
Shift of Microbiota and Modulation of Resistome in the Ceca of Broiler Chicken Fed Berry Pomace Alone or in Combination of a Multienzyme Mixture
 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds Housing, Diets and Experimental Design
2.2. Sample Collections
2.3. Bacterial Enumeration
2.4. DNA Extraction, 16S rRNA Sequencing and Targeting Resistome
2.5. Nuclear Magnetic Resonance (NMR) Analysis
2.6. Statistical Analyses
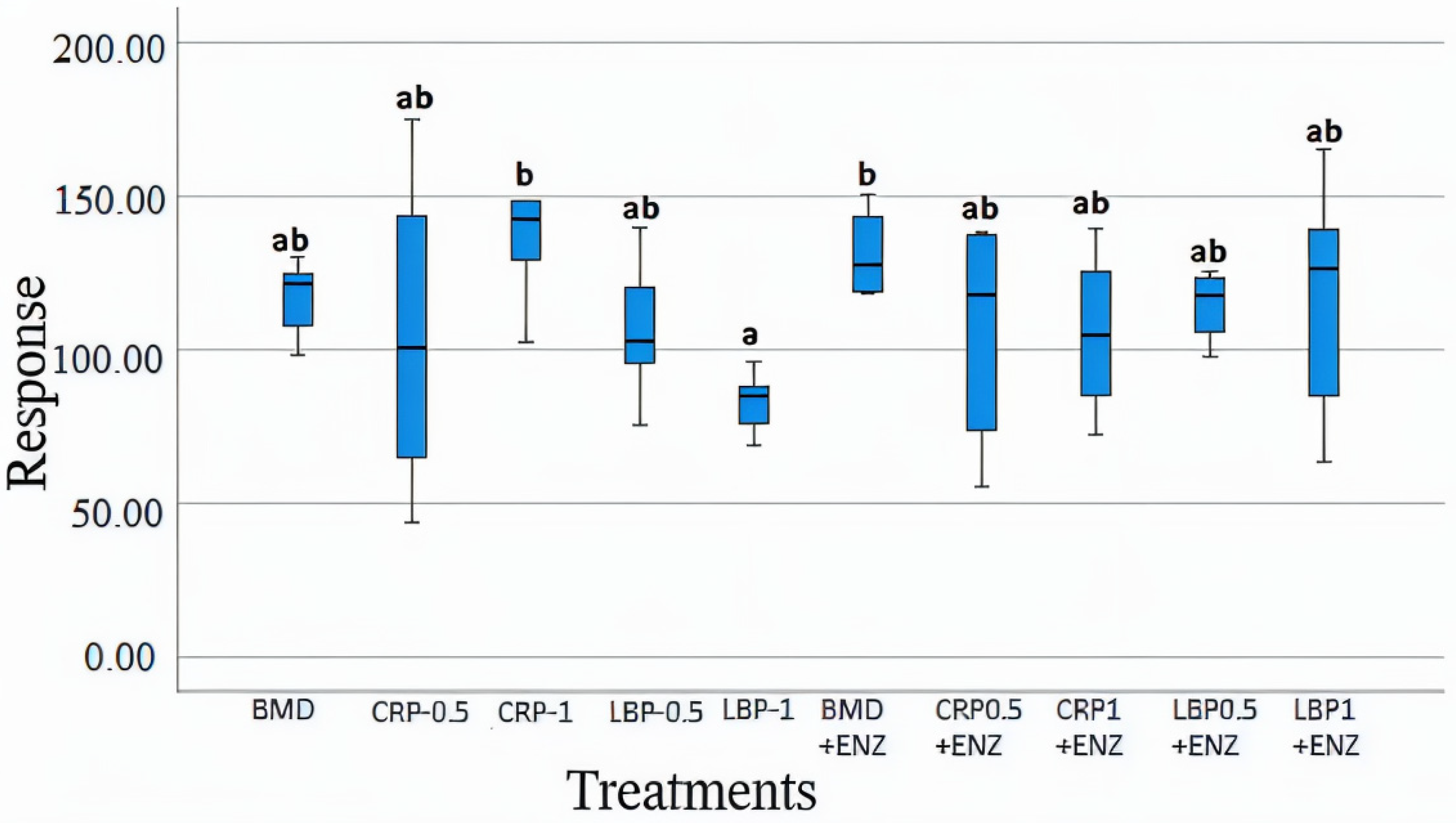
3. Results
3.1. Bacterial Counts
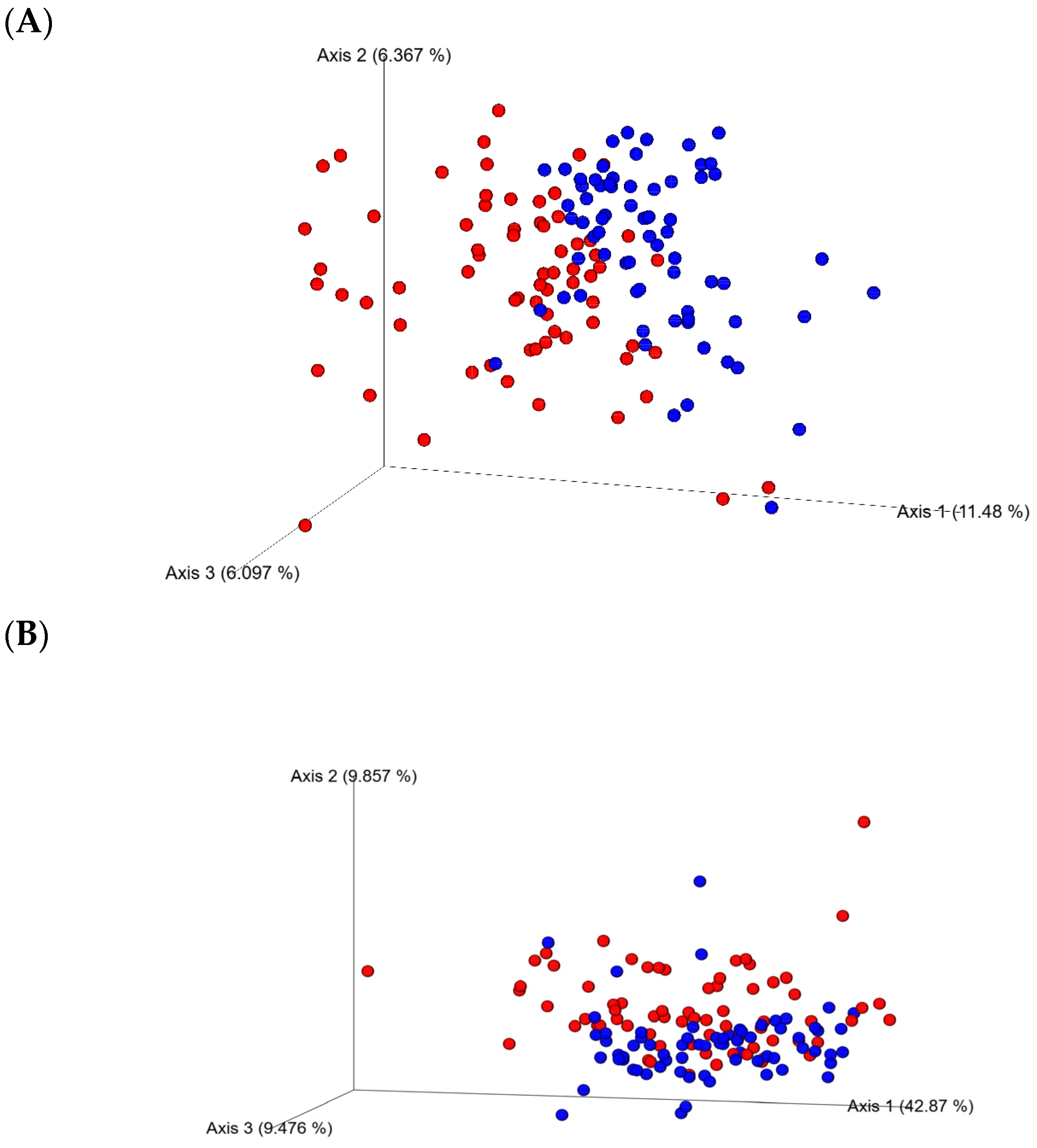
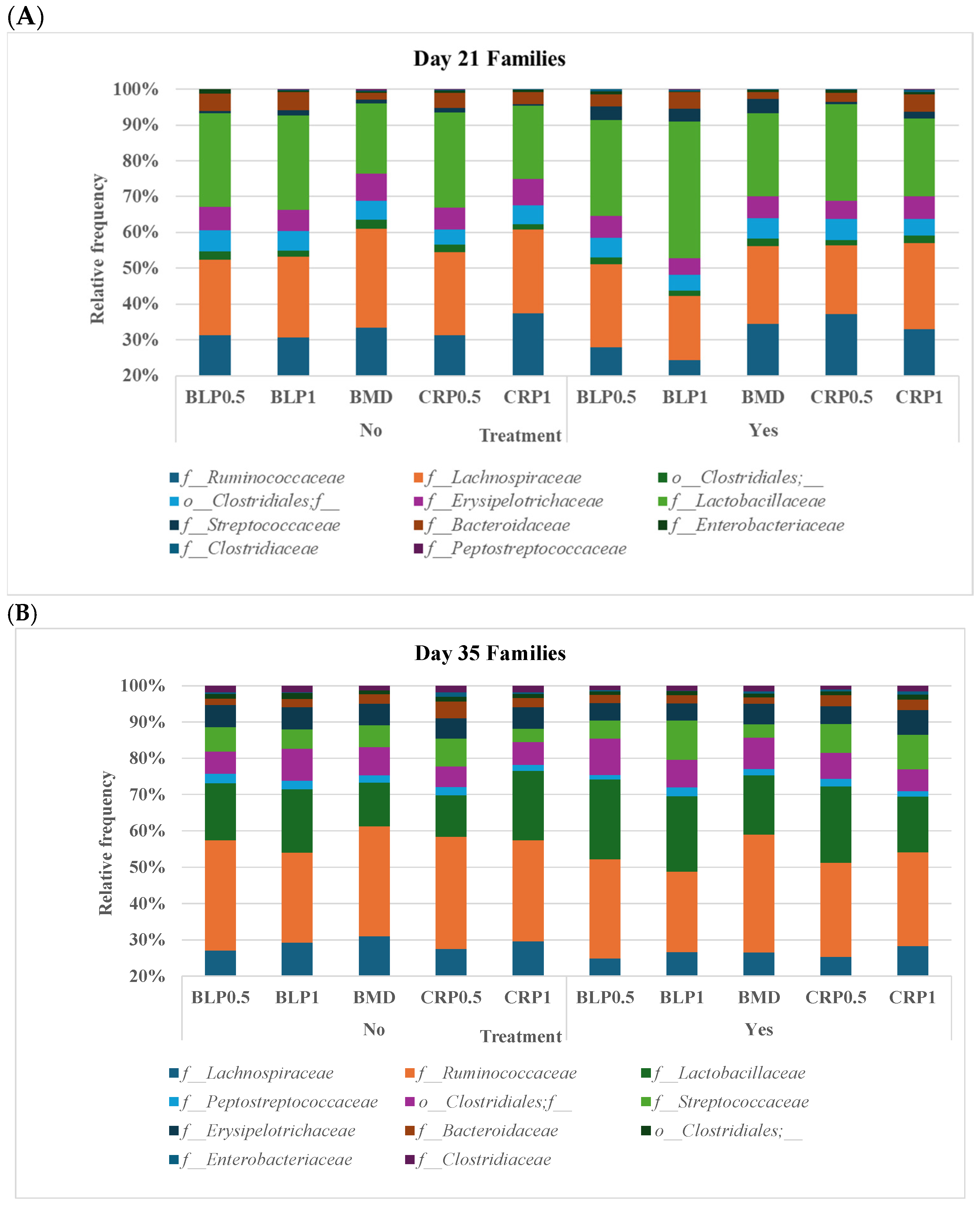
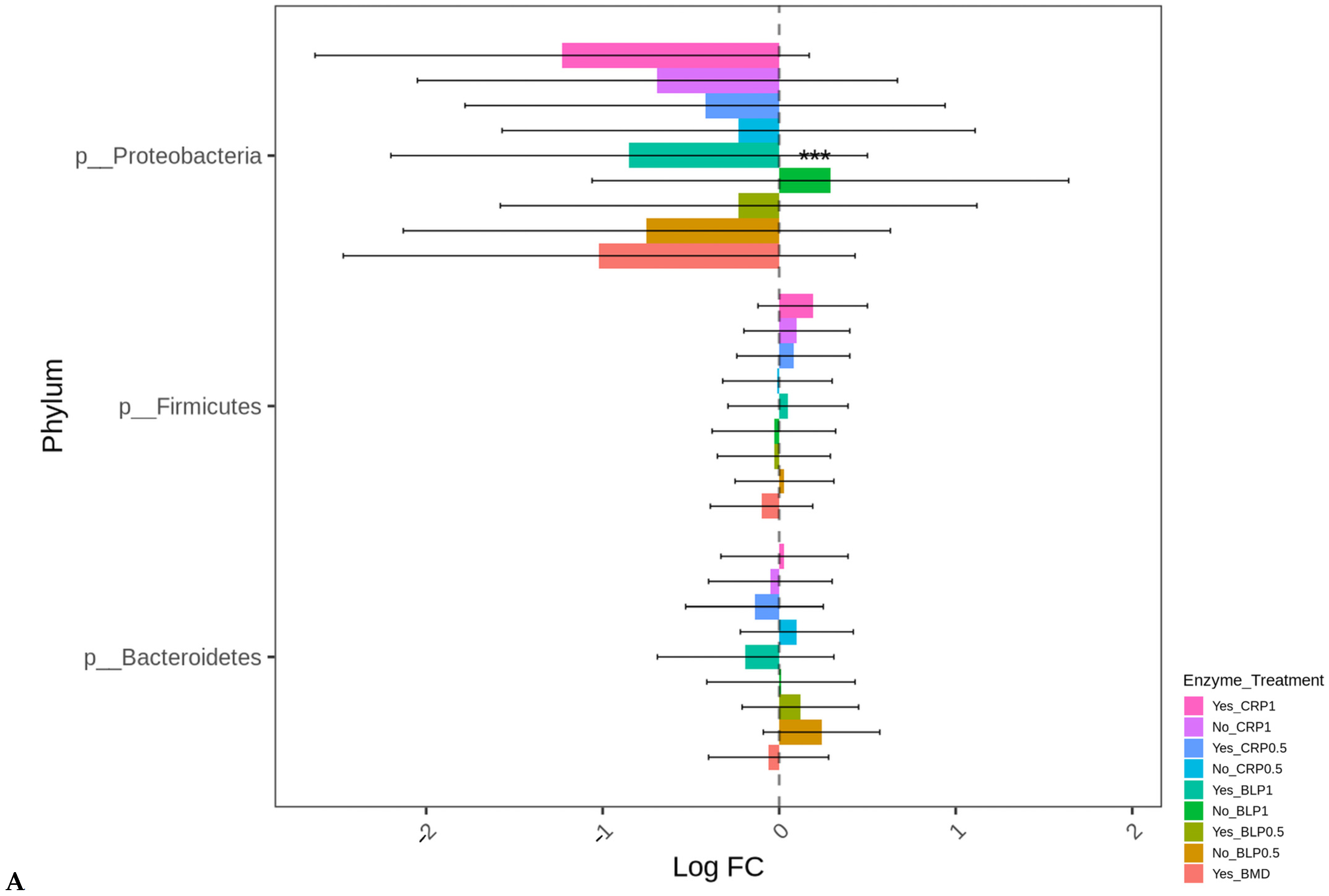
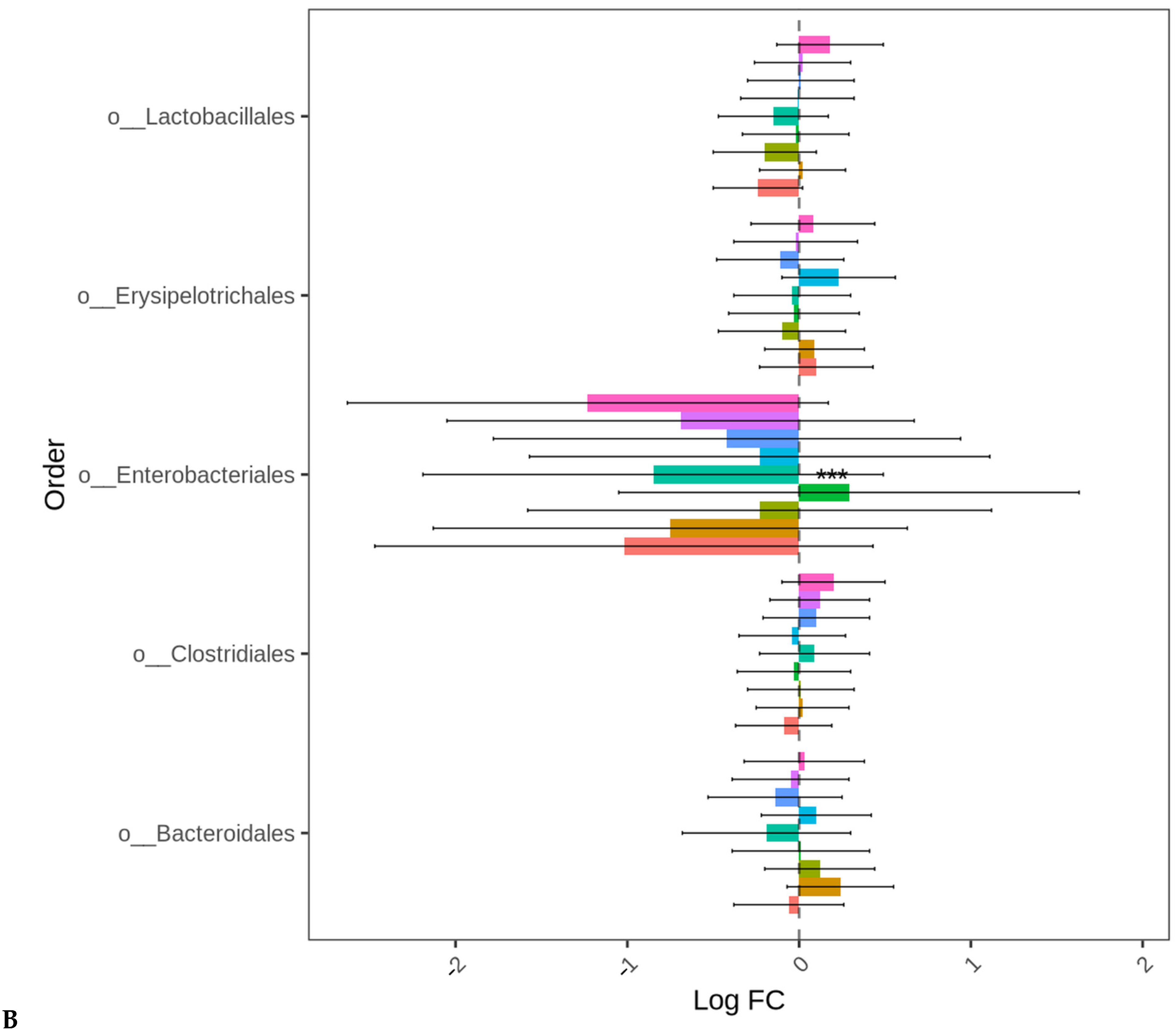
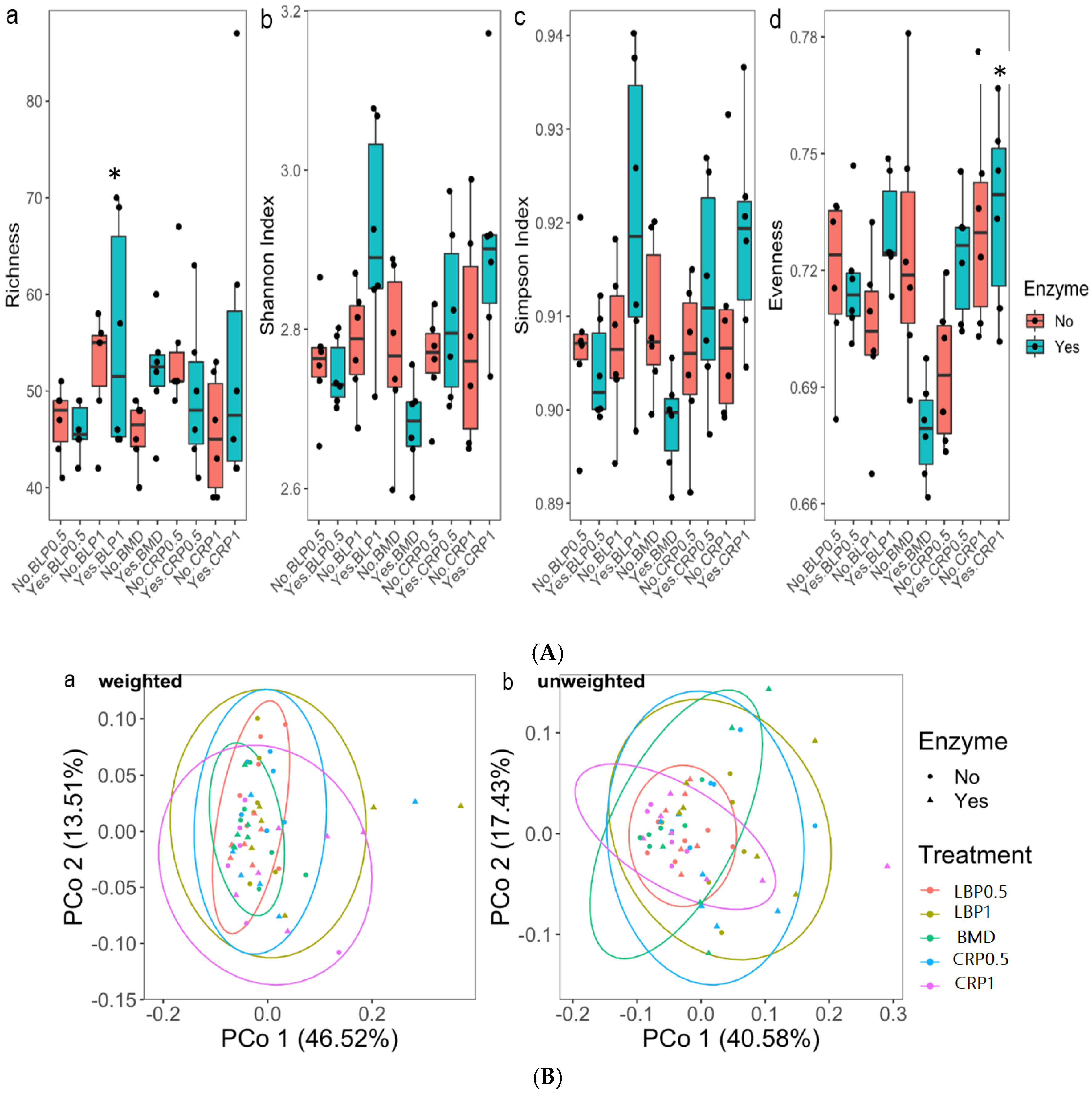
3.2. Ceca Bacterial Diversity
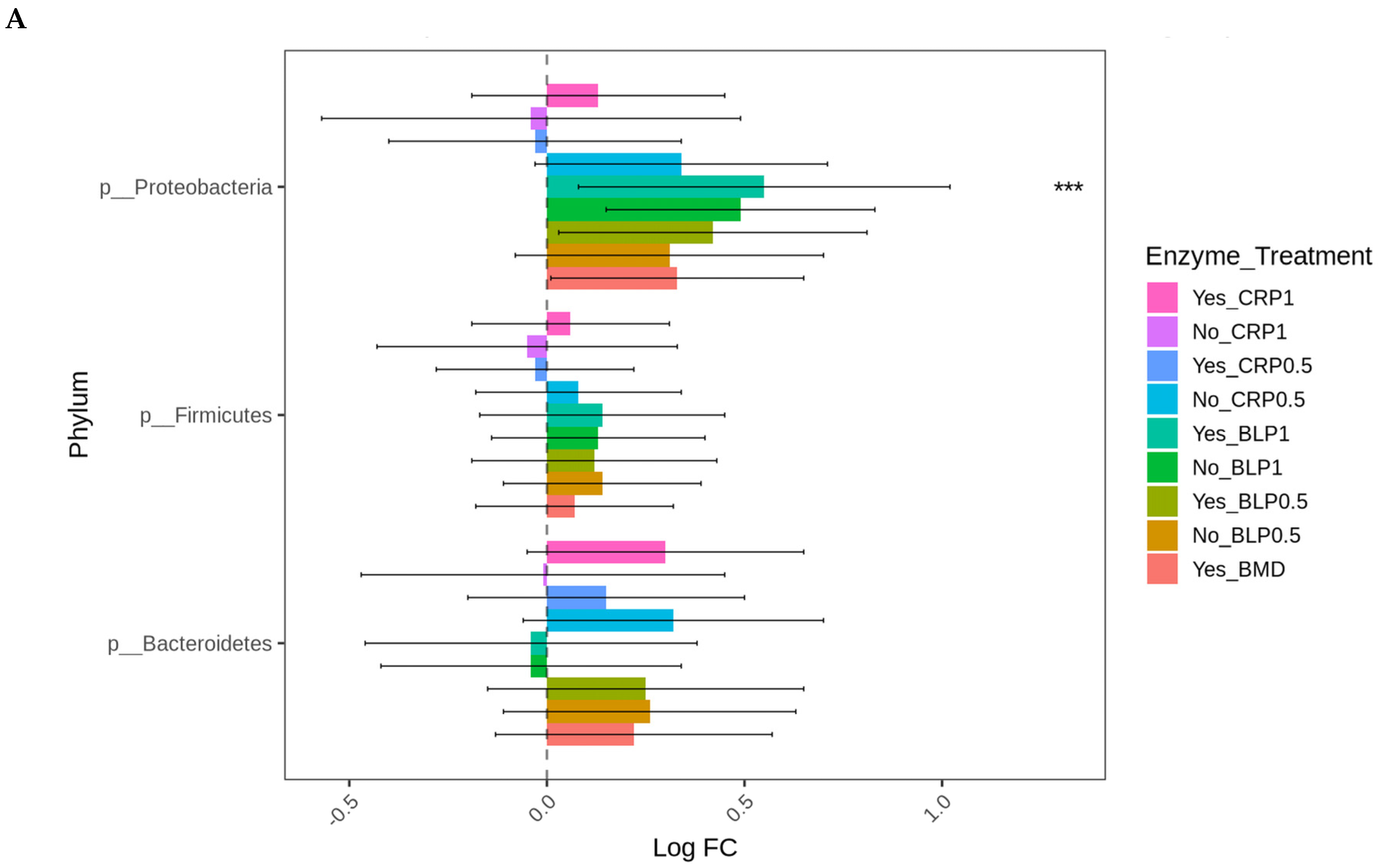
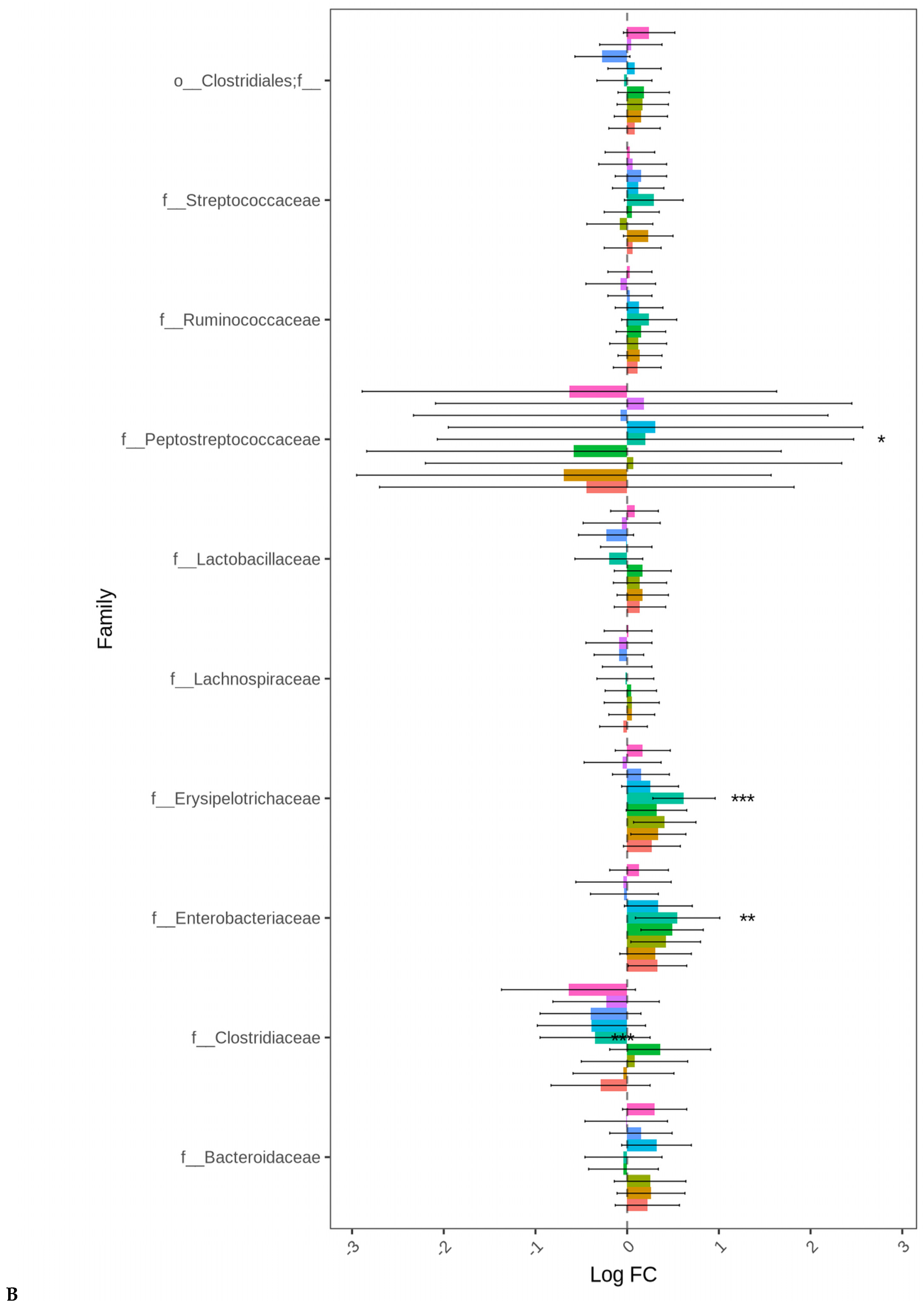
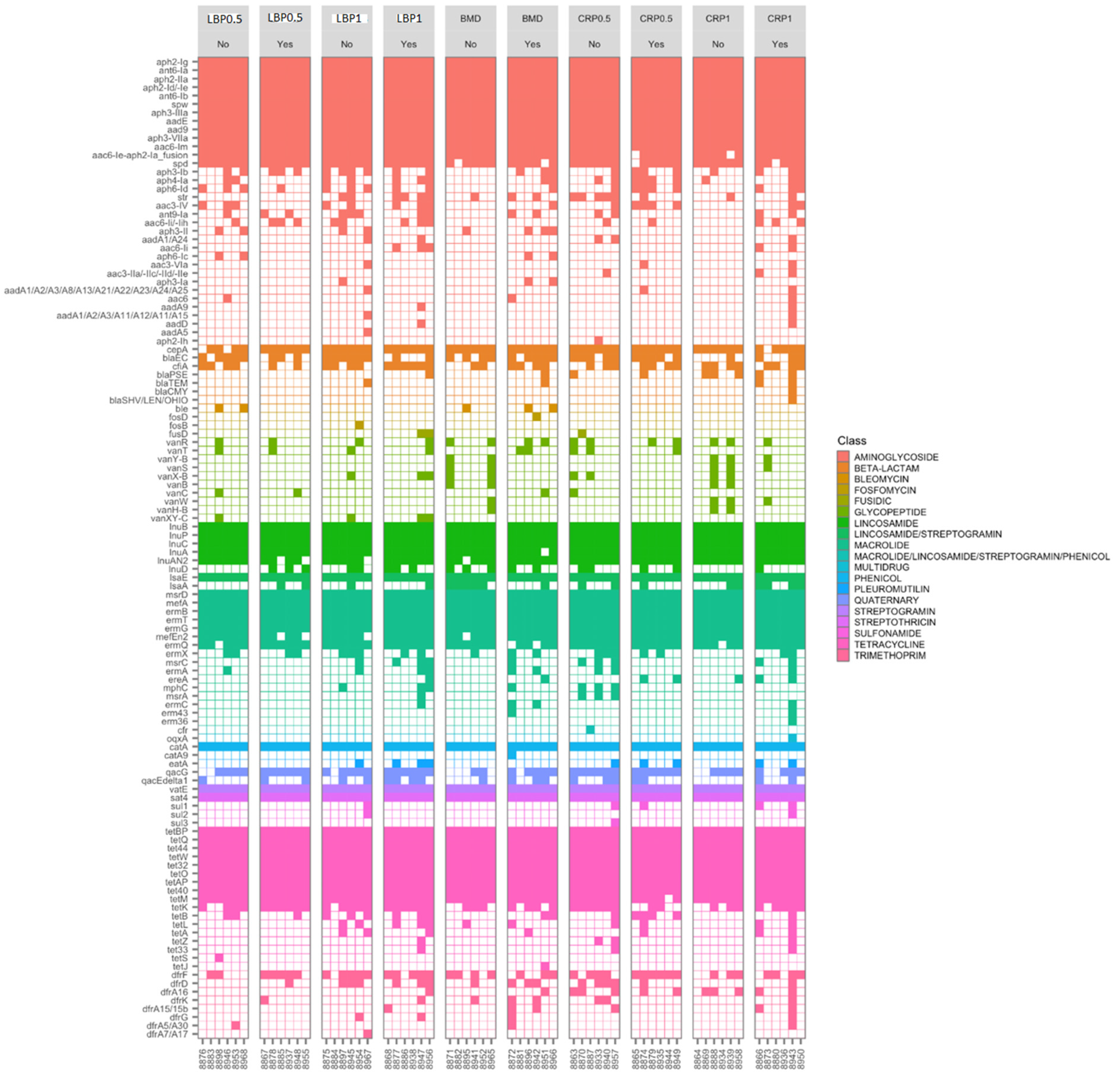
3.3. Targeted Resistome Profile
3.4. Correlations Between Cecal Microbiota and ARGs
3.5. Metabolomics Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D. Diversity and function of the avian gut microbiota. J. Comp. Physiol. B 2012, 182, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Akinyemi, F.T.; Ding, J.; Zhou, H.; Xu, K.; He, C.; Han, C.; Zheng, Y.; Luo, H.; Yang, K.; Gu, C.; et al. Dynamic distribution of gut microbiota during embryonic development in chicken. Poult. Sci. 2020, 99, 5079–5090. [Google Scholar] [CrossRef]
- Van Der Wielen, P.W.J.J.; Keuzenkamp, D.A.; Lipman, L.J.A.; Van Knapen, F.; Biesterveld, S. Spatial and temporal variation of the intestinal bacterial community in commercially raised broiler chickens during growth. Microb. Ecol. 2002, 44, 286–293. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Poole, T.; Sheffield, C. Use and misuse of antimicrobial drugs in poultry and livestock: Mechanisms of antimicrobial resistance. Pak. Vet. J. 2013, 33, 266–271. [Google Scholar]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial Growth Promoters Used in Animal Feed: Effects of Less Well Known Antibiotics on Gram-Positive Bacteria. Clin. Microbiol. Rev. 2003, 16, 175. [Google Scholar] [CrossRef]
- PHAC. Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS); PHAC: Ottawa, ON, Canada, 2016.
- Winokur, P.L.; Vonstein, D.L.; Hoffman, L.J.; Uhlenhopp, E.K.; Doern, G.V. Evidence for transfer of CMY-2 AmpC beta-lactamase plasmids between Escherichia coli and Salmonella isolates from food animals and humans. Antimicrob. Agents Chemother. 2001, 45, 2716–2722. [Google Scholar] [CrossRef]
- CFC. Chicken Farmers of Canada. AMU Strategy. A Prescription for Changel. Available online: https://www.chickenfarmers.ca/antibiotics (accessed on 1 July 2022).
- Chalmers, G.; Cormier, A.C.; Nadeau, M.; Côté, G.; Reid-Smith, R.J.; Boerlin, P. Determinants of virulence and of resistance to ceftiofur, gentamicin, and spectinomycin in clinical Escherichia coli from broiler chickens in Québec, Canada. Vet. Microbiol. 2017, 203, 149–157. [Google Scholar] [CrossRef]
- Boerlin, P.; Reid-Smith, R.J. Antimicrobial resistance: Its emergence and transmission. Anim. Health Res. Rev. 2008, 9, 115–126. [Google Scholar] [CrossRef]
- Guo, R.; Chang, Y.; Wang, D.; Sun, H.; Zhao, A.; Gu, T.; Zong, Y.; Zhou, S.; Huang, Z.; Chen, L. Multiple Omics Find New Cecal Microbial Features Associated with Feed Efficiency in Ducks. bioRxiv 2024. [Google Scholar] [CrossRef]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.-M.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur Resistance in Salmonella enterica Serovar Heidelberg from Chicken Meat and Humans, Canada. Emerg. Infect. Dis. J. 2010, 16, 48. [Google Scholar] [CrossRef]
- Poppe, C.; Martin, L.C.; Gyles, C.L.; Reid-Smith, R.; Boerlin, P.; McEwen, S.A.; Prescott, J.F.; Forward, K.R. Acquisition of Resistance to Extended-Spectrum Cephalosporins by Salmonella enterica subsp. enterica Serovar Newport and Escherichia coli in the Turkey Poult Intestinal Tract. Appl. Environ. Microbiol. 2005, 71, 1184. [Google Scholar] [CrossRef]
- Taggar, G.; Rehman, M.A.; Yin, X.; Lepp, D.; Ziebell, K.; Handyside, P.; Boerlin, P.; Diarra, M.S. Antimicrobial-Resistant E. coli from Surface Waters in Southwest Ontario Dairy Farms. J. Environ. Qual. 2018, 47, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Yin, R.; Kuo, H.-C.; Hudlikar, R.; Sargsyan, D.; Li, S.; Wang, L.; Wu, R.; Kong, A.-N. Gut microbiota, dietary phytochemicals, and benefits to human health. Curr. Pharmacol. Rep. 2019, 5, 332–344. [Google Scholar]
- Das, Q.; Islam, M.R.; Lepp, D.; Tang, J.; Yin, X.; Mats, L.; Liu, H.; Ross, K.; Kennes, Y.M.; Yacini, H. Gut Microbiota, Blood Metabolites, and Spleen Immunity in Broiler Chickens Fed Berry Pomaces and Phenolic-Enriched Extractives. Front. Vet. Sci. 2020, 7, 150. [Google Scholar] [CrossRef]
- Islam, M.R.; Hassan, Y.I.; Das, Q.; Lepp, D.; Hernandez, M.; Godfrey, D.V.; Orban, S.; Ross, K.; Delaquis, P.; Diarra, M.S. Dietary organic cranberry pomace influences multiple blood biochemical parameters and cecal microbiota in pasture-raised broiler chickens. J. Funct. Foods 2020, 72, 104053. [Google Scholar] [CrossRef]
- Islam, M.R.; Oomah, D.B.; Diarra, M.S. Potential immunomodulatory effects of non-dialyzable materials of cranberry extract in poultry production. Poult. Sci. 2017, 96, 341–350. [Google Scholar] [CrossRef]
- Juskiewicz, J.; Jankowski, J.; Kosmala, M.; Zdunczyk, Z.; Slominski, B.A.; Zdunczyk, P. The effects of dietary dried fruit pomaces on growth performance and gastrointestinal biochemistry of turkey poults. J. Anim. Physiol. Anim. Nutr. 2016, 100, 967–976. [Google Scholar] [CrossRef]
- Meng, X.; Slominski, B.; Nyachoti, C.; Campbell, L.; Guenter, W. Degradation of cell wall polysaccharides by combinations of carbohydrase enzymes and their effect on nutrient utilization and broiler chicken performance. Poult. Sci. 2005, 84, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Slominski, B.A.; Meng, X.; Campbell, L.D.; Guenter, W.; Jones, O. The Use of Enzyme Technology for Improved Energy Utilization from Full-Fat Oilseeds. Part II: Flaxseed. Poult. Sci. 2006, 85, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Habeanu, M.; Gras, M.; Pistol, G.; Lefter, N.; Palade, M.; Ropota, M.; Sanda Chedea, V.; Marin, D.E. Assessment of the effect of grape seed cake inclusion in the diet of healthy fattening-finishing pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e30–e42. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shafi, M.E.; Qattan, S.Y.; Batiha, G.E.; Khafaga, A.F.; Abdel-Moneim, A.M.E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1835–1850. [Google Scholar] [CrossRef]
- Koorakula, R.; Schiavinato, M.; Ghanbari, M.; Wegl, G.; Grabner, N.; Koestelbauer, A.; Klose, V.; Dohm, J.C.; Domig, K.J. Metatranscriptomic analysis of the chicken gut resistome response to in-feed antibiotics and natural feed additives. Front. Microbiol. 2022, 13, 833790. [Google Scholar] [CrossRef]
- Kithama, M.; Ross, K.; Diarra, M.S.; Kiarie, E.G. Utilization of grape (Vitis vinifera), cranberry (Vaccinium macrocarpon), wild blueberry (Vaccinium angustifolium) and apple (Malus pumila/domestica) pomaces in broiler chickens when fed without or with multi-enzyme supplement. Can. J. Anim. Sci. 2022, 103, 15–25. [Google Scholar] [CrossRef]
- Kithama, M.; Hassan, Y.I.; Yin, X.; Ross, K.; Julien, C.; Kennes, Y.-M.; Kiarie, E.G.; Diarra, M.S. Growth performance, organ weight, and plasma metabolites in broiler chickens fed corn-soybean meal diet containing berry pomaces and fed without or with multienzymes supplement. Poult. Sci. 2023, 102, 102544. [Google Scholar] [CrossRef]
- CCAC. Guidelines on the Care and Use of Farm Animals in Research, Teaching and Testing; CCAC: Ottawa, ON, Canada, 2009. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Prodan, A.; Tremaroli, V.; Brolin, H.; Zwinderman, A.H.; Nieuwdorp, M.; Levin, E. Comparing bioinformatic pipelines for microbial 16S rRNA amplicon sequencing. PLoS ONE 2020, 15, e0227434. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Asimenos, G.; Toh, H. Multiple alignment of DNA sequences with MAFFT. Methods Mol. Biol. 2009, 537, 39–64. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Dabdoub, S.M.; Fellows, M.L.; Paropkari, A.D.; Mason, M.R.; Huja, S.S.; Tsigarida, A.A.; Kumar, P.S. PhyloToAST: Bioinformatics tools for species-level analysis and visualization of complex microbial datasets. Sci. Rep. 2016, 6, 29123. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbo, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- Morton, J.T.; Marotz, C.; Washburne, A.; Silverman, J.; Zaramela, L.S.; Edlund, A.; Zengler, K.; Knight, R. Establishing microbial composition measurement standards with reference frames. Nat. Commun. 2019, 10, 2719. [Google Scholar] [CrossRef]
- Fedarko, M.W.; Martino, C.; Morton, J.T.; Gonzalez, A.; Rahman, G.; Marotz, C.A.; Minich, J.J.; Allen, E.E.; Knight, R. Visualizing ‘omic feature rankings and log-ratios using Qurro. NAR Genom. Bioinform. 2020, 2, lqaa023. [Google Scholar] [CrossRef]
- Shay, J.A.; Haniford, L.S.; Cooper, A.; Carrillo, C.D.; Blais, B.W.; Lau, C.H.F. Exploiting a targeted resistome sequencing approach in assessing antimicrobial resistance in retail foods. Environ. Microbiome 2022, 18, 25. [Google Scholar] [CrossRef]
- Kim, H.K.; Kostidis, S.; Choi, Y.H. NMR Analysis of Fecal Samples. In Clinical Metabolomics: Methods and Protocols; Giera, M., Ed.; Springer: New York, NY, USA, 2018; pp. 317–328. [Google Scholar]
- Saladrigas-García, M.; Solà-Oriol, D.; López-Vergé, S.; D’Angelo, M.; Collado, M.C.; Nielsen, B.; Faldyna, M.; Pérez, J.F.; Martín-Orúe, S.M. Potential effect of two Bacillus probiotic strains on performance and fecal microbiota of breeding sows and their piglets. J. Anim. Sci. 2022, 100, skac163. [Google Scholar] [CrossRef]
- SAS Institute. Base SAS 9.4 Procedures Guide: Statistical Procedures; SAS Institute Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Lau, C.H.-F.; Capitani, S.; Tien, Y.-C.; Verellen, L.A.; Kithama, M.; Kang, H.; Kiarie, E.G.; Topp, E.; Diarra, M.S.; Fruci, M. Dynamic effects of black soldier fly larvae meal on the cecal bacterial microbiota and prevalence of selected antimicrobial resistant determinants in broiler chickens. Anim. Microbiome 2024, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Alireza, S.; Marco, R.; Vito, L.; Vincenzo, T. Practical applications of agricultural wastes in poultry feeding in Mediterranean and Middle East regions. Part 1: Citrus, grape, pomegranate and apple wastes. World’s Poult. Sci. J. 2018, 74, 489–498. [Google Scholar] [CrossRef]
- Heidarisafar, Z.; Sadeghi, G.; Karimi, A.; Azizi, O. Apple peel waste as a natural antioxidant for heat-stressed broiler chickens. Trop. Anim. Health Prod. 2016, 48, 831–835. [Google Scholar] [CrossRef]
- Islam, M.R.; Lepp, D.; Godfrey, D.V.; Orban, S.; Ross, K.; Delaquis, P.; Diarra, M.S. Effects of wild blueberry (Vaccinium angustifolium) pomace feeding on gut microbiota and blood metabolites in free-range pastured broiler chickens. Poult. Sci. 2019, 98, 3739–3755. [Google Scholar] [CrossRef]
- Tejeda, O.J.; Kim, W.K. Role of Dietary Fiber in Poultry Nutrition. Animals 2021, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.A.; Ehret, D.; Godfrey, D.; Fukumoto, L.; Diarra, M. Characterization of Pilot Scale Processed Canadian Organic Cranberry (Vaccinium macrocarpon) and Blueberry (Vaccinium angustifolium) Juice Pressing Residues and Phenolic-Enriched Extractives. Int. J. Fruit Sci. 2017, 17, 202–232. [Google Scholar] [CrossRef]
- Pajarillo, E.A.B.; Lee, E.; Kang, D.-K. Trace metals and animal health: Interplay of the gut microbiota with iron, manganese, zinc, and copper. Anim. Nutr. 2021, 7, 750–761. [Google Scholar] [CrossRef]
- Maeusli, M.; Skandalis, N.; Lee, B.; Lu, P.; Miller, S.; Yan, J.; Talyansky, Y.; Li, R.; Reyna, Z.; Guerrero, N.; et al. Acacia Fiber Protects the Gut from Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Colonization Enabled by Antibiotics. Msphere 2022, 7, e00071-22. [Google Scholar] [CrossRef] [PubMed]
- Revolledo, L.; Ferreira, A.J.P.; Mead, G.C. Prospects in Salmonella Control: Competitive Exclusion, Probiotics, and Enhancement of Avian Intestinal Immunity. J. Appl. Poult. Res. 2006, 15, 341–351. [Google Scholar] [CrossRef]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Józefiak, D.; Rutkowski, A.; Martin, S.A. Carbohydrate fermentation in the avian ceca: A review. Anim. Feed Sci. Technol. 2004, 113, 1–15. [Google Scholar] [CrossRef]
- Lister, S.A.; Barrow, P. Chapter 8—Enterobacteriaceae. In Poultry Diseases, 6th ed.; Pattison, M., McMullin, P.F., Bradbury, J.M., Alexander, D.J., Eds.; W.B. Saunders: Edinburgh, UK, 2008; pp. 110–145. [Google Scholar]
- Cho, S.; Kumar, S.S.; Ramirez, S.; Valientes, R.; Kim, I.H. Dietary eubiotics of microbial muramidase and glycan improve intestinal villi, ileum microbiota composition and production trait of broiler. J. Anim. Sci. Biotechnol. 2024, 15, 59. [Google Scholar] [CrossRef]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, V. Feed enzymes: The science, practice, and metabolic realities. J. Appl. Poult. Res. 2013, 22, 628–636. [Google Scholar] [CrossRef]
- Slominski, B.A. Recent advances in research on enzymes for poultry diets. Poult. Sci. 2011, 90, 2013–2023. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Das, Q.; Rehman, M.A.; Yin, X.; Shay, J.; Gauthier, M.; Lau, C.H.-F.; Ross, K.; Diarra, M.S. Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces. Microorganisms 2023, 11, 1184. [Google Scholar] [CrossRef]
- Sydenham, T.V.; Sóki, J.; Hasman, H.; Wang, M.; Justesen, U.S. Identification of antimicrobial resistance genes in multidrug-resistant clinical Bacteroides fragilis isolates by whole genome shotgun sequencing. Anaerobe 2015, 31, 59–64. [Google Scholar] [CrossRef]
- Gutacker, M.; Valsangiacomo, C.; Piffaretti, J.-C. Identification of two genetic groups in Bacteroides fragilis by multilocus enzyme electrophoresis: Distribution of antibiotic resistance (cfiA, cepA) and enterotoxin (bft) encoding genes. Microbiology 2000, 146, 1241–1254. [Google Scholar] [CrossRef]
- Abad, C.L.; Safdar, N. The Role of Lactobacillus Probiotics in the Treatment or Prevention of Urogenital Infections—A Systematic Review. J. Chemother. 2009, 21, 243–252. [Google Scholar] [CrossRef]
- Oliver, A.; Xue, Z.; Villanueva, Y.T.; Durbin-Johnson, B.; Alkan, Z.; Taft, D.H.; Liu, J.; Korf, I.; Laugero, K.D.; Stephensen, C.B.; et al. Association of Diet and Antimicrobial Resistance in Healthy U.S. Adults. mBio 2022, 13, e0010122. [Google Scholar] [CrossRef]
- Bilal, M.; Achard, C.; Barbe, F.; Chevaux, E.; Ronholm, J.; Zhao, X. Bacillus pumilus and Bacillus subtilis Promote Early Maturation of Cecal Microbiota in Broiler Chickens. Microorganisms 2021, 9, 1899. [Google Scholar] [CrossRef] [PubMed]
- Colombino, E.; Ferrocino, I.; Biasato, I.; Cocolin, L.S.; Prieto-Botella, D.; Zduńczyk, Z.; Jankowski, J.; Milala, J.; Kosmala, M.; Fotschki, B.; et al. Dried fruit pomace inclusion in poultry diet: Growth performance, intestinal morphology and physiology. J. Anim. Sci. Biotechnol. 2020, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Shao, Y.; Sun, G.; Yang, Y.; Zhang, L.; Guo, Y.; Luo, X.; Lu, L. The relationship among gut microbiota, short-chain fatty acids, and intestinal morphology of growing and healthy broilers. Poult. Sci. 2020, 99, 5883–5895. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No Enzyme | Plus Enzyme | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day | * Treatment | Treatment | p-Values | |||||||||||
| BMD | LBP0.5 | LBP1 | CRP0.5 | CRP1 | BMD | LBP0.5 | LBP1 | CRP0.5 | CRP1 | SEM | ENZ | Treat * | ENZ * Treat | |
| 21 | 1.9 a | 0.6 c | 0.56 c | 1.61 ab | 1.38 b | 1.52 b | 1.67 ab | 1.81 a | 1.44 b | 1.57 b | 0.336 | 0.067 | 0.378 | 0.054 |
| 35 | 2.38 | 2.71 | 2.62 | 1.86 | 2.38 | 2.24 | 1.76 | 2.22 | 1.27 | 2.33 | 0.234 | 0.005 | 0.003 | 0.287 |
| Gene | Enzyme | Treatment * | p-Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No | Plus | SEM | BMD | CRP0.5 | CRP1 | LBP0.5 | LBP1 | SEM | Enzyme | Treatment | Enz * Treat | |
| cepA | 3.2 b | 3.3 a | 2.52 | 3.3 a | 3.1 b | 3.3 a | 3.3 a | 3.2 ab | 2.72 | 0.005 | 0.031 | 0.994 |
| lnuAN2 | 3.4 b | 3.6 a | 2.82 | 3.4 bc | 3.5 b | 3.8 a | 3.2 d | 3.4 bc | 3.02 | 0.023 | 0.030 | 0.866 |
| lnuD | 1.8 b | 2.1 a | 1.55 | 2.0 bc | 2.2 a | 1.3 d | 1.7 c | 2.1 ab | 1.75 | 0.018 | 0.050 | 0.953 |
| ermG | 5.0 | 5.0 | 3.94 | 5.1 | 5.0 | 5.0 | 5.0 | 5.1 | 4.14 | 0.058 | 0.104 | 0.968 |
| mefEn2 | 3.6 b | 3.9 a | 3.06 | 3.6 bc | 3.8 ab | 4.0 a | 3.4 d | 3.6 bc | 3.26 | 0.022 | 0.032 | 0.858 |
| MACROLIDE | 4.4 b | 4.6 a | 3.24 | 4.4 a | 4.3 b | 4.4 a | 4.4 a | 4.4 a | 3.44 | 0.012 | 0.028 | 0.997 |
| NBMICROLIDE | 0.9 | 0.9 | 0.51 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | −0.32 | 0.536 | 0.731 | 0.051 |
| sat4 | 4.7 a | 4.6 b | 3.50 | 4.7 | 4.6 | 4.6 | 4.7 | 4.7 | 3.70 | 0.020 | 0.244 | 0.960 |
| tetBP | 4.2 | 4.3 | 3.56 | 3.8 d | 4.3 b | 4.1 bc | 4.3 b | 4.5 a | 3.76 | 0.880 | 0.049 | 0.810 |
| tetQ | 3.9 b | 4.1 a | 3.31 | 4.0 b | 4.0 b | 4.2 a | 4.0 b | 4.0 b | 3.51 | 0.006 | 0.024 | 0.976 |
| tetO | 4.6 | 4.5 | 3.41 | 4.6 | 4.5 | 4.5 | 4.5 | 4.6 | 3.61 | 0.098 | 0.155 | 0.922 |
| tetAP | 4.0 | 3.9 | 3.22 | 3.5 cd | 4.0 ab | 3.9 bc | 3.6 c | 4.2 a | 3.42 | 0.480 | 0.033 | 0.831 |
| TETRACYCLINE | 4.4 | 4.4 | 3.21 | 4.4 b | 4.4 b | 4.4 b | 4.4 b | 4.5 a | 3.41 | 0.088 | 0.003 | 0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kithama, M.; Hassan, Y.I.; Yin, X.; Tang, J.; Clairmont, L.; Sienkiewicz, O.; Ross, K.; Lau, C.H.-F.; Lepp, D.; Zhao, X.; et al. Shift of Microbiota and Modulation of Resistome in the Ceca of Broiler Chicken Fed Berry Pomace Alone or in Combination of a Multienzyme Mixture. Microorganisms 2025, 13, 1044. https://doi.org/10.3390/microorganisms13051044
Kithama M, Hassan YI, Yin X, Tang J, Clairmont L, Sienkiewicz O, Ross K, Lau CH-F, Lepp D, Zhao X, et al. Shift of Microbiota and Modulation of Resistome in the Ceca of Broiler Chicken Fed Berry Pomace Alone or in Combination of a Multienzyme Mixture. Microorganisms. 2025; 13(5):1044. https://doi.org/10.3390/microorganisms13051044
Chicago/Turabian StyleKithama, Munene, Yousef I. Hassan, Xianhua Yin, Joshua Tang, Lindsey Clairmont, Olimpia Sienkiewicz, Kelly Ross, Calvin Ho-Fung Lau, Dion Lepp, Xin Zhao, and et al. 2025. "Shift of Microbiota and Modulation of Resistome in the Ceca of Broiler Chicken Fed Berry Pomace Alone or in Combination of a Multienzyme Mixture" Microorganisms 13, no. 5: 1044. https://doi.org/10.3390/microorganisms13051044
APA StyleKithama, M., Hassan, Y. I., Yin, X., Tang, J., Clairmont, L., Sienkiewicz, O., Ross, K., Lau, C. H.-F., Lepp, D., Zhao, X., Kiarie, E. G., & Diarra, M. S. (2025). Shift of Microbiota and Modulation of Resistome in the Ceca of Broiler Chicken Fed Berry Pomace Alone or in Combination of a Multienzyme Mixture. Microorganisms, 13(5), 1044. https://doi.org/10.3390/microorganisms13051044