
Molecular Analysis of Indole and Skatole Decomposition Metabolism in Acinetobacter piscicola p38 Utilizing Biochemical and Omics Approaches

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Skatole by High-Performance Liquid Chromatography (HPLC)
2.3. Screening of Skatole Biodegrading Bacteria Using Skatole as the Sole Carbon Source
2.4. Bacterial Identification and Phylogenetic Analysis
2.5. Whole Genome Sample Preparation and Analysis
2.6. Prokaryotic Transcriptome Sample Preparation and Analysis
2.7. Heterologous Expression of Genes and Degradation of Indole and Skatole
2.8. Enzyme Analysis and Detection of Reaction Products
2.9. Protein Phylogenetic Tree Analysis
2.10. Data Analysis
3. Results and Discussion
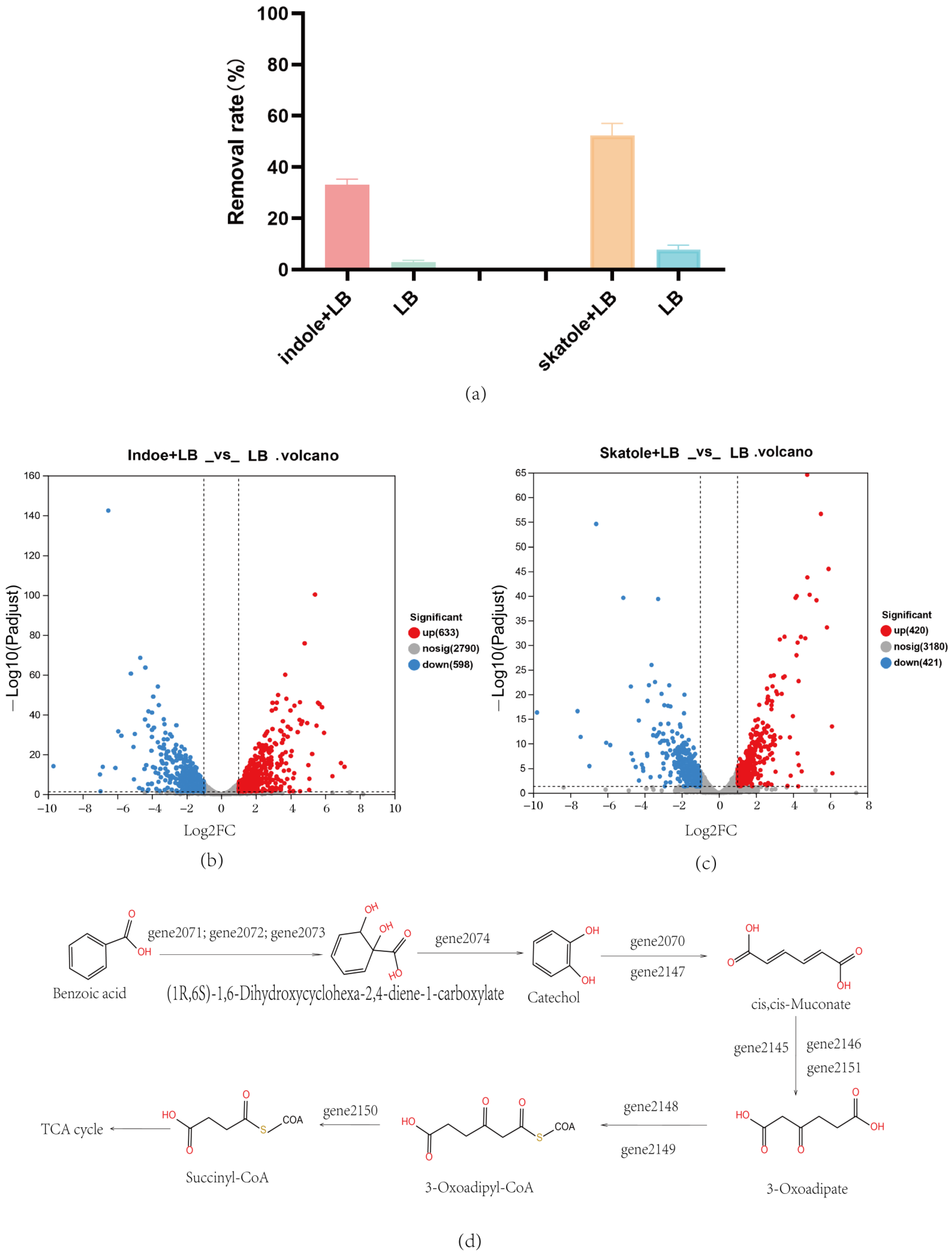
3.1. Screening of Skatole-Degrading Bacteria
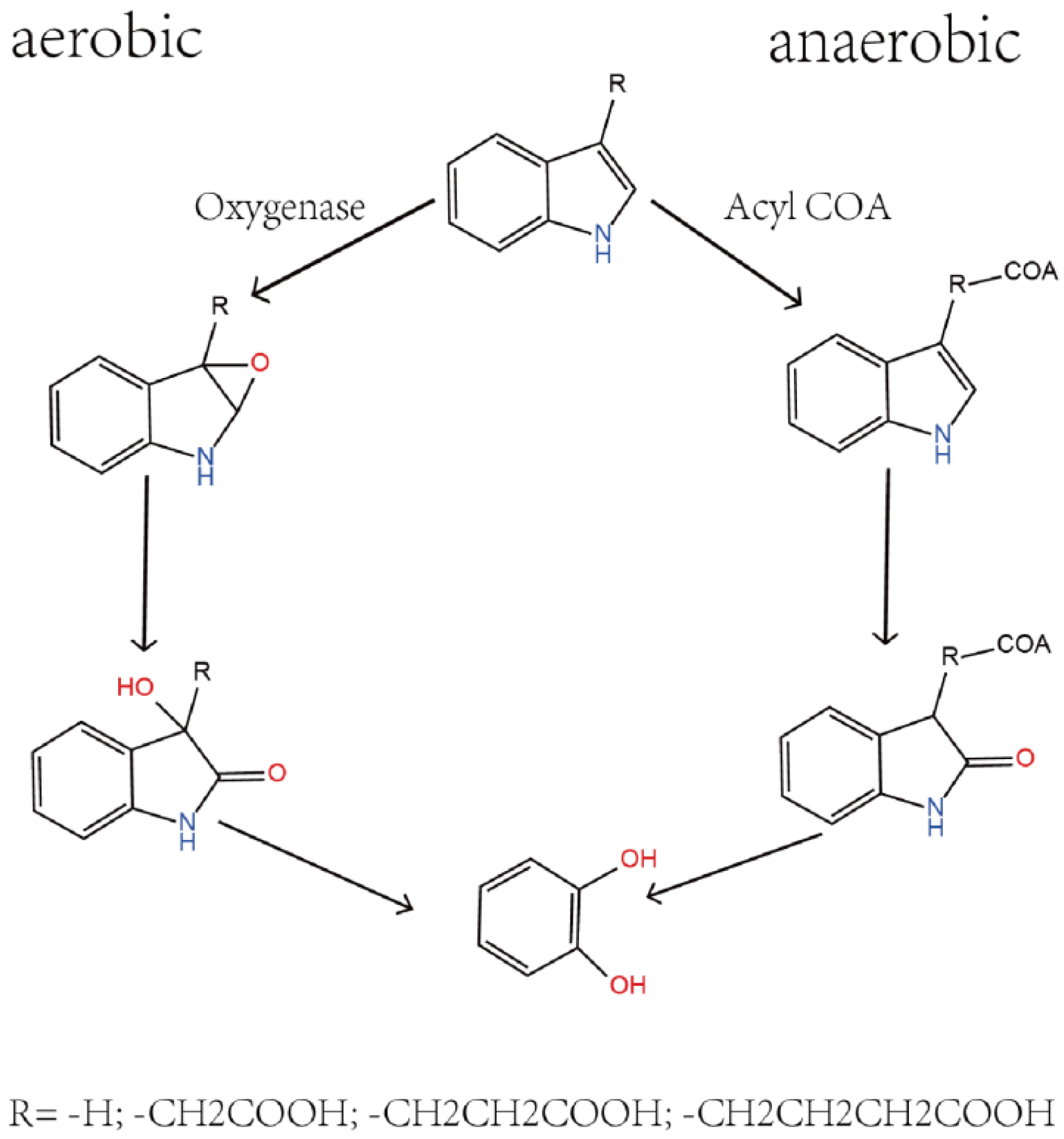
3.2. Metabolite Detection of Metabolism with Skatole as the Sole Carbon Source
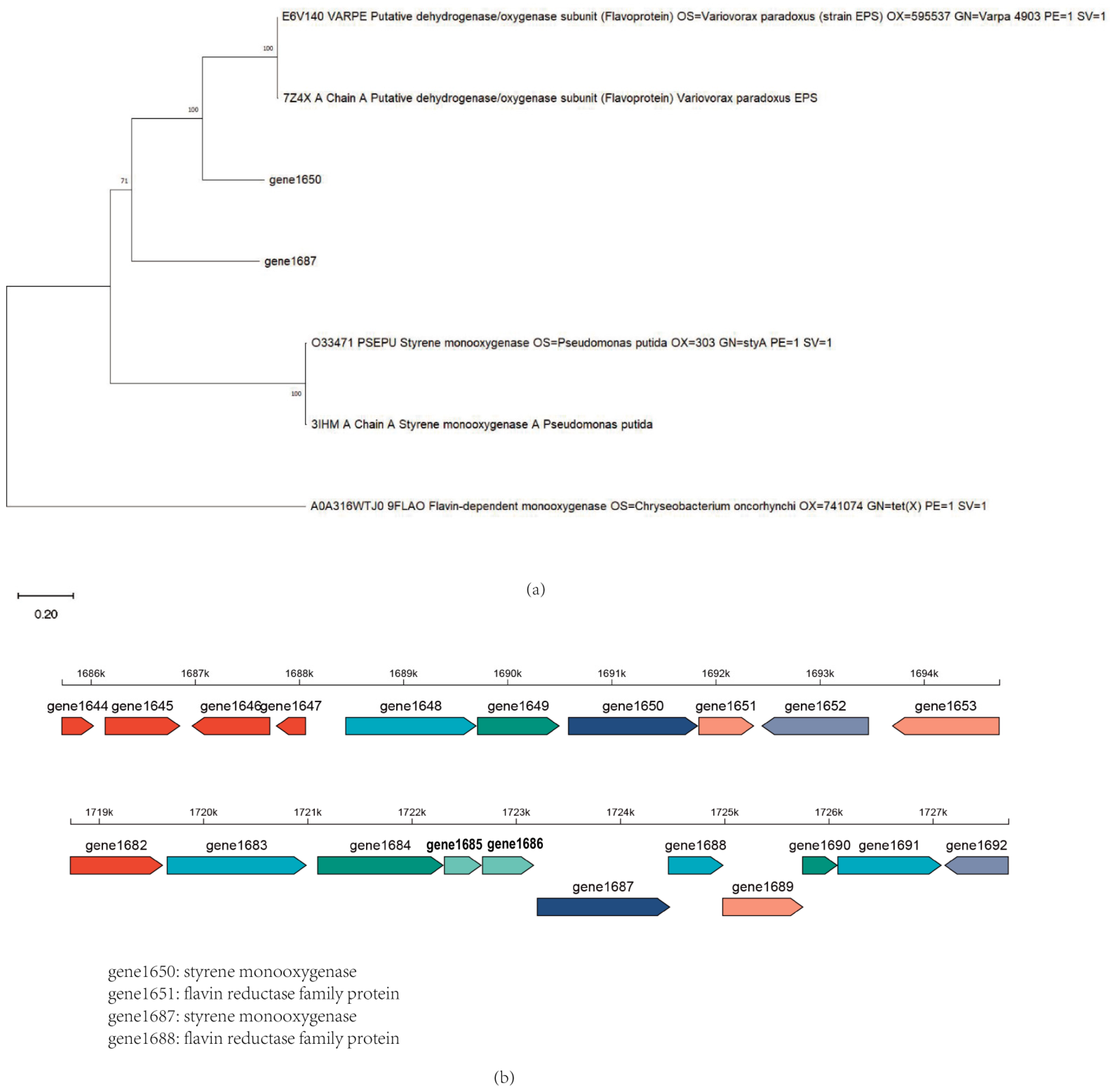
3.3. Whole Genome and Potential Gene Analysis of Acinetobacter_Piscicola p38
3.4. Prokaryotic Transcriptome Analysis of Acinetobacter_Piscicola p38
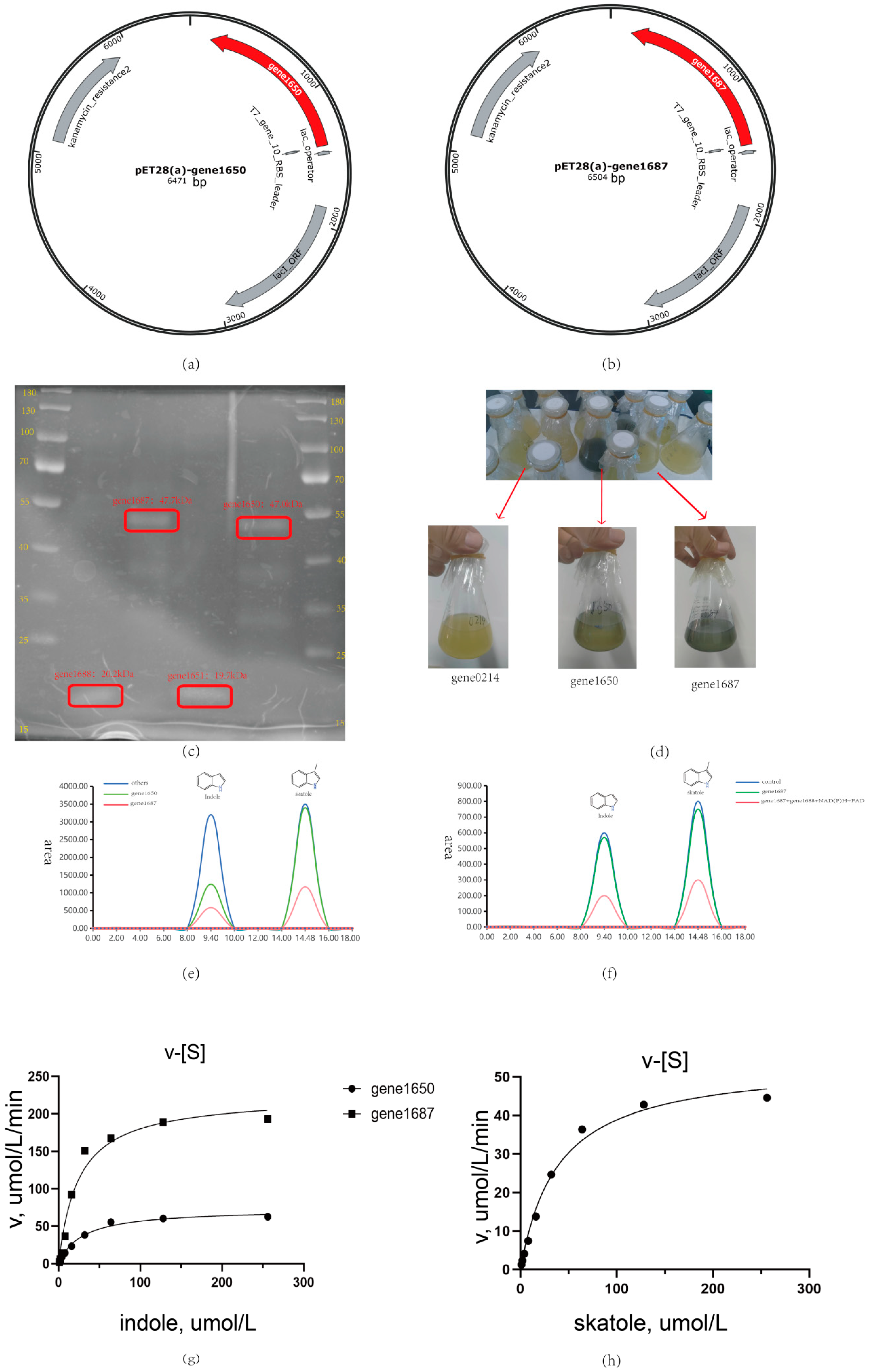
3.5. Heterologous Expression Analysis of Oxygenases and Acyl-Coenzyme a in Acinetobacter_Piscicola p38
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, J.; Aoll, J.; Niclass, Y.; Velazco, M.I.; Wünsche, L.; Pika, J.; Starkenmann, C. Qualitative and quantitative analysis of volatile constituents from latrines. Environ. Sci. Technol. 2013, 47, 7876–7882. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gallagher, D.L.; Dietrich, A.M.; Su, M.; Wang, Q.; Guo, Q.; Zhang, J.; An, W.; Yu, J.; Yang, M. Data Analytics Determines Co-occurrence of Odorants in Raw Water and Evaluates Drinking Water Treatment Removal Strategies. Environ. Sci. Technol. 2021, 55, 16770–16782. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hallis, S.A.; Vitko, T.; Suffet, I.H. Identification, quantification and treatment of fecal odors released into the air at two wastewater treatment plants. J. Environ. Manag. 2016, 180, 257–263. [Google Scholar] [CrossRef]
- Zhang, D.; Ji, H.; Wang, S.; Liu, Y.; Chen, M.; Liu, H. Lactobacillus-driven feed fermentation regulates microbiota metabolism and reduces odor emission from the feces of pigs. mSystems 2023, 8, e0098823. [Google Scholar] [CrossRef] [PubMed]
- Douglas, P.; Robertson, S.; Gay, R.; Hansell, A.L.; Gant, T.W. A systematic review of the public health risks of bioaerosols from intensive farming. Int. J. Hyg. Environ. Health 2018, 221, 134–173. [Google Scholar] [CrossRef] [PubMed]
- Lebrero, R.; Bouchy, L.; Stuetz, R.; Muñoz, R. Odor Assessment and Management in Wastewater Treatment Plants: A Review. Crit. Rev. Environ. Sci. Technol. 2011, 41, 915–950. [Google Scholar] [CrossRef]
- Ma, Q.; Zhang, X.; Qu, Y. Biodegradation and Biotransformation of Indole: Advances and Perspectives. Front. Microbiol. 2018, 9, 2625. [Google Scholar] [CrossRef]
- Ma, Q.; Meng, N.; Li, Y.; Wang, J. Occurrence, impacts, and microbial transformation of 3-methylindole (skatole): A critical review. J. Hazard. Mater. 2021, 416, 126181. [Google Scholar] [CrossRef]
- Yin, K.; Wang, Q.; Lv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Arora, P.K.; Sharma, A.; Bae, H. Microbial degradation of indole and its derivatives. J. Chem. 2015, 2015, 129159. [Google Scholar] [CrossRef]
- Du, H.; Cheng, J.L.; Li, Z.Y.; Zhong, H.N.; Wei, S.; Gu, Y.J.; Yao, C.C.; Zhang, M.; Cai, Q.Y.; Zhao, H.M.; et al. Molecular insights into the catabolism of dibutyl phthalate in Pseudomonas aeruginosa PS1 based on biochemical and multi-omics approaches. Sci. Total Environ. 2024, 926, 171852. [Google Scholar] [CrossRef]
- Ma, Q.; Liu, Z.; Yang, B.; Dai, C.; Qu, Y. Characterization and functional gene analysis of a newly isolated indole-degrading bacterium Burkholderia sp. IDO3. J. Hazard. Mater. 2019, 367, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Laird, T.S.; Flores, N.; Leveau, J.H.J. Bacterial catabolism of indole-3-acetic acid. Appl. Microbiol. Biotechnol. 2020, 104, 9535–9550. [Google Scholar] [CrossRef] [PubMed]
- Sadauskas, M.; Statkevičiūtė, R.; Vaitekūnas, J.; Meškys, R. Bioconversion of Biologically Active Indole Derivatives with Indole-3-Acetic Acid-Degrading Enzymes from Caballeronia glathei DSM50014. Biomolecules 2020, 10, 663. [Google Scholar] [CrossRef]
- Fujioka, M.; Wada, H. The bacterial oxidation of indole. Biochim. Biophys. Acta 1968, 158, 70–78. [Google Scholar] [CrossRef]
- Sadauskas, M.; Vaitekūnas, J.; Gasparavičiūtė, R.; Meškys, R. Indole Biodegradation in Acinetobacter sp. Strain O153: Genetic and Biochemical Characterization. Appl. Environ. Microbiol. 2017, 83, e01453-17. [Google Scholar] [CrossRef]
- Lin, G.H.; Chen, H.P.; Huang, J.H.; Liu, T.T.; Lin, T.K.; Wang, S.J.; Tseng, C.H.; Shu, H.Y. Identification and characterization of an indigo-producing oxygenase involved in indole 3-acetic acid utilization by Acinetobacter baumannii. Antonie van Leeuwenhoek 2012, 101, 881–890. [Google Scholar] [CrossRef]
- Donoso, R.; Leiva-Novoa, P.; Zúñiga, A.; Timmermann, T.; Recabarren-Gajardo, G.; González, B. Biochemical and Genetic Bases of Indole-3-Acetic Acid (Auxin Phytohormone) Degradation by the Plant-Growth-Promoting Rhizobacterium Paraburkholderia phytofirmans PsJN. Appl. Environ. Microbiol. 2016, 83, e01991-16. [Google Scholar] [CrossRef]
- Greenhut, I.V.; Slezak, B.L.; Leveau, J.H.J. Iac Gene Expression in the Indole-3-Acetic Acid-Degrading Soil Bacterium Enterobacter soli LF7. Appl. Environ. Microbiol. 2018, 84, e01057-18. [Google Scholar] [CrossRef]
- Leveau, J.H.; Lindow, S.E. Utilization of the plant hormone indole-3-acetic acid for growth by Pseudomonas putida strain 1290. Appl. Environ. Microbiol. 2005, 71, 2365–2371. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.J.; Hu, J.Y.; Han, X.Y.; Li, Y.; Luo, X.C.; Wang, Z.L.; Li, J.Z. Degradation of indole via a two-component indole oxygenase system from Enterococcus hirae GDIAS-5. J. Hazard. Mater. 2023, 458, 131707. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Ma, Q.; Liu, Z.; Wang, W.; Tang, H.; Zhou, J.; Xu, P. Unveiling the biotransformation mechanism of indole in a Cupriavidus sp. strain. Mol. Microbiol. 2017, 106, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, L.; Gao, F.; Diao, W.; Ma, H.; Feng, F.; Quan, S.; Xiang, L.; Zhang, X. Screening, identification, and degradation characteristics of 3-methylindole degrading bacteria. Front. Environ. Sci. 2022, 10, 1028699. [Google Scholar] [CrossRef]
- Meng, X.; He, Z.F.; Li, H.J.; Zhao, X. Removal of 3-methylindole by lactic acid bacteria in vitro. Exp. Ther. Med. 2013, 6, 983–988. [Google Scholar] [CrossRef]
- Tesso, T.A.; Zheng, A.; Cai, H.; Liu, G. Isolation and characterization of two Acinetobacter species able to degrade 3-methylindole. PLoS ONE 2019, 14, e0211275. [Google Scholar] [CrossRef]
- Fukuoka, K.; Ozeki, Y.; Kanaly, R.A. Aerobic biotransformation of 3-methylindole to ring cleavage products by Cupriavidus sp. strain KK10. Biodegradation 2015, 26, 359–373. [Google Scholar] [CrossRef]
- Li, P.; Tong, L.; Liu, K.; Wang, Y. Biodegradation of 3-methylindole by Pseudomonas putida LPC24 under oxygen limited conditions. Fresenius Environ. Bull. 2010, 19, 238–242. [Google Scholar]
- Yin, B.; Gu, J.D. Aerobic Degradation of 3-Methylindole by Pseudomonas aeruginosa Gs Isolated from Mangrove Sediment. Hum. Ecol. Risk Assess. 2006, 12, 248–258. [Google Scholar] [CrossRef]
- Gu, J.D.; Fan, Y.; Shi, H. Relationship between structures of substituted indolic compounds and their degradation by marine anaerobic microorganisms. Mar. Pollut. Bull. 2002, 45, 379–384. [Google Scholar] [CrossRef]
- Dai, C.; Ma, F.; Ma, Q.; Yang, J.; Li, Y.; Yang, B.; Qu, Y. Investigation of indole biodegradation by Cupriavidus sp. strain IDO with emphases on downstream biotransformation and indigo production. Environ. Sci. Pollut. Res. Int. 2022, 29, 8369–8381. [Google Scholar] [CrossRef]
- Ma, Q.; Meng, N.; Su, J.; Li, Y.; Gu, J.; Wang, Y.; Wang, J.; Qu, Y.; Zhao, Z.; Sun, Y. Unraveling the skatole biodegradation process in an enrichment consortium using integrated omics and culture-dependent strategies. J. Environ. Sci. 2023, 127, 688–699. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cow | Removal Rate | Chicken | Removal Rate | Sheep | Removal Rate | Pig | Removal Rate |
|---|---|---|---|---|---|---|---|
| 1 | 18.51% | 11 | 1.89% | 21 | 0.52% | 31 | 38.06% |
| 2 | 0.50% | 12 | 9.39% | 22 | none | 32 | none |
| 3 | 2.47% | 13 | none | 23 | none | 33 | 1.39% |
| 4 | 19.38% | 14 | none | 24 | 5.45% | 34 | none |
| 5 | 0.76% | 15 | 35.74% | 25 | none | 35 | 100.00% |
| 6 | none | 16 | 12.75% | 26 | 8.79% | 36 | 0.39% |
| 7 | none | 17 | 86.34% | 27 | 4.11% | 37 | none |
| 8 | none | 18 | 41.84% | 28 | 3.28% | 38 | 100.00% |
| 9 | none | 19 | 5.73% | 29 | none | 39 | none |
| 10 | none | 20 | none | 30 | 1.60% | 40 | none |
| Gene_Id | Gene Name | Gene Description |
|---|---|---|
| gene0215 | sfnG | MULTISPECIES: dimethyl sulfone monooxygenase SfnG |
| gene0321 | - | MULTISPECIES: DOPA 4,5-dioxygenase family protein |
| gene1650 | styA | styrene monooxygenase |
| gene1684 | - | aromatic ring-hydroxylating dioxygenase subunit alpha |
| gene2073 | benC-xylZ | benzoate 1,2-dioxygenase electron transfer component BenC |
| gene2147 | catA | MULTISPECIES: catechol 1,2-dioxygenase |
| gene2332 | npd | MULTISPECIES: nitronate monooxygenase |
| gene2700 | tauD | MULTISPECIES: taurine dioxygenase |
| gene3162 | mhpB | MULTISPECIES: 3-carboxyethylcatechol 2,3-dioxygenase |
| gene3524 | ssuD | FMNH2-dependent alkanesulfonate monooxygenase |
| gene0214 | dszC | acyl-CoA dehydrogenase family protein |
| gene0703 | dmdC | MULTISPECIES: acyl-CoA dehydrogenase C-terminal domain-containing protein |
| gene0704 | dmdC | MULTISPECIES: acyl-CoA dehydrogenase C-terminal domain-containing protein |
| gene1225 | desA3 | MULTISPECIES: acyl-CoA desaturase |
| gene2011 | bcd | MULTISPECIES: acyl-CoA dehydrogenase family protein |
| gene2462 | ybgC | MULTISPECIES: tol-pal system-associated acyl-CoA thioesterase |
| gene2913 | atuD | MULTISPECIES: acyl-CoA dehydrogenase family protein |
| gene2914 | atuC | MULTISPECIES: acyl-CoA carboxylase subunit beta |
| gene2970 | dmdB | MULTISPECIES: acyl-CoA synthetase |
| gene3178 | atuH | MULTISPECIES: long-chain-acyl-CoA synthetase |
| Gene_Id | Gene Name | Gene Description |
|---|---|---|
| gene0215 | sfnG | MULTISPECIES: dimethyl sulfone monooxygenase SfnG |
| gene1684 | - | aromatic ring-hydroxylating dioxygenase subunit alpha |
| gene1687 | - | MULTISPECIES: styrene monooxygenase |
| gene1690 | hcaC | MULTISPECIES: non-heme iron oxygenase ferredoxin subunit |
| gene2070 | catA | MULTISPECIES: catechol 1,2-dioxygenase |
| gene2071 | benA-xylX | MULTISPECIES: benzoate 1,2-dioxygenase large subunit |
| gene2072 | benB-xylY | MULTISPECIES: benzoate 1,2-dioxygenase small subunit |
| gene2073 | benC-xylZ | benzoate 1,2-dioxygenase electron transfer component BenC |
| gene2147 | catA | MULTISPECIES: catechol 1,2-dioxygenase |
| gene2332 | npd | MULTISPECIES: nitronate monooxygenase |
| gene2700 | tauD | MULTISPECIES: taurine dioxygenase |
| gene3524 | ssuD | FMNH2-dependent alkanesulfonate monooxygenase |
| gene0214 | dszC | acyl-CoA dehydrogenase family protein |
| gene0660 | ybgC | MULTISPECIES: acyl-CoA thioesterase |
| gene0703 | dmdC | MULTISPECIES: acyl-CoA dehydrogenase C-terminal domain-containing protein |
| gene0704 | dmdC | MULTISPECIES: acyl-CoA dehydrogenase C-terminal domain-containing protein |
| gene1225 | desA3 | MULTISPECIES: acyl-CoA desaturase |
| gene1970 | paaH | 3-hydroxyacyl-CoA dehydrogenase |
| gene2011 | bcd | MULTISPECIES: acyl-CoA dehydrogenase family protein |
| gene3178 | atuH | MULTISPECIES: long-chain-acyl-CoA synthetase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Sun, J.; Yang, P.; Zhang, W.; Jiang, Y.; Liu, Q.; Yang, Y.; Hao, R.; Guo, G.; Huo, W.; et al. Molecular Analysis of Indole and Skatole Decomposition Metabolism in Acinetobacter piscicola p38 Utilizing Biochemical and Omics Approaches. Microorganisms 2024, 12, 1792. https://doi.org/10.3390/microorganisms12091792
Wang Z, Sun J, Yang P, Zhang W, Jiang Y, Liu Q, Yang Y, Hao R, Guo G, Huo W, et al. Molecular Analysis of Indole and Skatole Decomposition Metabolism in Acinetobacter piscicola p38 Utilizing Biochemical and Omics Approaches. Microorganisms. 2024; 12(9):1792. https://doi.org/10.3390/microorganisms12091792
Chicago/Turabian StyleWang, Zhonghao, Jiajin Sun, Pu Yang, Wanjun Zhang, Yihong Jiang, Qiang Liu, Yunqi Yang, Ruirong Hao, Gang Guo, Wenjie Huo, and et al. 2024. "Molecular Analysis of Indole and Skatole Decomposition Metabolism in Acinetobacter piscicola p38 Utilizing Biochemical and Omics Approaches" Microorganisms 12, no. 9: 1792. https://doi.org/10.3390/microorganisms12091792
APA StyleWang, Z., Sun, J., Yang, P., Zhang, W., Jiang, Y., Liu, Q., Yang, Y., Hao, R., Guo, G., Huo, W., Zhang, Q., & Li, Q. (2024). Molecular Analysis of Indole and Skatole Decomposition Metabolism in Acinetobacter piscicola p38 Utilizing Biochemical and Omics Approaches. Microorganisms, 12(9), 1792. https://doi.org/10.3390/microorganisms12091792