasmid that was isolated from Proteus mirabilis strain Pm-Oxa48 from Gaza in 2012 (Supplementary Figure S3B) [40]. Plasmid pOXA48-Pm also carries multiple AMR genes, including blaOXA-48 and the blaCTX-M-14b ESBL (Figure 3B) [40]. Thus, our analysis suggests that blaOXA-48 carbapenemase genes, carried via K22 and K45, are likely located on IncL and IncM1 plasmids, which we term pK22-OXA48 and pK45-OXA48, respectively (Figure 3). As these plasmids carry multiple tra genes, it is possible that they might be conjugative in nature (Figure 3; Supplementary Figure S2).
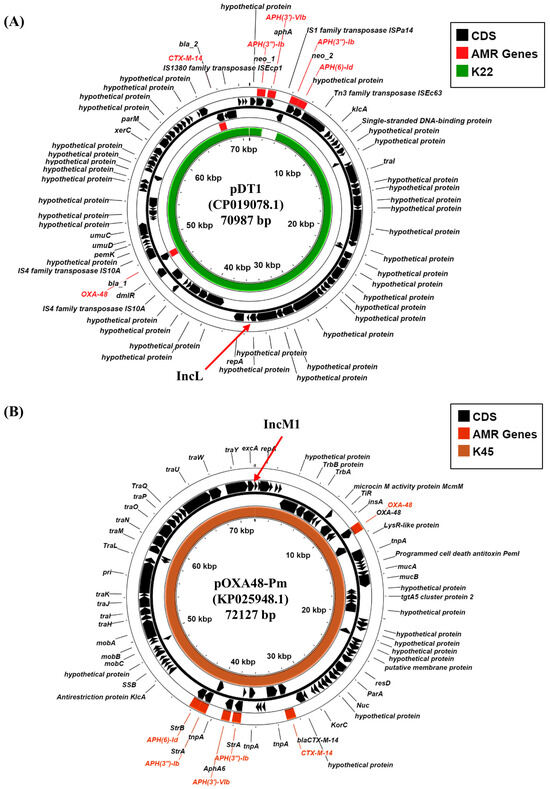
Figure 3.
Analysis of the IncL and IncM plasmid replicons carried via K. pneumoniae isolates K22 and K45. (A) The panel shows a comparison of K. pneumoniae IncL plasmid pDT1 (CP019078.1) [39] with the draft genome of K. pneumoniae K22, using ProkSee [22]. (B) The panel shows a comparison of P. mirabilis IncM1 plasmid pOXA48-Pm (KP025948.1) [40] with the draft genome of K. pneumoniae K45, using ProkSee [22]. The green and brown rings depict the BLAST results when the contigs from the draft genome sequences of K22 and K45, respectively, are compared with each relevant plasmid.
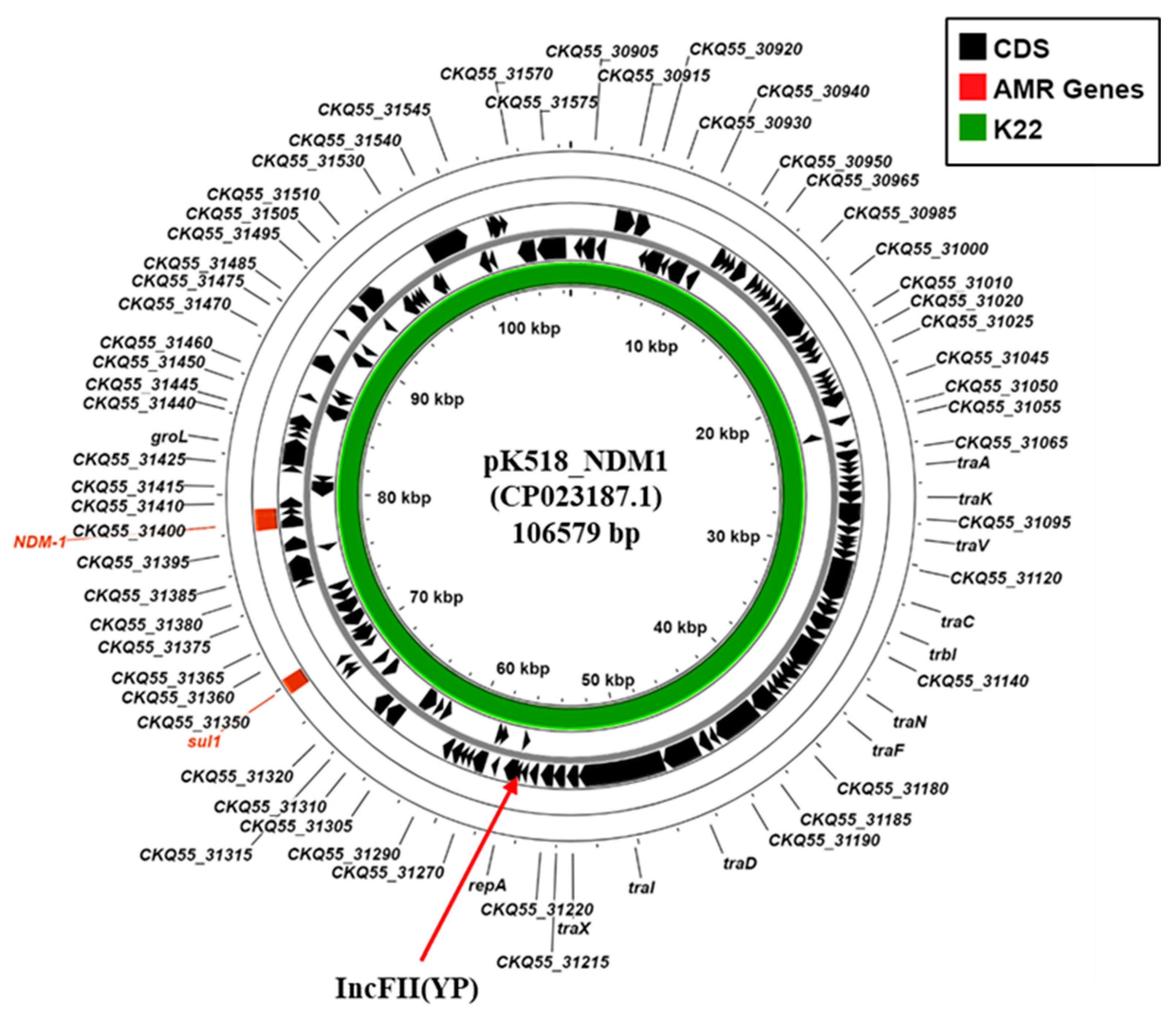
In addition, K. pneumoniae K22 also carries the blaNDM-1 carbapenemase gene. Our analysis indicated that the K22 contigs carrying both blaNDM-1 (K22 contig 34: 30,026 bp) and the IncFII(Yp) replicon (K22 contig 27: 55,018 bp) were very similar to those of the Klebsiella michiganensis plasmid, pK518_NDM1 (CP023187.1) (Figure 4; Supplementary Figure S4), which was isolated from Zhejiang, China, in 2017 [41]. Thus, due to similarities observed between pK518_NDM1, we propose that the blaNDM-1 carbapenemase of K. pneumoniae strain K22 is carried on a similar conjugative IncFII (YP) plasmid, which we term pK22-NDM1.
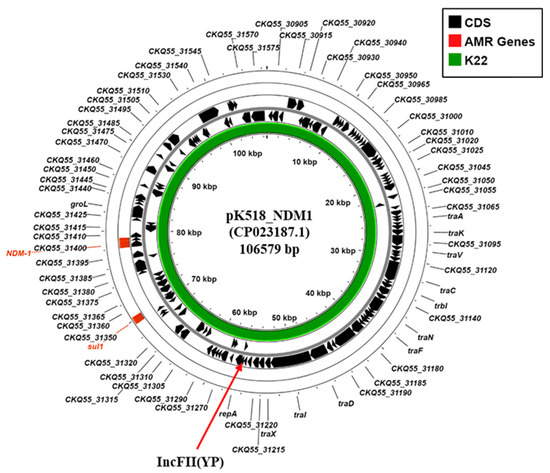
Figure 4.
Analysis of the IncFII(YP) replicon carried via K. pneumoniae isolates K22. The figure shows the comparison of K. michiganensis IncFII(YP) plasmid pK518_NDM1 (CP023187.1) [41] with the draft genome of K. pneumoniae K22, using ProkSee [22]. The green rings depict the BLAST results when the contigs from the draft genome of K22 are compared to pK518_NDM1.
K. pneumoniae strain K22 also carries IncFIB(pQil) and Col440II plasmid replicons, whilst strain K45 carried Col(pHAD28) and Col440I replicons (Table 2; Supplementary Figures S5 and S6). Of note is that the K22 IncFIB(pQil) replicon (K22 contig 38: 18,962 bp) was similar to plasmid pKPN4 (CP000649.1) from the archetypal K. pneumoniae strain MGH 78587 [33], though this was restricted to the surrounding replicon. As none of these replicon contigs carried virulence or AMR genes, their contribution to the virulence and AMR profiles of K22 and K45 is unclear.
4. Discussion
Carbapenem-resistant hypervirulent K. pneumoniae is considered a worldwide threat due to its ability to cause life-threatening infections with limited therapeutic options [4,9]. In the current study, we have investigated the phenotypic and genomic characteristics of two clinical CR-hvKp isolates, K22 and K45, recovered from VAP patients admitted to the pediatric ICU at Assiut University Hospitals in 2015. Both isolates were identified as hypervirulent based on the presence of the virulence genes; iucA, rmpA, and rmpA2 (Table 3) [42]. In addition to specific hypervirulence genes, each strain carried other K. pneumoniae virulence-associated genes, including the mrkABCDF type III fimbriae operon, as well as genes involved in the production and/or uptake of enterobactin and salmochelin siderophores (Table 3). Moreover, strain K45 carried the kfuABD iron scavenging system and was found to be of capsular type K2, which is frequently associated with hypervirulent strains [34,43,44]. Both isolates carried the O1 O-antigen locus suggested to be the most common O-antigen locus detected in hvKp strains [43]. Interestingly, both strains were negative for the string test, most probably due to the presence of a frameshift mutation in rmpA2 (Table 1) [8].
AMR profiling indicated that K22 exhibited an XDR phenotype, whilst K45 was MDR (Table 1). The AMR phenotype was consistent with the presence of many acquired resistance genes and specific chromosomal point mutations detected in their draft genomes (Table 3). Furthermore, both strains carried multiple plasmid replicons, which are localized on contigs containing multiple AMR genes, as well as tra genes (Table 2; Supplementary Figures S1–S3). The detection of numerous resistance genes located on conjugative plasmids suggests that these KP isolates may serve as potential reservoirs for AMR dissemination in the pediatric ICU setting. While no conjugation assays were performed in this study, similar plasmid-mediated transmission has been reported in previous studies [45,46].
Plasmids acquire resistance genes through small mobile elements, such as integrons and transposons, and contribute to resistance expansion through horizontal transmission [47]. In the current study, K22 and K45 were found to carry several resistance/virulence plasmids. Both strains were resistant to the carbapenem imipenem and carried known carbapenemase genes (Table 1 and Table 3). Our analysis indicated that both K22 and K45 carried an IncL/M plasmid (i.e., pK22-OXA48 and pK45-OXA48, respectively) encoding blaOXA-48 carbapenemase, as well as aminoglycosides resistance genes (e.g., aph(3″)-Ib and aph(6)-Id)) (Figure 3; Supplementary Figures S2 and S3). The K22 IncL plasmid pK22-OXA48 was similar to K. pneumoniae pDT1, which was isolated during a German ICU outbreak in 2015 (Figure 3A) [39], whilst the IncM1 plasmid pK45-OXA48, carried by K45, was very similar to P. mirabilis plasmid pOXA48-Pm, isolated in 2012 from Gaza (Figure 3B) [40]. Furthermore, K22 also carries the blaNDM-1 carbapenemase gene on an IncFII(Yp) plasmid (i.e., pK22-NDM1) (Figure 4; Supplementary Figure S4), which is very similar to K. michiganensis plasmid pK518_NDM1 that was isolated in China in 2017 [41]. The presence of similar plasmids among different Gram-negative bacilli could suggest that these plasmids are transmitted by conjugation between different species.
The co-localization of MDR and virulence genes on a single plasmid is a catastrophic incident, likely establishing a speedy evolutionary path for MDR-hvKp from susceptible cKp through a sole conjugation incident [48]. Importantly, both K22 and K45 carried a hybrid IncHI1B/FIB plasmid, which contained both AMR and virulence genes (i.e., rmpA and iucABCD-iutA). As these genes are similarly organized in the draft genomes of K22 and K45 (Supplementary Figure S1), it is suggested that both strains harbor similar IncHI1B/FIB hybrid plasmids (i.e., pK22-Vir and pK45-Vir). Moreover, 39/53 of the hvKp strains isolated later in a second study (2017–2019), harbored both rmpA and iucA genes, suggesting that these 39 hvKp strains might also have carried this hybrid IncHI1B/FIB plasmid [11,12].
Sequence typing indicated that K22 belongs to ST383, whilst K45 was ST14 (Table 1), and both STs are considered as high-risk K. pneumoniae clones. ST14 has been linked to hypervirulence and carbapenemase carriage, being the first K. pneumoniae sequence type in which the blaNDM-1 carbapenemase gene was identified [35,49,50,51,52]. On the other hand, ST383 is an evolving high-risk clonal complex that was first detected in Greek hospitals in 2009 [53]. Since then, it has disseminated throughout the world to many countries, including Egypt where CR-hvKp ST383 isolates have been detected in different cities [11,14,54]. It is noteworthy that a hvKp ST383 K. pneumoniae strain isolated in Alexandria (Egypt) in 2021 and another CR-hvKp ST11 strain isolated in Cairo in 2017, both harbored an IncHI1B/FIB virulence plasmid and an IncL-carrying blaOXA-48 plasmid, similar to those of K22 (Supplementary Figure S7) [14,55].
The IncHI1B/FIB hybrid plasmids found in K22 and K45 are similar to conjugative plasmid pKP-1PI_ HIB-FIB, isolated during an Italian K. pneumoniae outbreak in 2018 (Figure 2; Supplementary Figure S1) [37] and very similar plasmids have been isolated from K. pneumoniae strains in the UK, Poland, Russia, and Egypt (Supplementary Figures S7 and S8) [55,56,57,58,59]. The earliest documented occurrence of this specific type of hybrid IncHI1B/FIB plasmid is pKpvST147L (CM007852.1), which was isolated from a patient in the UK in 2016 (Supplementary Figure S8A) [56]. As this patient’s country of residence was Egypt, this could suggest that their infection with K. pneumoniae occurred whilst the patient was in Egypt. Accordingly, as strain K22 was isolated in 2015, it could possibly be considered to be the “ground zero” strain, being the first isolate to harbor this specific hybrid IncHI1B/FIB AMR/virulence plasmid. Therefore, further large-scale studies are desperately required to reveal the prevalence of such potentially devastating plasmids and to better understand their role in the spread of MDR-hvKp strains in Egypt.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/microorganisms13051058/s1, Figure S1: Analysis of the IncHI1B/FIB hybrid plasmid carried via K. pneumoniae strains K22 and K45; Figure S2: Comparison of the related IncL and IncM1 plasmid replicons carried bviay K. pneumoniae strains K22 and K45; Figure S3: Analysis of the IncL and IncM1 plasmid replicons carried via K. pneumoniae strains K22 and K45; Figure S4: Analysis of the blaNDM-1 carrying plasmid from K. pneumoniae strain K22; Figure S5: Analysis of the IncFIB(pQil) and Col440II plasmid replicons carried via K. pneumoniae isolate K22; Figure S6: Analysis of the Col(pHAD28) and Col440I plasmid replicons carried via K. pneumoniae isolate K45; Figure S7: Analysis of K. pneumoniae strain K22 (EGY22) from Alexandria; Figure S8: Analysis of hybrid K. pneumoniae AMR/virulence plasmids.
Author Contributions
Conceptualization, D.F.B. and S.A.A.; formal analysis, H.A.H., R.A., D.F.B. and S.A.A.; investigation, H.A.H., R.A. and D.F.B.; data curation, D.F.B.; writing—original draft preparation, H.A.H., R.A., D.F.B. and S.A.A.; writing—review and editing, H.A.H., R.A., D.F.B. and S.A.A.; supervision, S.A.A.; project administration, S.A.A.; funding acquisition, D.F.B. and S.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This paper is based upon work supported by the Science, Technology & Innovation Funding Authority (STDF) One Health Grant under grant number 48362 to S.A.A. Additionally, D.F.B. was supported by a Biotechnology and Biological Sciences Research Council (BBSRC) Impact Acceleration Award (IAA) to Aston University and a BBSRC Engineering Biology Mission Award (BB/Y007603/1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All genome sequence data and assemblies have been deposited at NCBI GenBank under BioProject ID PRJNA1079765. The assembled and annotated genomes of Klebsiella pneumoniae strains K22 and K45 have been deposited with the accession numbers JBAMJT000000000 and JBAMJS000000000, respectively.
Acknowledgments
We thank MicrobesNG for genome sequencing and Jane F. Turton for sharing information about the origin of virulence plasmid pKpvST147L. We thank the Children’s Cancer Hospital Egypt (57357) Genomics and Epigenomics Research Program for assisting in genome sequencing of K. pneumoniae.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, X.; Torres, V.V.L.; Liu, H.; Rocker, A.; Zhang, Y.; Wang, J.; Chen, L.; Bi, W.; Lin, J.; et al. An Outbreak of Carbapenem-Resistant and Hypervirulent Klebsiella pneumoniae in an Intensive Care Unit of a Major Teaching Hospital in Wenzhou, China. Front. Public Health 2019, 7, 229. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef]
- Liu, C.; Du, P.; Xiao, N.; Ji, F.; Russo, T.A.; Guo, J. Hypervirulent Klebsiella pneumoniae is emerging as an increasingly prevalent K. pneumoniae pathotype responsible for nosocomial and healthcare-associated infections in Beijing, China. Virulence 2020, 11, 1215–1224. [Google Scholar] [CrossRef]
- Lin, Y.T.; Siu, L.K.; Lin, J.C.; Chen, T.L.; Tseng, C.P.; Yeh, K.M.; Chang, F.Y.; Fung, C.P. Seroepidemiology of Klebsiella pneumoniae colonizing the intestinal tract of healthy Chinese and overseas Chinese adults in Asian countries. BMC Microbiol. 2012, 12, 13. [Google Scholar] [CrossRef]
- Choby, J.E.; Howard-Anderson, J.; Weiss, D.S. Hypervirulent Klebsiella pneumoniae—Clinical and molecular perspectives. J. Intern. Med. 2020, 287, 283–300. [Google Scholar] [CrossRef]
- Shankar, C.; Vasudevan, K.; Jacob, J.J.; Baker, S.; Isaac, B.J.; Neeravi, A.R.; Sethuvel, D.P.M.; George, B.; Veeraraghavan, B. Hybrid Plasmids Encoding Antimicrobial Resistance and Virulence Traits Among Hypervirulent Klebsiella pneumoniae ST2096 in India. Front. Cell Infect. Microbiol. 2022, 12, 875116. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chou, S.H.; Liang, S.W.; Ni, C.E.; Lin, Y.T.; Huang, Y.W.; Yang, T.C. Emergence of an XDR and carbapenemase-producing hypervirulent Klebsiella pneumoniae strain in Taiwan. J. Antimicrob. Chemother. 2018, 73, 2039–2046. [Google Scholar] [CrossRef]
- He, Z.; Xu, W.; Zhao, H.; Li, W.; Dai, Y.; Lu, H.; Zhao, L.; Zhang, C.; Li, Y.; Sun, B. Epidemiological characteristics an outbreak of ST11 multidrug-resistant and hypervirulent Klebsiella pneumoniae in Anhui, China. Front. Microbiol. 2022, 13, 996753. [Google Scholar] [CrossRef]
- Sherif, M.; Palmieri, M.; Mirande, C.; El-Mahallawy, H.; Rashed, H.G.; Abd-El-Reheem, F.; El-Manakhly, A.R.; Abdel-Latif, R.A.R.; Aboulela, A.G.; Saeed, L.Y.; et al. Whole-genome sequencing of Egyptian multidrug-resistant Klebsiella pneumoniae isolates: A multi-center pilot study. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Elzeny, H.; Mohamed, W.; Daef, E.; El-Badawy, O.; Shaaban, L.; Osman, N.S.; Hadiya, S.; Aly, S. Detection of multiple extensively-drug resistant hypervirulent Klebsiella pneumoniae clones from patients with ventilator-associated pneumonia in Egypt. J. Med. Microbiol. 2023, 72, 001701. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Y.; Ahmed, M.; Qin, M.; He, R.; Wu, Y.; Liang, X.; Zhong, L.L.; Chen, P.; Deng, B.; et al. Carriage of distinct bla(KPC-2) and bla(OXA-48) plasmids in a single ST11 hypervirulent Klebsiella pneumoniae isolate in Egypt. BMC Genom. 2022, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Yang, Y.; Yang, Y.; Yan, B.; Chen, G.; Hassan, R.M.; Zhong, L.L.; Chen, Y.; Roberts, A.P.; Wu, Y.; et al. Emergence of Hypervirulent Carbapenem-Resistant Klebsiella pneumoniae Coharboring a bla(NDM-1)-Carrying Virulent Plasmid and a bla(KPC-2)-Carrying Plasmid in an Egyptian Hospital. mSphere 2021, 6, e00088-21. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Document M100, 33rd ed.; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2023. [Google Scholar]
- Yao, H.; Liu, J.; Jiang, X.; Chen, F.; Lu, X.; Zhang, J. Analysis of the Clinical Effect of Combined Drug Susceptibility to Guide Medication for Carbapenem-Resistant Klebsiella pneumoniae Patients Based on the Kirby-Bauer Disk Diffusion Method. Infect. Drug Resist. 2021, 14, 79–87. [Google Scholar] [CrossRef]
- Fang, C.T.; Chuang, Y.P.; Shun, C.T.; Chang, S.C.; Wang, J.T. A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J. Exp. Med. 2004, 199, 697–705. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef] [PubMed]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis Comparison Tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; Edalatmand, A.; Petkau, A.; Syed, S.A.; Tsang, K.K.; et al. CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive Web: User-Friendly Capsule and Lipopolysaccharide Serotype Prediction for Klebsiella Genomes. J. Clin. Microbiol. 2018, 56, e00197-18. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- McClelland, M.; Sanderson, K.E.; Spieth, J.; Clifton, S.W.; Latreille, P.; Courtney, L.; Porwollik, S.; Ali, J.; Dante, M.; Du, F.; et al. Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 2001, 413, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. MMBR 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Ofir, G.; Melamed, S.; Sberro, H.; Mukamel, Z.; Silverman, S.; Yaakov, G.; Doron, S.; Sorek, R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat. Microbiol. 2018, 3, 90–98. [Google Scholar] [CrossRef]
- Di Pilato, V.; Henrici De Angelis, L.; Aiezza, N.; Baccani, I.; Niccolai, C.; Parisio, E.M.; Giordano, C.; Camarlinghi, G.; Barnini, S.; Forni, S.; et al. Resistome and virulome accretion in an NDM-1-producing ST147 sublineage of Klebsiella pneumoniae associated with an outbreak in Tuscany, Italy: A genotypic and phenotypic characterisation. Lancet Microbe 2022, 3, e224–e234. [Google Scholar] [CrossRef]
- Blackwell, G.A.; Doughty, E.L.; Moran, R.A. Evolution and dissemination of L and M plasmid lineages carrying antibiotic resistance genes in diverse Gram-negative bacteria. Plasmid 2021, 113, 102528. [Google Scholar] [CrossRef]
- Both, A.; Büttner, H.; Huang, J.; Perbandt, M.; Belmar Campos, C.; Christner, M.; Maurer, F.P.; Kluge, S.; König, C.; Aepfelbacher, M.; et al. Emergence of ceftazidime/avibactam non-susceptibility in an MDR Klebsiella pneumoniae isolate. J. Antimicrob. Chemother. 2017, 72, 2483–2488. [Google Scholar] [CrossRef]
- Chen, L.; Al Laham, N.; Chavda, K.D.; Mediavilla, J.R.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. First report of an OXA-48-producing multidrug-resistant Proteus mirabilis strain from Gaza, Palestine. Antimicrob. Agents Chemother. 2015, 59, 4305–4307. [Google Scholar] [CrossRef]
- Zheng, B.; Xu, H.; Yu, X.; Lv, T.; Jiang, X.; Cheng, H.; Zhang, J.; Chen, Y.; Huang, C.; Xiao, Y. Identification and genomic characterization of a KPC-2-, NDM-1- and NDM-5-producing Klebsiella michiganensis isolate. J. Antimicrob. Chemother. 2018, 73, 536–538. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of Biomarkers for Differentiation of Hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J. Clin. Microbiol. 2018, 56, e00776-18. [Google Scholar] [CrossRef] [PubMed]
- Follador, R.; Heinz, E.; Wyres, K.L.; Ellington, M.J.; Kowarik, M.; Holt, K.E.; Thomson, N.R. The diversity of Klebsiella pneumoniae surface polysaccharides. Microb. Genom. 2016, 2, e000073. [Google Scholar] [CrossRef]
- Wu, J.W.; Wang, J.T.; Lin, T.L.; Liu, Y.Z.; Wu, L.T.; Pan, Y.J. Identification of three capsule depolymerases in a bacteriophage infecting Klebsiella pneumoniae capsular types K7, K20, and K27 and therapeutic application. J. Biomed. Sci. 2023, 30, 31. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.A.; Mohamed, I.S.; El-Badawy, O.; Zakaria, A.M.; Shabaan, L.; Aly, S.A. pKpQIL-like plasmid contributes to the dissemination of bla(NDM-1) and plasmid mediated quinolone resistance determinants among multi drug resistant Klebsiella pneumoniae in Assiut university hospital, Egypt. Iran. J. Microbiol. 2023, 15, 208–218. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Wang, M.; Li, X.; Liu, Z.; Kuang, D.; Deng, Z.; Ou, H.Y.; Qu, J. Mobilizable plasmids drive the spread of antimicrobial resistance genes and virulence genes in Klebsiella pneumoniae. Genome Med. 2023, 15, 106. [Google Scholar] [CrossRef]
- Abdel-Rahim, M.H.; El-Badawy, O.; Hadiya, S.; Daef, E.A.; Suh, S.J.; Boothe, D.M.; Aly, S.A. Patterns of Fluoroquinolone Resistance in Enterobacteriaceae Isolated from the Assiut University Hospitals, Egypt: A Comparative Study. Microb. Drug Resist. 2019, 25, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef]
- Mei, Y.F.; Liu, P.P.; Wan, L.G.; Liu, Y.; Wang, L.H.; Wei, D.D.; Deng, Q.; Cao, X.W. Virulence and Genomic Feature of a Virulent Klebsiella pneumoniae Sequence Type 14 Strain of Serotype K2 Harboring bla(NDM-5) in China. Front. Microbiol. 2017, 8, 335. [Google Scholar] [CrossRef]
- Al-Zahrani, I.A.; Aljabri, A.; Alhazmi, W.A.; Yasir, M.; Abujamel, T.; Alghamdi, A.K.; Azhar, E.I. Genomic analysis of extensively drug resistant (XDR) Klebsiella pneumoniae high-risk clone ST14 co-harboring bla(NDM) and bla(OXA-48) recovered from Saudi Arabia. J. Infect. Public Health 2024, 17, 669–675. [Google Scholar] [CrossRef]
- Kondo, S.; Phornsiricharoenphant, W.; Na-Rachasima, L.; Phokhaphan, P.; Ruangchai, W.; Palittapongarnpim, P.; Apisarnthanarak, A. Genomic characterization of extended-spectrum β-lactamase-producing Enterobacterales isolated from abdominal surgical patients. Epidemiol. Infect. 2024, 152, e70. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Giakkoupi, P.; Vatopoulos, A.C.; Tryfinopoulou, K.; Miriagou, V.; Tzouvelekis, L.S. Emergence of Klebsiella pneumoniae of a novel sequence type (ST383) producing VIM-4, KPC-2 and CMY-4 β-lactamases. Int. J. Antimicrob. Agents 2010, 36, 573–574. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.K.; Ben Abid, F.; Al Ismail, K.; McElheny, C.L.; Al Maslamani, M.; Omrani, A.S.; Doi, Y. Genomic Epidemiology of Carbapenem-Resistant Klebsiella in Qatar: Emergence and Dissemination of Hypervirulent Klebsiella pneumoniae Sequence Type 383 Strains. Antimicrob. Agents Chemother. 2023, 67, e0003023. [Google Scholar] [CrossRef] [PubMed]
- Edward, E.A.; Mohamed, N.M.; Zakaria, A.S. Whole Genome Characterization of the High-Risk Clone ST383 Klebsiella pneumoniae with a Simultaneous Carriage of bla(CTX-M-14) on IncL/M Plasmid and bla(CTX-M-15) on Convergent IncHI1B/IncFIB Plasmid from Egypt. Microorganisms 2022, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Payne, Z.; Coward, A.; Hopkins, K.L.; Turton, J.A.; Doumith, M.; Woodford, N. Virulence genes in isolates of Klebsiella pneumoniae from the UK during 2016, including among carbapenemase gene-positive hypervirulent K1-ST23 and ‘non-hypervirulent’ types ST147, ST15 and ST383. J. Med. Microbiol. 2018, 67, 118–128. [Google Scholar] [CrossRef]
- Turton, J.; Davies, F.; Turton, J.; Perry, C.; Payne, Z.; Pike, R. Hybrid Resistance and Virulence Plasmids in “High-Risk” Clones of Klebsiella pneumoniae, Including Those Carrying bla(NDM-5). Microorganisms 2019, 7, 326. [Google Scholar] [CrossRef]
- Starkova, P.; Lazareva, I.; Avdeeva, A.; Sulian, O.; Likholetova, D.; Ageevets, V.; Lebedeva, M.; Gostev, V.; Sopova, J.; Sidorenko, S. Emergence of Hybrid Resistance and Virulence Plasmids Harboring New Delhi Metallo-β-Lactamase in Klebsiella pneumoniae in Russia. Antibiotics 2021, 10, 691. [Google Scholar] [CrossRef]
- Falcone, M.; Tiseo, G.; Arcari, G.; Leonildi, A.; Giordano, C.; Tempini, S.; Bibbolino, G.; Mozzo, R.; Barnini, S.; Carattoli, A.; et al. Spread of hypervirulent multidrug-resistant ST147 Klebsiella pneumoniae in patients with severe COVID-19: An observational study from Italy, 2020-21. J. Antimicrob. Chemother. 2022, 77, 1140–1145. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).