The Impact of Immune System Aging on Infectious Diseases


Abstract
1. Introduction
2. Aging of Immune System Components in Physiological Conditions
2.1. Age-Associated Changes in the Immune Compartments
2.2. Sex-Differences in Innate and Adaptive Immunity Increase with Age
2.3. Immune System Complexity Increases with Age
2.4. Biomarkers of Aged Immune System
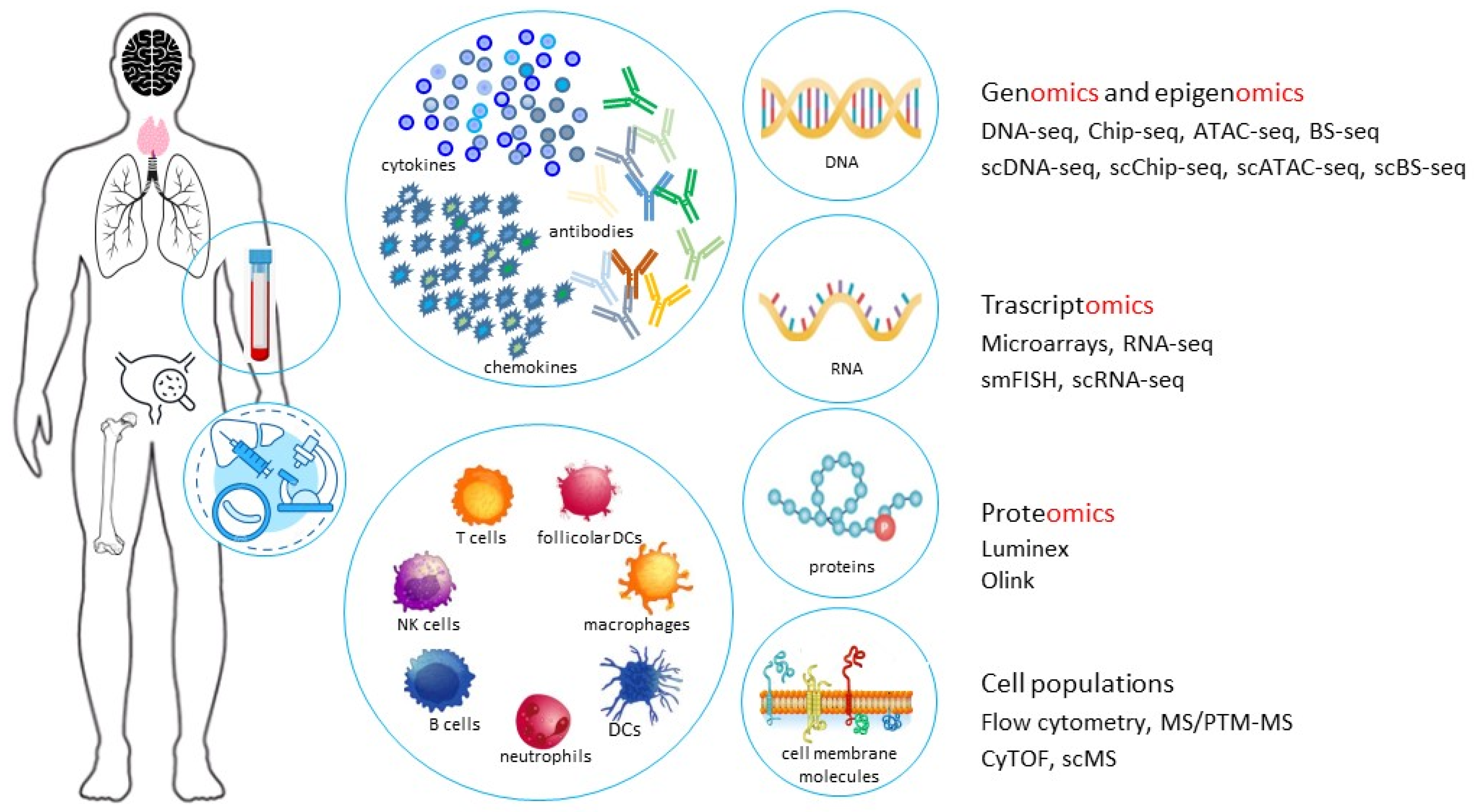
2.5. The Systems Immunology for the Study of Age-Related Immune System Variations
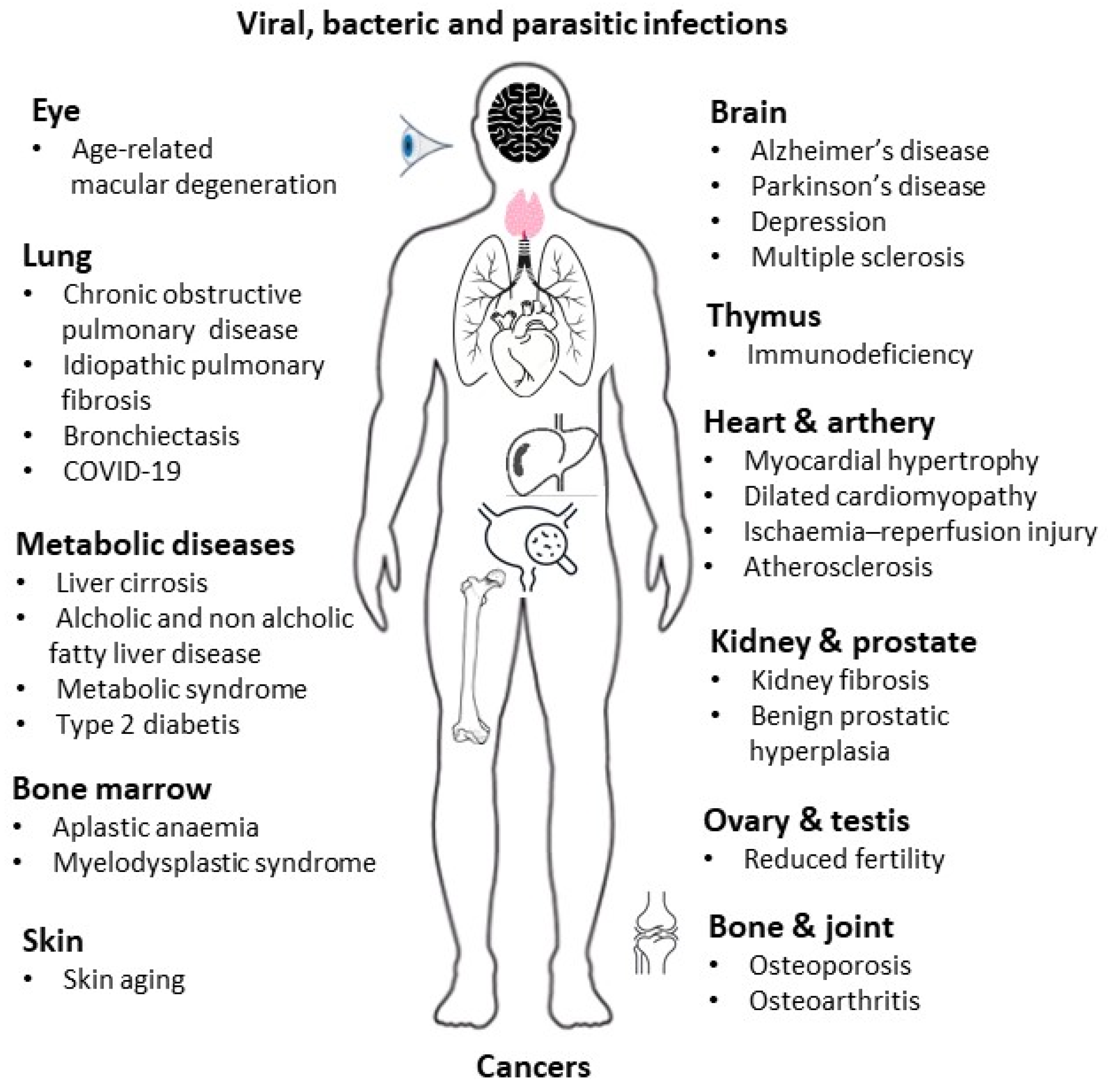
3. Aging of Immune System Components in Infectious Diseases
3.1. Viral Infections
3.2. Bacterial Infections
3.3. Parasitic Infections
4. Immunosenescence and Vaccines
5. Limitations of Current Research on Immunosenescence, Future Research Directions and Potential Therapeutic Interventions
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Dong, C.; Han, Y.; Gu, Z.; Sun, C. Immunosenescence, aging and successful aging. Front. Immunol. 2022, 13, 942796. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.R. (Ed.) Evolutionary Biology of Aging. Oxford University Press: New York, NY, USA, 1991; ISBN 978-01-9509-530-2. [Google Scholar]
- Kirkwood, T.B.; Austad, S.N. Why do we age? Nature 2000, 408, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Saborido, C.; García-Barranquero, P. Is aging a disease? The theoretical definition of aging in the light of the philosophy of medicine. J. Med. Philos. 2022, 47, 770–783. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in healthy aging and disease. Nat. Aging 2021, 1, 634–650. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Tuttle CS, L.; Waaijer ME, C.; Slee-Valentijn, M.S.; Stijnen, T.; Westendorp, R.; Maier, A.B. Cellular senescence and chronological age in various human tissues: A systematic review and meta-analysis. Aging Cell 2020, 19, e13083. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, M.; Song, W.M.; Wang, Q.; Yuan, G.C.; Sudmant, P.H.; Zare, H.; Tu, Z.; Orr, M.E.; Zhang, B. The landscape of human tissue and cell type specific expression and co-regulation of senescence genes. Mol. Neurodegener. 2022, 1, 5. [Google Scholar] [CrossRef]
- Low, E.; Alimohammadiha, G.; Smith, L.A.; Costello, L.F.; Przyborski, S.A.; von Zglinicki, T.; Miwa, S. How good is the evidence that cellular senescence causes skin ageing? Ageing Res. Rev. 2021, 71, 101456. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Li, T.; Xu, H.; Zhang, H. Senescence in osteoarthritis: From mechanism to potential treatment. Arthritis Res. Ther. 2022, 24, 174. [Google Scholar] [CrossRef]
- Butcher, S.K.; Wang, K.; Lascelles, D.; Lord, J.M. Neutrophil ageing and immunosenescence. NeuroImmune Biol. 2004, 4, 41–55. [Google Scholar] [CrossRef]
- Aw, D.; Silva, A.B.; Palmer, D.B. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Bolton, C. An evaluation of the recognised systemic inflammatory biomarkers of chronic sub-optimal inflammation provides evidence for inflammageing (IFA) during multiple sclerosis (MS). Immun. Ageing 2021, 18, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liang, Q.; Ren, Y.; Guo, C.; Ge, X.; Wang, L.; Cheng, Q.; Luo, P.; Zhang, Y.; Han, X. Immunosenescence: Molecular mechanisms and diseases. Signal Transduct. Target. Ther. 2023, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Marchegiani, F.; Matacchione, G.; Giuliani, A.; Ramini, D.; Fazioli, F.; Sabbatinelli, J.; Bonafè, M. Sex/gender-related differences in inflammaging. Mech. Ageing Dev. 2023, 211, 111792. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Coppé, J.P.; Lam, E.W. Cellular senescence: The sought or the unwanted? Trends Mol. Med. 2018, 24, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An Update on inflamm-aging: Mechanisms, prevention, and treatment. J. Immunol. Res. 2016, 2016, 8426874. [Google Scholar] [CrossRef]
- Rossiello, F.; Jurk, D.; Passos, J.F.; d’Adda di Fagagna, F. Telomere dysfunction in ageing and age-related diseases. Nat. Cell Biol. 2022, 24, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Fraile-Martinez, O.; De Leon-Oliva, D.; Boaru, D.L.; De Castro-Martinez, P.; Garcia-Montero, C.; Barrena-Blázquez, S.; García-García, J.; García-Honduvilla, N.; Alvarez-Mon, M.; Lopez-Gonzalez, L.; et al. Connecting epigenetics and inflammation in vascular senescence: State of the art, biomarkers and senotherapeutics. Front. Genet. 2024, 15, 1345459. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Martin-Ruiz, C.M.; Takayama, M.; Abe, Y.; Takebayashi, T.; Koyasu, S.; Suematsu, M.; Hirose, N.; von Zglinicki, T. Inflammation, but not telomere length, predicts successful ageing at extreme old age: A longitudinal study of semi-supercentenarians. EBioMedicine 2015, 2, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and inflamm-aging as two sides of the same coin: Friends or foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Bientinesi, E.; Monti, D. Immunosenescence and inflammaging in the aging process: Age-related diseases or longevity? Ageing Res. Rev. 2021, 71, 101422. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Pawelec, G.; Khalil, A.; Cohen, A.A.; Hirokawa, K.; Witkowski, J.M.; Franceschi, C. Immunology of aging: The birth of inflammaging. Clin. Rev. Allergy Immunol. 2023, 64, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Human T cell immunosenescence and inflammation in aging. J. Leukoc. Biol. 2017, 102, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G. Age and immunity: What is “immunosenescence”? Exp. Gerontol. 2018, 105, 4–9. [Google Scholar] [CrossRef]
- Chinn, I.K.; Blackburn, C.C.; Manley, N.R.; Sempowski, G.D. Changes in primary lymphoid organs with aging. Semin. Immunol. 2012, 24, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.P.; Teixeira, V.R.; Alencar-Silva, T.; Simonassi-Paiva, B.; Pereira, R.W.; Pogue, R.; Carvalho, J.L. Hallmarks of aging and immunosenescence: Connecting the dots. Cytokine Growth Factor. Rev. 2021, 59, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Chatta, G.S.; Andrews, R.G.; Rodger, E.; Schrag, M.; Hammond, W.P.; Dale, D.C. Hematopoietic progenitors and aging: Alterations in granulocytic precursors and responsiveness to recombinant human G-CSF, GM-CSF, and IL-3. J. Gerontol. 1993, 48, M207–M212. [Google Scholar] [CrossRef]
- Sabbatini, M.; Bona, E.; Novello, G.; Migliario, M.; Renò, F. Aging hampers neutrophil extracellular traps (NETs) efficacy. Aging Clin. Exp. Res. 2022, 34, 2345–2353. [Google Scholar] [CrossRef]
- van Beek, A.A.; Van den Bossche, J.; Mastroberardino, P.G.; de Winther, M.P.J.; Leenen, P.J.M. Metabolic alterations in aging Macrophages: Ingredients for inflammaging? Trends Immunol. 2019, 40, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Taabazuing, C.Y.; Okondo, M.C.; Bachovchin, D.A. Pyroptosis and apoptosis pathways engage in bidirectional crosstalk in monocytes and macrophages. Cell Chem. Biol. 2017, 24, 507–514.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Qiu, J.; Song, Y.; Liang, T.; Liu, S.; Ren, C.; Song, X.; Cui, L.; Sun, Y. Pyroptosis and degenerative diseases of the elderly. Cell Death Dis. 2023, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Gounder, S.S.; Abdullah, B.J.J.; Radzuanb, N.E.I.B.M.; Zain, F.D.B.M.; Sait, N.B.M.; Chua, C.; Subramani, B. Effect of aging on NK cell population and their proliferation at ex vivo culture condition. Anal. Cell. Pathol. 2018, 2018, 7871814. [Google Scholar] [CrossRef] [PubMed]
- Douek, D.C.; McFarland, R.D.; Keiser, P.H.; Gage, E.A.; Massey, J.M.; Haynes, B.F.; Polis, M.A.; Haase, A.T.; Feinberg, M.B.; Sullivan, J.L.; et al. Changes in thymic function with age and during the treatment of HIV infection. Nature 1998, 396, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Gulla, S.; Reddy, M.C.; Reddy, V.C.; Chitta, S.; Bhanoori, M.; Lomada, D. Role of thymus in health and disease. Int. Rev. Immunol. 2023, 42, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Bains, I.; Yates, A.J.; Callard, R.E. Heterogeneity in thymic emigrants: Implications for thymectomy and immunosenescence. PLoS ONE 2013, 8, e49554. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.A.; Helm, E.Y.; Fink, P.J. Reinterpreting recent thymic emigrant function: Defective or adaptive? Curr. Opin. Immunol. 2018, 51, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Prat, M.; Álvarez-Sierra, D.; Aguiló-Cucurull, A.; Salgado-Perandrés, S.; Briongos-Sebastian, S.; Franco-Jarava, C.; Martin-Nalda, A.; Colobran, R.; Montserrat, I.; Hernández-González, M.; et al. Extended immunophenotyping reference values in a healthy pediatric population. Cytom. B Clin. Cytom. 2019, 96, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Steffens, C.M.; Al-Harthi, L.; Shott, S.; Yogev, R.; Landay, A. Evaluation of thymopoiesis using T cell receptor excision circles (TRECs): Differential correlation between adult and pediatric TRECs and naïve phenotypes. Clin. Immunol. 2000, 97, 95–101. [Google Scholar] [CrossRef]
- Sottini, A.; Serana, F.; Bertoli, D.; Chiarini, M.; Valotti, M.; Vaglio Tessitore, M.; Imberti, L. Simultaneous quantification of T-cell receptor excision circles (TRECs) and K-deleting recombination excision circles (KRECs) by real-time PCR. J. Vis. Exp. 2014, 94, 52184. [Google Scholar] [CrossRef]
- Adams, S.P.; Kricke, S.; Ralph, E.; Gilmour, N.; Gilmour, K.C. A comparison of TRECs and flow cytometry for naive T cell quantification. Clin. Exp. Immunol. 2018, 191, 198–202. [Google Scholar] [CrossRef]
- Weng, N.P. Numbers and odds: TCR repertoire size and its age changes impacting on T cell functions. Semin. Immunol. 2023, 69, 101810. [Google Scholar] [CrossRef]
- Qi, Q.; Liu, Y.; Cheng, Y.; Glanville, J.; Zhang, D.; Lee, J.Y.; Olshen, R.A.; Weyand, C.M.; Boyd, S.D.; Goronzy, J.J. Diversity and clonal selection in the human T-cell repertoire. Proc. Natl. Acad. Sci. USA 2014, 111, 13139–13144. [Google Scholar] [CrossRef] [PubMed]
- Derhovanessian, E.; Maier, A.B.; Beck, R.; Jahn, G.; Hähnel, K.; Slagboom, P.E.; de Craen, A.J.; Westendorp, R.G.; Pawelec, G. Hallmark features of immunosenescence are absent in familial longevity. J. Immunol. 2010, 185, 4618–4624. [Google Scholar] [CrossRef] [PubMed]
- Britanova, O.V.; Shugay, M.; Merzlyak, E.M.; Staroverov, D.B.; Putintseva, E.V.; Turchaninova, M.A.; Mamedov, I.Z.; Pogorelyy, M.V.; Bolotin, D.A.; Izraelson, M.; et al. Dynamics of individual T cell repertoires: From cord blood to centenarians. J. Immunol. 2016, 196, 5005–5013. [Google Scholar] [CrossRef] [PubMed]
- Tessitore, M.V.; Sottini, A.; Roccaro, A.M.; Ghidini, C.; Bernardi, S.; Martellosio, G.; Serana, F.; Imberti, L. Detection of newly produced T and B lymphocytes by digital PCR in blood stored dry on nylon flocked swabs. J. Transl. Med. 2017, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Landin, A.M.; Lechner, S.C.; Ryan, J.G.; Schwartz, R.; Riley, R.L.; Blomberg, B.B. Aging down-regulates the transcription factor E2A, activation-induced cytidine deaminase, and Ig class switch in human B cells. J. Immunol. 2008, 180, 5283–5290. [Google Scholar] [CrossRef]
- Dowery, R.; Benhamou, D.; Benchetrit, E.; Harel, O.; Nevelsky, A.; Zisman-Rozen, S.; Braun-Moscovici, Y.; Balbir-Gurman, A.; Avivi, I.; Shechter, A.; et al. Peripheral B cells repress B-cell regeneration in aging through a TNF-α/IGFBP-1/IGF-1 immune-endocrine axis. Blood 2021, 138, 1817–1829. [Google Scholar] [CrossRef]
- Ma, S.; Wang, C.; Mao, X.; Hao, Y.B. Cell dysfunction associated with aging and autoimmune diseases. Front. Immunol. 2019, 10, 318. [Google Scholar] [CrossRef]
- de Mol, J.; Kuiper, J.; Tsiantoulas, D.; Foks, A.C. The dynamics of B cell aging in health and disease. Front. Immunol. 2021, 12, 733566. [Google Scholar] [CrossRef]
- Fulop, T.; Witkowski, J.M.; Le Page, A.; Fortin, C.; Pawelec, G.; Larbi, A. Intracellular signalling pathways: Targets to reverse immunosenescence. Clin. Exp. Immunol. 2017, 187, 35–43. [Google Scholar] [CrossRef]
- Murakami, G.; Taniguchi, I. Histologic heterogeneity and intranodal shunt flow in lymph nodes from elderly subjects: A cadaveric study. Ann. Surg. Oncol. 2004, 11, 279S–284S. [Google Scholar] [CrossRef]
- Erofeeva, L.M.; Mnikhovich, M.V. Structural and functional changes in the mesenteric lymph nodes in humans during aging. Bull. Exp. Biol. Med. 2020, 168, 694–698. [Google Scholar] [CrossRef]
- Cakala-Jakimowicz, M.; Kolodziej-Wojnar, P.; Puzianowska-Kuznicka, M. Aging-related cellular, structural and functional changes in the lymph nodes: A significant component of immunosenescence? An Overview. Cells 2021, 10, 3148. [Google Scholar] [CrossRef]
- Denton, A.; Silva-Cayetano, A.; Dooley, J.; Hill, D.L.; Carr, E.J.; Robert, P.; Meyer-Hermann, M.; Liston, A.; Linterman, M.A. Intrinsic defects in lymph node stromal cells underpin poor germinal center responses during aging. bioRxiv. 2020. [Google Scholar] [CrossRef]
- Bronte, V.; Pittet, M.J. The spleen in local and systemic regulation of immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef]
- Turner, V.M.; Mabbott, N.A. Influence of ageing on the microarchitecture of the spleen and lymph nodes. Biogerontology 2017, 18, 723–738. [Google Scholar] [CrossRef]
- Budamagunta, V.; Foster, T.C.; Zhou, D. Cellular senescence in lymphoid organs and immunosenescence. Aging 2021, 13, 19920–19941. [Google Scholar] [CrossRef]
- Sato, S.; Kiyono, H.; Fujihashi, K. Mucosal Immunosenescence in the gastrointestinal tract: A Mini-Review. Gerontology 2015, 61, 336–342. [Google Scholar] [CrossRef]
- Márquez, E.J.; Chung, C.H.; Marches, R.; Rossi, R.J.; Nehar-Belaid, D.; Eroglu, A.; Mellert, D.J.; Kuchel, G.A.; Banchereau, J.; Ucar, D. Sexual-dimorphism in human immune system aging. Nat. Commun. 2020, 11, 751. [Google Scholar] [CrossRef]
- Bongen, E.; Lucian, H.; Khatri, A.; Fragiadakis, G.K.; Bjornson, Z.B.; Nolan, G.P.; Utz, P.J.; Khatri, P. Sex differences in the blood transcriptome identify robust changes in immune cell proportions with aging and influenza infection. Cell Rep. 2019, 29, 1961–1973. [Google Scholar] [CrossRef]
- Pietschmann, P.; Gollob, E.; Brosch, S.; Hahn, P.; Kudlacek, S.; Willheim, M.; Woloszczuk, W.; Peterlik, M.; Tragl, K.H. The effect of age and gender on cytokine production by human peripheral blood mononuclear cells and markers of bone metabolism. Exp. Gerontol. 2003, 38, 1119–1127. [Google Scholar] [CrossRef]
- Sakiani, S.; Olsen, N.J.; Kovacs, W.J. Gonadal steroids and humoral immunity. Nat. Rev. Endocrinol. 2013, 9, 56–62. [Google Scholar] [CrossRef]
- Keestra, S.M.; Male, V.; Salali, G.D. Out of balance: The role of evolutionary mismatches in the sex disparity in autoimmune disease. Med. Hypotheses 2021, 151, 110558. [Google Scholar] [CrossRef]
- Aiello, A.; Farzaneh, F.; Candore, G.; Caruso, C.; Davinelli, S.; Gambino, C.M.; Ligotti, M.E.; Zareian, N.; Accardi, G. Immunosenescence and its hallmarks: How to oppose aging strategically? A review of potential options for therapeutic intervention. Front. Immunol. 2019, 10, 2247. [Google Scholar] [CrossRef]
- Hirokawa, K.; Utsuyama, M.; Hayashi, Y.; Kitagawa, M.; Makinodan, T.; Fulop, T. Slower immune system aging in women versus men in the Japanese population. Immun. Ageing 2013, 10, 19. [Google Scholar] [CrossRef]
- Dudkowska, M.; Janiszewska, D.; Karpa, A.; Broczek, K.; Dabrowski, M.; Sikora, E. The role of gender and labour status in immunosenescence of 65+ Polish population. Biogerontology 2017, 18, 581–590. [Google Scholar] [CrossRef]
- Piasecka, B.; Duffy, D.; Urrutia, A.; Quach, H.; Patin, E.; Posseme, C.; Bergstedt, J.; Charbit, B.; Rouilly, V.; MacPherson, C.R.; et al. Distinctive roles of age, sex, and genetics in shaping transcriptional variation of human immune responses to microbial challenges. Proc. Natl. Acad. Sci. USA 2018, 115, E488–E497. [Google Scholar] [CrossRef]
- Qu, K.; Zaba, L.C.; Giresi, P.G.; Li, R.; Longmire, M.; Kim, Y.H.; Greenleaf, W.J.; Chang, H.Y. Individuality and variation of personal regulomes in primary human T cells. Cell Syst. 2015, 1, 51–61. [Google Scholar] [CrossRef]
- Klein, S.L.; Morgan, R. The impact of sex and gender on immunotherapy outcomes. Biol. Sex Differ. 2020, 11, 24. [Google Scholar] [CrossRef]
- Brodin, P.; Jojic, V.; Gao, T.; Bhattacharya, S.; Angel, C.J.; Furman, D.; Shen-Orr, S.; Dekker, C.L.; Swan, G.E.; Butte, A.J.; et al. Variation in the human immune system is largely driven by non-heritable influences. Cell 2015, 160, 37–47. [Google Scholar] [CrossRef]
- Brodin, P. Systems-level patterns emerge. Nat. Rev. Immunol. 2019, 19, 87–88. [Google Scholar] [CrossRef]
- Olin, A.; Henckel, E.; Chen, Y.; Lakshmikanth, T.; Pou, C.; Mikes, J.; Gustafsson, A.; Bernhardsson, A.K.; Zhang, C.; Bohlin, K.; et al. Stereotypic immune system development in newborn children. Cell 2018, 174, 1277–1292.e14. [Google Scholar] [CrossRef]
- Alpert, A.; Pickman, Y.; Leipold, M.; Rosenberg-Hasson, Y.; Ji, X.; Gaujoux, R.; Rabani, H.; Starosvetsky, E.; Kveler, K.; Schaffert, S.; et al. A clinically meaningful metric of immune age derived from high-dimensional longitudinal monitoring. Nat. Med. 2019, 25, 487–495. [Google Scholar] [CrossRef]
- Yan, Z.; Maecker, H.T.; Brodin, P.; Nygaard, U.C.; Lyu, S.C.; Davis, M.M.; Nadeau, K.C.; Andorf, S. Aging and CMV discordance are associated with increased immune diversity between monozygotic twins. Immun. Ageing 2021, 18, 5. [Google Scholar] [CrossRef]
- Lakshmikanth, T.; Muhammad, S.A.; Olin, A.; Chen, Y.; Mikes, J.; Fagerberg, L.; Gummesson, A.; Bergström, G.; Uhlen, M.; Brodin, P. Human immune system variation during 1 Year. Cell Rep. 2020, 32, 107923. [Google Scholar] [CrossRef]
- Tran Van Hoi, E.; De Glas, N.A.; Portielje, J.E.A.; Van Heemst, D.; Van Den Bos, F.; Jochems, S.P.; Mooijaart, S.P. Biomarkers of the ageing immune system and their association with frailty—A systematic review. Exp. Gerontol. 2023, 176, 112163. [Google Scholar] [CrossRef]
- Alberro, A.; Iribarren-Lopez, A.; Sáenz-Cuesta, M.; Matheu, A.; Vergara, I.; Otaegui, D. Inflammaging markers characteristic of advanced age show similar levels with frailty and dependency. Sci. Rep. 2021, 11, 4358. [Google Scholar] [CrossRef]
- Xu, W.; Wong, G.; Hwang, Y.Y.; Larbi, A. The untwining of immunosenescence and aging. Semin. Immunopathol. 2020, 42, 559–572. [Google Scholar] [CrossRef]
- Medoro, A.; Saso, L.; Scapagnini, G.; Davinelli, S. NRF2 signaling pathway and telomere length in aging and age-related diseases. Mol. Cell. Biochem. 2023. [Google Scholar] [CrossRef]
- Vaiserman, A.; Krasnienkov, D. Telomere length as a marker of biological age: State-of-the-art, open issues, and future perspectives. Front. Genet. 2021, 11, 630186. [Google Scholar] [CrossRef]
- Zhang, J.; He, T.; Xue, L.; Guo, H. Senescent T cells: A potential biomarker and target for cancer therapy. EBioMedicine 2021, 68, 103409. [Google Scholar] [CrossRef]
- Verma, K.; Ogonek, J.; Varanasi, P.R.; Luther, S.; Bünting, I.; Thomay, K.; Behrens, Y.L.; Mischak-Weissinger, E.; Hambach, L. Human CD8+ CD57- TEMRA cells: Too young to be called “old”. PLoS ONE 2017, 12, e0177405. [Google Scholar] [CrossRef]
- Miron, M.; Meng, W.; Rosenfeld, A.M.; Dvorkin, S.; Poon, M.M.L.; Lam, N.; Kumar, B.V.; Louzoun, Y.; Luning Prak, E.T.; Farber, D.L. Maintenance of the human memory T cell repertoire by subset and tissue site. Genome Med. 2021, 13, 100. [Google Scholar] [CrossRef]
- Guo, L.; Liu, X.; Su, X. The role of TEMRA cell-mediated immune senescence in the development and treatment of HIV disease. Front. Immunol. 2023, 14, 1284293. [Google Scholar] [CrossRef]
- Yeh, Y.W.; Xiang, Z. Mouse hygiene status-A tale of two environments for mast cells and allergy. Allergol. Int. 2023, 73, 58–64. [Google Scholar] [CrossRef]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef]
- Forlin, R.; James, A.; Brodin, P. Making human immune systems more interpretable through systems immunology. Trends Immunol. 2023, 44, 577–584. [Google Scholar] [CrossRef]
- Narang, V.; Decraene, J.; Wong, S.Y.; Aiswarya, B.S.; Wasem, A.R.; Leong, S.R.; Gouaillard, A. Systems immunology: A survey of modeling formalisms, applications and simulation tools. Immunol. Res. 2012, 53, 251–265. [Google Scholar] [CrossRef]
- Pulendran, B.; Davis, M.M. The science and medicine of human immunology. Science 2020, 369, eaay4014. [Google Scholar] [CrossRef]
- Davis, M.M.; Tato, C.M.; Furman, D. Systems immunology: Just getting started. Nat. Immunol. 2017, 18, 725–732. [Google Scholar] [CrossRef]
- Ginhoux, F.; Yalin, A.; Dutertre, C.A.; Amit, I. Single-cell immunology: Past, present, and future. Immunity 2022, 55, 393–404. [Google Scholar] [CrossRef]
- Poon, M.M.L.; Farber, D.L. The whole body as the system in systems immunology. iScience 2020, 23, 101509. [Google Scholar] [CrossRef]
- Tebani, A.; Gummesson, A.; Zhong, W.; Koistinen, I.S.; Lakshmikanth, T.; Olsson, L.M.; Boulund, F.; Neiman, M.; Stenlund, H.; Hellström, C.; et al. Integration of molecular profiles in a longitudinal wellness profiling cohort. Nat. Commun. 2020, 11, 4487. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. QuickStats: Death Rates* from Influenza and Pneumonia† Among Persons Aged ≥65 Years, by Sex and Age Group—National Vital Statistics System, United States. 2018. Available online: https://www.cdc.gov/mmwr/volumes/69/wr/mm6940a5.htm#:~:text=In%202018%2C%20the%20death%20rate,those%20aged%20%E2%89%A585%20years (accessed on 18 March 2024).
- Montgomery, R.R. Age-related alterations in immune responses to West Nile virus infection. Clin. Exp. Immunol. 2017, 187, 26–34. [Google Scholar] [CrossRef]
- Bartleson, J.M.; Radenkovic, D.; Covarrubias, A.J.; Furman, D.; Winer, D.A.; Verdin, E. SARS-CoV-2, COVID-19 and the ageing immune system. Nat. Aging 2021, 1, 769–782. [Google Scholar] [CrossRef]
- Cho, S.J.; Stout-Delgado, H.W. Aging and lung disease. Annu. Rev. Physiol. 2020, 82, 433–459. [Google Scholar] [CrossRef]
- Gomez, C.R.; Boehmer, E.D.; Kovacs, E.J. The aging innate immune system. Curr. Opin. Immunol. 2005, 17, 457–462. [Google Scholar] [CrossRef]
- Shinjyo, N.; Kita, K. Infection and immunometabolism in the central nervous system: A possible mechanistic link between metabolic imbalance and dementia. Front. Cell. Neurosci. 2021, 15, 765217. [Google Scholar] [CrossRef]
- Al-Jabri, M.; Rosero, C.; Saade, E.A. Vaccine-preventable diseases in older adults. Infect. Dis. Clin. N. Am. 2023, 37, 103–121. [Google Scholar] [CrossRef]
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T cell exhaustion during persistent viral infections. Virology 2015, 479–480, 180–193. [Google Scholar] [CrossRef]
- Marrella, V.; Facoetti, A.; Cassani, B. Cellular Senescence in immunity against infections. Int. J. Mol. Sci. 2022, 23, 11845. [Google Scholar] [CrossRef]
- Wang, A.S.; Steers, N.J.; Parab, A.R.; Gachon, F.; Sweet, M.J.; Mysorekar, I.U. Timing is everything: Impact of development, ageing and circadian rhythm on macrophage functions in urinary tract infections. Mucosal Immunol. 2022, 15, 1114–1126. [Google Scholar] [CrossRef]
- Reyes, A.; Ortiz, G.; Duarte, L.F.; Fernández, C.; Hernández-Armengol, R.; Palacios, P.A.; Prado, Y.; Andrade, C.A.; Rodriguez-Guilarte, L.; Kalergis, A.M.; et al. Contribution of viral and bacterial infections to senescence and immunosenescence. Front. Cell. Infect. Microbiol. 2023, 13, 1229098. [Google Scholar] [CrossRef]
- Batista, M.A.; Calvo-Fortes, F.; Silveira-Nunes, G.; Camatta, G.C.; Speziali, E.; Turroni, S.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Neretti, N.; Maioli, T.U.; et al. Inflammaging in endemic areas for infectious diseases. Front. Immunol. 2020, 11, 579972. [Google Scholar] [CrossRef]
- Saavedra, D.; Añé-Kourí, A.L.; Barzilai, N.; Caruso, C.; Cho, K.H.; Fontana, L.; Franceschi, C.; Frasca, D.; Ledón, N.; Niedernhofer, L.J.; et al. Aging and chronic inflammation: Highlights from a multidisciplinary workshop. Immun. Ageing 2023, 20, 25. [Google Scholar] [CrossRef]
- Strulik, H.; Grossmann, V. The economics of aging with infectious and chronic diseases. Econ. Hum. Biol. 2023, 52, 101319. [Google Scholar] [CrossRef]
- Rothan-Tondeur, M.; Gavazzi, G.; Piette, F.; Lejeune, B.; de Wazières, B. L’observatoire du risque infectieux en gériatrie. Neurol. Psychiatr. Gériatr. 2005, 5, 20–25. [Google Scholar] [CrossRef]
- Yoshikawa, T.T. Epidemiology and unique aspects of aging and infectious diseases. Clin. Infect. Dis. 2000, 30, 931–933. [Google Scholar] [CrossRef] [PubMed]
- Du, W.Y.; Yin, C.N.; Wang, H.T.; Li, Z.W.; Wang, W.J.; Xue, F.Z.; Zhao, L.; Cao, W.C.; Cheeloo EcoHealth Consortium (CLEC). Infectious diseases among elderly persons: Results from a population-based observational study in Shandong province, China, 2013–2017. J. Glob. Health. 2021, 11, 08010. [Google Scholar] [CrossRef] [PubMed]
- Bender, B.S.H.I.V. Aging as a model for immunosenescence. J. Gerontol. A. Biol. Sci. Med. Sci. 1997, 52, M261–M263. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G. HIV infection, inflammation, immunosenescence, and aging. Annu. Rev. Med. 2011, 62, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Appay, V.; Rowland-Jones, S.L. Premature ageing of the immune system: The cause of AIDS? Trends Immunol. 2002, 23, 580–855. [Google Scholar] [CrossRef]
- Grifoni, A.; Alonzi, T.; Alter, G.; Noonan, D.M.; Landay, A.L.; Albini, A.; Goletti, D. Impact of aging on immunity in the context of COVID-19, HIV, and tuberculosis. Front. Immunol. 2023, 14, 1146704. [Google Scholar] [CrossRef] [PubMed]
- Hatzakis, A.; Touloumi, G.; Karanicolas, R.; Karafoulidou, A.; Mandalaki, T.; Anastassopoulou, C.; Zhang, L.; Goedert, J.J.; Ho, D.D.; Kostrikis, L.G. Effect of recent thymic emigrants on progression of HIV-1 disease. Lancet 2000, 355, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lewin, S.R.; Markowitz, M.; Lin, H.H.; Skulsky, E.; Karanicolas, R.; He, Y.; Jin, X.; Tuttleton, S.; Vesanen, M.; et al. Measuring recent thymic emigrants in blood of normal and HIV-1-infected individuals before and after effective therapy. J. Exp. Med. 1999, 190, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Quiros-Roldan, E.; Serana, F.; Chiarini, M.; Zanotti, C.; Sottini, A.; Gotti, D.; Torti, C.; Caimi, L.; Imberti, L. Effects of combined antiretroviral therapy on B- and T-cell release from production sites in long-term treated HIV-1+ patients. J. Transl. Med. 2012, 10, 94. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elias Junior, E.; Gubert, V.T.; Bonin-Jacob, C.M.; Puga, M.A.M.; Gouveia, C.G.; Sichinel, A.H.; Tozetti, I.A. CD57 T cells associated with immunosenescence in adults living with HIV or AIDS. Immunology 2024, 171, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Heather, J.M.; Best, K.; Oakes, T.; Gray, E.R.; Roe, J.K.; Thomas, N.; Friedman, N.; Noursadeghi, M.; Chain, B. Dynamic perturbations of the T-cell receptor repertoire in chronic HIV infection and following antiretroviral therapy. Front. Immunol. 2016, 6, 644. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.T.; Brown, J.; Shaw, E.; Uddin, I.; Tsaliki, E.; Roe, J.K.; Pollara, G.; Sun, Y.; Heather, J.M.; Lipman, M.; et al. Persistent T cell repertoire perturbation and T cell activation in HIV after long term treatment. Front. Immunol. 2021, 12, 634489. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.S.; Espíndola, M.S.; Zambuzi, F.A.; Galvão-Lima, L.J.; Cacemiro, M.C.; Soares, M.R.; Santana, B.A.; Calado, R.T.; Bollela, V.R.; Frantz, F.G. Immunosenescence in chronic HIV infected patients impairs essential functions of their natural killer cells. Int. Immunopharmacol. 2020, 84, 106568. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Verdin, E.; McCune, J.M. Immunosenescence and HIV. Curr. Opin. Immunol. 2012, 24, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Tizazu, A.M.; Mengist, H.M.; Demeke, G. Aging, inflammaging and immunosenescence as risk factors of severe COVID-19. Immun. Ageing 2022, 19, 53. [Google Scholar] [CrossRef]
- Asghari, F.; Asghary, A.; Majidi Zolbanin, N.; Faraji, F.; Jafari, R. Immunosenescence and Inflammaging in COVID-19. Viral. Immunol. 2023, 36, 579–592. [Google Scholar] [CrossRef] [PubMed]
- OECD Data. Influenza Vaccination Rates. Available online: https://data.oecd.org/healthcare/influenza-vaccination-rates.htm. (accessed on 18 March 2024).
- Patel, M.M.; York, I.A.; Monto, A.S.; Thompson, M.G.; Fry, A.M. Immune-mediated attenuation of influenza illness after infection: Opportunities and challenges. Lancet Microbe 2021, 2, e715–e725. [Google Scholar] [CrossRef]
- Plowden, J.; Renshaw-Hoelscher, M.; Engleman, C.; Katz, J.; Sambhara, S. Innate immunity in aging: Impact on macrophage function. Aging Cell 2004, 3, 161–167. [Google Scholar] [CrossRef]
- Albright, J.M.; Dunn, R.C.; Shults, J.A.; Boe, D.M.; Afshar, M.; Kovacs, E.J. Advanced age alters monocyte and macrophage responses. Antioxid. Redox. Signal. 2016, 25, 805–815. [Google Scholar] [CrossRef]
- Hearps, A.C.; Martin, G.E.; Angelovich, T.A.; Cheng, W.J.; Maisa, A.; Landay, A.L.; Jaworowski, A.; Crowe, S.M. Aging is associated with chronic innate immune activation and dysregulation of monocyte phenotype and function. Aging Cell 2012, 11, 867–875. [Google Scholar] [CrossRef]
- McElhaney, J.E.; Verschoor, C.P.; Andrew, M.K.; Haynes, L.; Kuchel, G.A.; Pawelec, G. The immune response to influenza in older humans: Beyond immune senescence. Immun. Ageing. 2020, 17, 10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Fisher, E.M.; Murasko, D.M. CD8 T cell responses to influenza virus infection in aged mice. Ageing Res. Rev. 2011, 10, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Parks, O.B.; Eddens, T.; Sojati, J.; Lan, J.; Zhang, Y.; Oury, T.D.; Ramsey, M.; Erickson, J.J.; Byersdorfer, C.A.; Williams, J.V. Terminally exhausted CD8+ T cells contribute to age-dependent severity of respiratory virus infection. Immun. Ageing 2023, 20, 40. [Google Scholar] [CrossRef] [PubMed]
- van de Sandt, C.E.; Nguyen, T.H.O.; Gherardin, N.A.; Crawford, J.C.; Samir, J.; Minervina, A.A.; Pogorelyy, M.V.; Rizzetto, S.; Szeto, C.; Kaur, J.; et al. Newborn and child-like molecular signatures in older adults stem from TCR shifts across human lifespan. Nat. Immunol. 2023, 24, 1890–1907. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in elderly adults. Drugs Aging 2005, 22, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Mlinaric-Galinovic, G.; Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in the elderly. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 777–781. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Savic, M.; Penders, Y.; Shi, T.; Branche, A.; Pirçon, J.Y. Respiratory syncytial virus disease burden in adults aged 60 years and older in high-income countries: A systematic literature review and meta-analysis. Influenza Other Respir. Viruses 2023, 17, e13031. [Google Scholar] [CrossRef] [PubMed]
- Looney, R.J.; Falsey, A.R.; Walsh, E.; Campbell, D. Effect of aging on cytokine production in response to respiratory syncytial virus infection. J. Infect. Dis. 2002, 185, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Salaun, B.; De Smedt, J.; Vernhes, C.; Moureau, A.; Öner, D.; Bastian, A.R.; Janssens, M.; Balla-Jhagjhoorsingh, S.; Aerssens, J.; Lambert, C.; et al. T cells, more than antibodies, may prevent symptoms developing from respiratory syncytial virus infections in older adults. Front. Immunol. 2023, 14, 1260146. [Google Scholar] [CrossRef] [PubMed]
- Njue, A.; Nuabor, W.; Lyall, M.; Margulis, A.; Mauskopf, J.; Curcio, D.; Kurosky, S.; Gessner, B.D.; Begier, E. Systematic literature review of risk factors for poor outcomes among adults with respiratory syncytial virus infection in high-income countries. Open Forum Infect. Dis. 2023, 10, ofad513. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Wilkinson, T.M.A. Respiratory viral infections in the elderly. Ther. Adv. Respir. Dis. 2021, 15, 1753466621995050. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Pedrero, M.Á.; Osterhaus, A.D.M.E.; Becker, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Aging and options to halt declining immunity to virus infections. Front. Immunol. 2021, 12, 681449. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.; Pera, A.; Sanchez-Correa, B.; Alonso, C.; Lopez-Fernandez, I.; Morgado, S.; Tarazona, R.; Solana, R. Effect of age and CMV on NK cell subpopulations. Exp. Gerontol. 2014, 54, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Pita-Lopez, M.L.; Gayoso, I.; DelaRosa, O.; Casado, J.G.; Alonso, C.; Muñoz-Gomariz, E.; Tarazona, R.; Solana, R. Effect of ageing on CMV-specific CD8 T cells from CMV seropositive healthy donors. Immun. Ageing 2009, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, A.M.; Bennett, M.S.; Park, B.; Uhrlaub, J.L.; Martinez, C.; Pulko, V.; Currier, N.L.; Nikolich-Žugich, D.; Kaye, J.; Nikolich-Žugich, J. Aging and cytomegalovirus infection differentially and jointly affect distinct circulating T cell subsets in humans. J. Immunol. 2014, 192, 2143–2155. [Google Scholar] [CrossRef]
- Khan, N.; Shariff, N.; Cobbold, M.; Bruton, R.; Ainsworth, J.A.; Sinclair, A.J.; Nayak, L.; Moss, P.A. Cytomegalovirus seropositivity drives the CD8 T cell repertoire toward greater clonality in healthy elderly individuals. J. Immunol. 2002, 169, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Attaf, M.; Malik, A.; Severinsen, M.C.; Roider, J.; Ogongo, P.; Buus, S.; Ndung’u, T.; Leslie, A.; Kløverpris, H.N.; Matthews, P.C.; et al. Major TCR repertoire perturbation by immunodominant HLA-B*44:03-restricted CMV-specific T cells. Front. Immunol. 2018, 9, 2539. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, F.; Clave, E.; Wanke, K.; von Braun, A.; Bondet, V.; Alanio, C.; Douay, C.; Baque, M.; Lependu, C.; Marconi, P.; et al. Primary immune responses are negatively impacted by persistent herpesvirus infections in older people: Results from an observational study on healthy subjects and a vaccination trial on subjects aged more than 70 years old. EBioMedicine 2022, 76, 103852. [Google Scholar] [CrossRef]
- Müller-Durovic, B.; Grählert, J.; Devine, O.P.; Akbar, A.N.; Hess, C. CD56-negative NK cells with impaired effector function expand in CMV and EBV co-infected healthy donors with age. Aging 2019, 11, 724–740. [Google Scholar] [CrossRef] [PubMed]
- Lanfermeijer, J.; de Greef, P.C.; Hendriks, M.; Vos, M.; van Beek, J.; Borghans, J.A.M.; van Baarle, D. Age and CMV-infection jointly affect the EBV-specific CD8+ T-cell repertoire. Front. Aging 2021, 2, 665637. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, S.; Carlisi, M.; Santoro, M.; Napolitano, M.; Raso, S.; Siragusa, S. Immunosenescence and lymphomagenesis. Immun. Ageing 2018, 15, 22. [Google Scholar] [CrossRef] [PubMed]
- Martín-Escolano, R.; Vidal-Alcántara, E.J.; Crespo, J.; Ryan, P.; Real, L.M.; Lazo-Álvarez, J.I.; Cabezas-González, J.; Macías, J.; Arias-Loste, M.T.; Cuevas, G.; et al. Immunological and senescence biomarker profiles in patients after spontaneous clearance of hepatitis C virus: Gender implications for long-term health risk. Immun. Ageing 2023, 20, 62. [Google Scholar] [CrossRef] [PubMed]
- Voehringer, D.; Blaser, C.; Brawand, P.; Raulet, D.H.; Hanke, T.; Pircher, H. Viral infections induce abundant numbers of senescent CD8 T cells. J. Immunol. 2001, 167, 4838–4843. [Google Scholar] [CrossRef] [PubMed]
- Giannakoulis, V.G.; Dubovan, P.; Papoutsi, E.; Kataki, A.; Koskinas, J. Senescence in HBV-, HCV- and NAFLD- mediated hepatocellular carcinoma and senotherapeutics: Current evidence and future perspective. Cancers 2021, 13, 4732. [Google Scholar] [CrossRef]
- Kang, T.W.; Yevsa, T.; Woller, N.; Hoenicke, L.; Wuestefeld, T.; Dauch, D.; Hohmeyer, A.; Gereke, M.; Rudalska, R.; Potapova, A.; et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature 2011, 479, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.M.; McSharry, B.P.; Steain, M.; Ashhurst, T.M.; Slobedman, B.; Abendroth, A. Varicella zoster virus productively infects human natural killer cells and manipulates phenotype. PLoS Pathog. 2018, 14, e1006999. [Google Scholar] [CrossRef] [PubMed]
- Gerada, C.; Campbell, T.M.; Kennedy, J.J.; McSharry, B.P.; Steain, M.; Slobedman, B.; Abendroth, A. Manipulation of the innate immune response by varicella zoster virus. Front. Immunol. 2020, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J. Immune senescence and vaccines to prevent herpes zoster in older persons. Curr. Opin. Immunol. 2012, 24, 494–500. [Google Scholar] [CrossRef]
- Johnson, R.W. Herpes Zoster and Postherpetic Neuralgia: A review of the effects of vaccination. Aging Clin. Exp. Res. 2009, 21, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Curran, D.; Doherty, T.M.; Lecrenier, N.; Breuer, T. Healthy ageing: Herpes zoster infection and the role of zoster vaccination. NPJ Vaccines 2023, 8, 184. [Google Scholar] [CrossRef]
- Seoane, R.; Vidal, S.; Bouzaher, Y.H.; El Motiam, A.; Rivas, C. The interaction of viruses with the cellular senescence response. Biology 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Arvia, R.; Zakrzewska, K.; Giovannelli, L.; Ristori, S.; Frediani, E.; Del Rosso, M.; Mocali, A.; Stincarelli, M.A.; Laurenzana, A.; Fibbi, G.; et al. Parvovirus B19 induces cellular senescence in human dermal fibroblasts: Putative role in systemic sclerosis-associated fibrosis. Rheumatology 2022, 61, 3864–3874. [Google Scholar] [CrossRef] [PubMed]
- AbuBakar, S.; Shu, M.H.; Johari, J.; Wong, P.F. Senescence affects endothelial cells susceptibility to dengue virus infection. Int. J. Med. Sci. 2014, 11, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Siebels, S.; Czech-Sioli, M.; Spohn, M.; Schmidt, C.; Theiss, J.; Indenbirken, D.; Günther, T.; Grundhoff, A.; Fischer, N. Merkel cell polyomavirus DNA replication induces senescence in human dermal fibroblasts in a Kap1/Trim28-dependent manner. mBio 2020, 11, e00142-20. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. West Nile Virus. Available online: https://www.cdc.gov/westnile/statsmaps/historic-data.html?CDC_AA_refVal=https%3A%2F%2Fwww.cdc.gov%2Fwestnile%2Fstatsmaps%2FcumMapsData.html (accessed on 18 March 2024).
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Qian, F.; Wang, X.; Zhang, L.; Lin, A.; Zhao, H.; Fikrig, E.; Montgomery, R. Impaired interferon signaling in dendritic cells from older donors infected in vitro with West Nile virus. J. Infect. Dis. 2011, 203, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Strauss-Albee, D.M.; Zhou, J.Q.; Malawista, A.; Garcia, M.N.; Murray, K.O.; Blish, C.A.; Montgomery, R.R. The natural killer cell response to West Nile virus in young and old individuals with or without a prior history of infection. PLoS ONE 2017, 12, e0172625. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.A.; Mao-Draayer, Y. Aging and lymphocyte changes by immunomodulatory therapies impact PML risk in multiple sclerosis patients. Mult. Scler. 2018, 24, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, L.; Scarpazza, C.; Imberti, L.; Cordioli, C.; De Rossi, N.; Capra, R. Age as a risk factor for early onset of natalizumab-related progressive multifocal leukoencephalopathy. J. Neurovirol. 2017, 23, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, D.; Sottini, A.; Capra, R.; Scarpazza, C.; Bresciani, R.; Notarangelo, L.D.; Imberti, L. Lack of specific T- and B-cell clonal expansions in multiple sclerosis patients with progressive multifocal leukoencephalopathy. Sci. Rep. 2019, 9, 16605. [Google Scholar] [CrossRef] [PubMed]
- Behr, M.A.; Edelstein, P.H.; Ramakrishnan, L. Revisiting the timetable of tuberculosis. BMJ 2018, 362, k2738. [Google Scholar] [CrossRef] [PubMed]
- Teo, A.K.J.; Morishita, F.; Islam, T.; Viney, K.; Ong, C.W.M.; Kato, S.; Kim, H.; Liu, Y.; Oh, K.H.; Yoshiyama, T.; et al. Tuberculosis in older adults: Challenges and best practices in the Western Pacific Region. Lancet Reg. Health West. Pac. 2023, 36, 100770. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, S. Tuberculosis and aging: A global health problem. Clin. Infect. Dis. 2001, 33, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, H.; Ikushima, S.; Kondo, K.; Satake, K.; Inomata, M.; Moriya, A.; Ando, T. Diagnostic performance of interferon-gamma release assays in elderly populations in comparison with younger populations. J. Infect. Chemother. 2013, 19, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Kim, Y.H.; Jeon, K.; Jeong, B.H.; Ryu, Y.J.; Choi, J.C.; Kim, H.C.; Koh, W.J. Factors that predict negative results of QuantiFERON-TB Gold In-Tube Test in patients with culture-confirmed tuberculosis: A Multicenter Retrospective Cohort Study. PLoS ONE 2015, 10, e0129792. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, K.; Kubo, T.; Akagi, K.; Miyashita, R.; Kondo, A.; Ehara, N.; Takazono, T.; Sakamoto, N.; Mukae, H. Clinical evaluation of QuantiFERON®-TB Gold Plus directly compared with QuantiFERON®-TB Gold In-Tube and T-Spot®. TB for active pulmonary tuberculosis in the elderly. J. Infect. Chemother. 2021, 27, 1716–1722. [Google Scholar] [CrossRef] [PubMed]
- Byng-Maddick, R.; Noursadeghi, M. Does tuberculosis threaten our ageing populations? BMC Infect. Dis. 2016, 16, 119. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Rossi, R.; Nshimyumukiza, L.; Wusiman, A.; Zdraveska, N.; Eldin, M.S. Convergence of a diabetes mellitus, protein energy malnutrition, and TB epidemic: The neglected elderly population. BMC Infect. Dis. 2016, 16, 361. [Google Scholar] [CrossRef]
- Rajagopalan, S. Tuberculosis in Older Adults. Clin. Geriatr. Med. 2016, 32, 479–491. [Google Scholar] [CrossRef]
- Asokan, S. Immune issues in elderly with TB. Indian J. Tuberc. 2022, 69 (Suppl. S2), S241–S245. [Google Scholar] [CrossRef] [PubMed]
- Canan, C.H.; Gokhale, N.S.; Carruthers, B.; Lafuse, W.P.; Schlesinger, L.S.; Torrelles, J.B.; Turner, J. Characterization of lung inflammation and its impact on macrophage function in aging. J. Leukoc. Biol. 2014, 96, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Ault, R.; Dwivedi, V.; Koivisto, E.; Nagy, J.; Miller, K.; Nagendran, K.; Chalana, I.; Pan, X.; Wang, S.H.; Turner, J. Altered monocyte phenotypes but not impaired peripheral T cell immunity may explain susceptibility of the elderly to develop tuberculosis. Exp. Gerontol. 2018, 111, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Namdeo, M.; Kandel, R.; Thakur, P.K.; Mohan, A.; Dey, A.B.; Mitra, D.K. Old age-associated enrichment of peripheral T regulatory cells and altered redox status in pulmonary tuberculosis patients. Eur. J. Immunol. 2020, 50, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Hase, I.; Toren, K.G.; Hirano, H.; Sakurai, K.; Horne, D.J.; Saito, T.; Narita, M. Pulmonary tuberculosis in older adults: Increased mortality related to tuberculosis within two months of treatment initiation. Drugs Aging 2021, 38, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.Y.; Kim, M.J.; Cheong, H.K.; Oh, I.H. Estimating disability-adjusted life years due to tuberculosis in Korea through to the year 2040. Int. J. Environ. Res. Public Health 2020, 17, 5960. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Matsumoto, H.; Horita, N.; Hara, Y.; Kobayashi, N.; Kaneko, T. Prognostic factors for mortality in invasive pneumococcal disease in adult: A system review and meta-analysis. Sci. Rep. 2021, 11, 11865. [Google Scholar] [CrossRef] [PubMed]
- Simell, B.; Vuorela, A.; Ekström, N.; Palmu, A.; Reunanen, A.; Meri, S.; Käyhty, H.; Väkeväinen, M. Aging reduces the functionality of anti-pneumococcal antibodies and the killing of Streptococcus pneumoniae by neutrophil phagocytosis. Vaccine 2011, 29, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- Shivshankar, P.; Boyd, A.R.; Le Saux, C.J.; Yeh, I.T.; Orihuela, C.J. Cellular senescence increases expression of bacterial ligands in the lungs and is positively correlated with increased susceptibility to pneumococcal pneumonia. Aging Cell 2011, 10, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Weight, C.M.; Jochems, S.P.; Adler, H.; Ferreira, D.M.; Brown, J.S.; Heyderman, R.S. Insights into the effects of mucosal epithelial and innate immune dysfunction in older people on host interactions with Streptococcus pneumoniae. Front. Cell. Infect. Microbiol. 2021, 11, 651474. [Google Scholar] [CrossRef] [PubMed]
- Dunn-Walters, D.K.; O’Hare, J.S. Older Human B Cells and Antibodies. In Handbook of Immunosenescence: Basic Understanding and Clinical Implications; Fulop, T., Franceschi, C., Hirokawa, K., Pawelec, G., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 785–819. [Google Scholar] [CrossRef]
- Ligon, M.M.; Joshi, C.S.; Fashemi, B.E.; Salazar, A.M.; Mysorekar, I.U. Effects of aging on urinary tract epithelial homeostasis and immunity. Dev. Biol. 2023, 493, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Thomas-White, K.J.; Kliethermes, S.; Rickey, L.; Lukacz, E.S.; Richter, H.E.; Moalli, P.; Zimmern, P.; Norton, P.; Kusek, J.W.; Wolfe, A.J.; et al. Evaluation of the urinary microbiota of women with uncomplicated stress urinary incontinence. Am. J. Obstet. Gynecol. 2017, 216, 55.e1–55.e16. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.A.; Uhlemann, A.C.; Barasch, J. Innate bacteriostatic mechanisms defend the urinary tract. Annu. Rev. Physiol. 2022, 84, 533–558. [Google Scholar] [CrossRef] [PubMed]
- Mora-Bau, G.; Platt, A.M.; van Rooijen, N.; Randolph, G.J.; Albert, M.L.; Ingersoll, M.A. Macrophages subvert adaptive immunity to urinary tract infection. PLoS Pathog. 2015, 11, e1005044. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Y.F.; Wang, Y.S.; Yang, Q.; Xiao, Y.L.; Cai, H.R.; Xie, C.M. Using ROS as a second messenger, NADPH oxidase 2 mediates macrophage senescence via interaction with NF-κB during Pseudomonas aeruginosa infection. Oxid. Med. Cell. Longev. 2018, 2018, 9741838. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, M.; Kumamoto, Y.; Hirose, T.; Matsuura, A. Tissue gamma/delta T cells in experimental urinary tract infection relationship between other immuno-competent cells. Kansenshogaku Zasshi 1994, 68, 1498–1511. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sivick, K.E.; Schaller, M.A.; Smith, S.N.; Mobley, H.L. The innate immune response to uropathogenic Escherichia coli involves IL-17A in a murine model of urinary tract infection. J. Immunol. 2010, 184, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Szaraz-Szeles, M.; Mezei, Z.; Barath, S.; Hevessy, Z. Age-dependent frequency of unconventional T cells in a healthy adult Caucasian population: A combinational study of invariant natural killer T cells, γδ T cells, and mucosa-associated invariant T cells. Geroscience 2022, 44, 2047–2060. [Google Scholar] [CrossRef] [PubMed]
- Schmucker, D.L.; Heyworth, M.F.; Owen, R.L.; Daniels, C.K. Impact of aging on gastrointestinal mucosal immunity. Dig. Dis. Sci. 1996, 41, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Madhogaria, B.; Bhowmik, P.; Kundu, A. Correlation between human gut microbiome and diseases. Infect. Med. 2022, 1, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, H.; Zeng, G.; Pang, B.; Wang, Q.; Wei, J. Gut microbiome-mediated mechanisms in aging-related diseases: Are probiotics ready for prime time? Front. Pharmacol. 2023, 14, 1178596. [Google Scholar] [CrossRef] [PubMed]
- Dumic, I.; Nordin, T.; Jecmenica, M.; Stojkovic Lalosevic, M.; Milosavljevic, T.; Milovanovic, T. Gastrointestinal Tract Disorders in Older Age. Can. J. Gastroenterol. Hepatol. 2019, 2019, 6757524. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P.; LaMont, J.T. Clostridium difficile--more difficult than ever. N. Engl. J. Med. 2008, 359, 1932–1940. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.T.; Norman, D.C. Geriatric infectious diseases: Current concepts on diagnosis and management. J. Am. Geriatr. Soc. 2017, 65, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Kokkola, A.; Sipponen, P.; Rautelin, H.; Härkönen, M.; Kosunen, T.U.; Haapiainen, R.; Puolakkainen, P. The effect of Helicobacter pylori eradication on the natural course of atrophic gastritis with dysplasia. Aliment. Pharmacol. Ther. 2002, 16, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.M.; Kim, N.; Park, J.H.; Lee, D.H. Changes in gastric corpus microbiota with age and after Helicobacter pylori eradication: A long-term follow-up study. Front. Microbiol. 2021, 11, 621879. [Google Scholar] [CrossRef] [PubMed]
- Dukowicz, A.C.; Lacy, B.E.; Levine, G.M. Small intestinal bacterial overgrowth: A comprehensive review. Gastroenterol. Hepatol. 2007, 3, 112–122. [Google Scholar]
- Parlesak, A.; Klein, B.; Schecher, K.; Bode, J.C.; Bode, C. Prevalence of small bowel bacterial overgrowth and its association with nutrition intake in nonhospitalized older adults. J. Am. Geriatr. Soc. 2003, 51, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, R.; Elashiry, M.; Liu, Y.; El-Awady, A.; Hamrick, M.; Cutler, C.W. Porphyromonas gingivalis provokes exosome secretion and paracrine immune senescence in bystander dendritic cells. Front. Cell. Infect. Microbiol. 2021, 11, 669989. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Rowe, T.A.; McKoy, J.M. Sepsis in older adults. Infect. Dis. Clin. N. Am. 2017, 31, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, K.S.; Hsann, Y.M.; Lim, V.; Ong, B.C. The effect of comorbidity and age on hospital mortality and length of stay in patients with sepsis. J. Crit. Care 2010, 25, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J. Immune-mediated inflammatory diseases as long-term sepsis complications: Long-term persistence of host dysregulation? J. Intern. Med. 2024, 295, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Alebachew, G.; Teka, B.; Endris, M.; Shiferaw, Y.; Tessema, B. Etiologic agents of bacterial sepsis and their antibiotic susceptibility patterns among patients living with Human Immunodeficiency Virus at Gondar University Teaching Hospital, Northwest Ethiopia. Biomed. Res. Int. 2016, 2016, 5371875. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Huang, S.Y.; Sun, J.H.; Zhang, H.C.; Cai, Q.L.; Gao, C.; Li, L.; Cao, J.; Xu, F.; Zhou, Y.; et al. Sepsis-induced immunosuppression: Mechanisms, diagnosis and current treatment options. Mil. Med. Res. 2022, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Lu, Y.Q. Editorial: Immunosenescence after sepsis. Front. Immunol. 2023, 14, 1177148. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, R.; Maiwall, R.; Rajan, V.; Islam, M.; Baweja, S.; Kaur, N.; Kumar, G.; Ramakrishna, G.; Sarin, S.K.; Trehanpati, N. Granulocyte-Macrophage Colony-Stimulating Factor modulates myeloid-derived suppressor cells and Treg activity in decompensated cirrhotic patients with sepsis. Front. Immunol. 2022, 313, 828949. [Google Scholar] [CrossRef]
- Malavika, M.; Sanju, S.; Poorna, M.R.; Vishnu Priya, V.; Sidharthan, N.; Varma, P.; Mony, U. Role of myeloid derived suppressor cells in sepsis. Int. Immunopharmacol. 2022, 104, 108452. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Monneret, G. Advances in the understanding and treatment of sepsis-induced immunosuppression. Nat. Rev. Nephrol. 2018, 14, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Cuenca, A.G.; Delano, M.J.; Kelly-Scumpia, K.M.; Moreno, C.; Scumpia, P.O.; Laface, D.M.; Heyworth, P.G.; Efron, P.A.; Moldawer, L.L. A paradoxical role for myeloid-derived suppressor cells in sepsis and trauma. Mol. Med. 2011, 17, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Marik, P.E.; Zaloga, G.P. NORASEPT II Study Investigators. North American Sepsis Trial II. The effect of aging on circulating levels of proinflammatory cytokines during septic shock. Norasept II Study Investigators. J. Am. Geriatr. Soc. 2001, 49, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro da Silva, F.; Zampieri, F.G.; Barbeiro, D.F.; Barbeiro, H.V.; Goulart, A.C.; Torggler Filho, F.; Velasco, I.T.; da Cruz Neto, L.M.; de Souza, H.P.; Machado, M.C. Septic shock in older people: A prospective cohort study. Immun. Ageing 2013, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Ginde, A.A.; Blatchford, P.J.; Trzeciak, S.; Hollander, J.E.; Birkhahn, R.; Otero, R.; Osborn, T.M.; Moretti, E.; Nguyen, H.B.; Gunnerson, K.J.; et al. Age-related differences in biomarkers of acute inflammation during hospitalization for sepsis. Shock 2014, 42, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Babayan, S.A.; Sinclair, A.; Duprez, J.S.; Selman, C. Chronic helminth infection burden differentially affects haematopoietic cell development while ageing selectively impairs adaptive responses to infection. Sci. Rep. 2018, 8, 3802. [Google Scholar] [CrossRef] [PubMed]
- Covre, L.P.; Martins, R.F.; Devine, O.P.; Chambers, E.S.; Vukmanovic-Stejic, M.; Silva, J.A.; Dietze, R.; Rodrigues, R.R.; de Matos Guedes, H.L.; Falqueto, A.; et al. Circulating senescent T cells are linked to systemic inflammation and lesion size during human cutaneous leishmaniasis. Front. Immunol. 2019, 9, 3001. [Google Scholar] [CrossRef] [PubMed]
- Covre, L.P.; Devine, O.P.; Garcia de Moura, R.; Vukmanovic-Stejic, M.; Dietze, R.; Ribeiro-Rodrigues, R.; Guedes, H.L.M.; Lubiana Zanotti, R.; Falqueto, A.; Akbar, A.N.; et al. Compartmentalized cytotoxic immune response leads to distinct pathogenic roles of natural killer and senescent CD8+ T cells in human cutaneous leishmaniasis. Immunology 2020, 159, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Fantecelle, C.H.; Covre, L.P.; Garcia de Moura, R.; Guedes, H.L.M.; Amorim, C.F.; Scott, P.; Mosser, D.; Falqueto, A.; Akbar, A.N.; Gomes, D.C.O. Transcriptomic landscape of skin lesions in cutaneous leishmaniasis reveals a strong CD8+ T cell immunosenescence signature linked to immunopathology. Immunology 2021, 164, 754–765. [Google Scholar] [CrossRef]
- Guimarães-Pinto, K.; Ferreira, J.R.M.; da Costa, A.L.A.; Morrot, A.; Freire-de-Lima, L.; Decote-Ricardo, D.; Freire-de-Lima, C.G.; Filardy, A.A. Cellular stress and senescence induction during Trypanosoma cruzi infection. Trop. Med. Infect. Dis. 2022, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Albareda, M.C.; Olivera, G.C.; Laucella, S.A.; Alvarez, M.G.; Fernandez, E.R.; Lococo, B.; Viotti, R.; Tarleton, R.L.; Postan, M. Chronic human infection with Trypanosoma cruzi drives CD4+ T cells to immune senescence. J. Immunol. 2009, 183, 4103–4108. [Google Scholar] [CrossRef] [PubMed]
- Froy, H.; Sparks, A.M.; Watt, K.; Sinclair, R.; Bach, F.; Pilkington, J.G.; Pemberton, J.M.; McNeilly, T.N.; Nussey, D.H. Senescence in immunity against helminth parasites predicts adult mortality in a wild mammal. Science 2019, 365, 1296–1298. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Pawelec, G.; Cohen, A.A.; Provost, G.; Khalil, A.; Lacombe, G.; Rodrigues, S.; Desroches, M.; Hirokawa, K.; et al. Immunosenescence and altered vaccine efficiency in older subjects: A myth difficult to change. Vaccines 2022, 10, 607. [Google Scholar] [CrossRef] [PubMed]
- Demicheli, V.; Jefferson, T.; Di Pietrantonj, C.; Ferroni, E.; Thorning, S.; Thomas, R.E.; Rivetti, A. Vaccines for preventing influenza in the elderly. Cochrane Database Syst. Rev. 2018, 2, CD004876. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Melgar, M.; Britton, A.; Roper, L.E.; Talbot, H.K.; Long, S.S.; Kotton, C.N.; Havers, F.P. Use of Respiratory Syncytial Virus Vaccines in Older Adults: Recommendations of the Advisory Committee on Immunization Practices—United States, 2023. MMWR Morb. Mortal. Wkly. Rep. 2023, 72, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, B.; Schirmer, M.; Matteucci Gothe, R.; Siebert, U.; Fuchs, D.; Grubeck-Loebenstein, B. Recall responses to tetanus and diphtheria vaccination are frequently insufficient in elderly persons. PLoS ONE 2013, 8, e82967. [Google Scholar] [CrossRef] [PubMed]
- Soegiarto, G.; Purnomosari, D. Challenges in the vaccination of the elderly and strategies for improvement. Pathophysiology 2023, 30, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Paladin, F.; Martino, G.; Gangemi, S. Impact of immunosenescence on viral Infections with an emphasis on COVID-19. Front. Biosci. 2023, 28, 225. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence and human vaccine immune responses. Immun. Ageing 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, C.E.; Kim, C.; Weyand, C.M.; Goronzy, J.J. Influence of immune aging on vaccine responses. J. Allergy Clin. Immunol. 2020, 145, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Linterman, M.A. Age-dependent changes in T follicular helper cells shape the humoral immune response to vaccination. Semin. Immunol. 2023, 69, 101801. [Google Scholar] [CrossRef] [PubMed]
- Coe, C.L.; Lubach, G.R.; Kinnard, J. Immune senescence in old and very old rhesus monkeys: Reduced antibody response to influenza vaccination. Age 2012, 34, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Murasko, D.M.; Bernstein, E.D.; Gardner, E.M.; Gross, P.; Munk, G.; Dran, S.; Abrutyn, E. Role of humoral and cell-mediated immunity in protection from influenza disease after immunization of healthy elderly. Exp. Gerontol. 2002, 37, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Ademokun, A.; Wu, Y.C.; Martin, V.; Mitra, R.; Sack, U.; Baxendale, H.; Kipling, D.; Dunn-Walters, D.K. Vaccination-induced changes in human B-cell repertoire and pneumococcal IgM and IgA antibody at different ages. Aging Cell 2011, 10, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Schenkein, J.G.; Nahm, M.H.; Dransfield, M.T. Pneumococcal vaccination for patients with COPD: Current practice and future directions. Chest 2008, 133, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Schenkein, J.G.; Park, S.; Nahm, M.H. Pneumococcal vaccination in older adults induces antibodies with low opsonic capacity and reduced antibody potency. Vaccine 2008, 26, 5521–5526. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Oxman, M.N.; Zhang, J.H.; Johnson, G.R.; Stanley, H.; Hayward, A.R.; Caulfield, M.J.; Irwin, M.R.; Smith, J.G.; Clair, J.; et al. Varicella-zoster virus-specific immune responses in elderly recipients of a herpes zoster vaccine. J. Infect. Dis. 2008, 197, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Arrazola Martínez, M.P.; Eiros Bouza, J.M.; Plans Rubió, P.; Puig-Barberà, J.; Ruiz Aragón, J.; Torres Lana, A.J. Efficacy, effectiveness and safety of the adjuvanted influenza vaccine in the population aged 65 or over. Rev. Esp. Quimioter. 2023, 36, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Ciabattini, A.; Nardini, C.; Santoro, F.; Garagnani, P.; Franceschi, C.; Medaglini, D. Vaccination in the elderly: The challenge of immune changes with aging. Semin. Immunol. 2018, 40, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; S Arunachalam, P.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Schuff-Werner, P. Challenges of laboratory diagnostics in the elderly. J. Lab. Med. 2018, 42, 105–107. [Google Scholar] [CrossRef]
- Agrawal, L.; Engel, K.B.; Greytak, S.R.; Moore, H.M. Understanding preanalytical variables and their effects on clinical biomarkers of oncology and immunotherapy. Semin. Cancer. Biol. 2018, 52, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Palleschi, L.; Galdi, F.; Pedone, C. Acute medical illness and disability in the elderly. Geriatr. Care 2018, 4, 7561. [Google Scholar] [CrossRef]
- Noppert, G.A.; Stebbins, R.C.; Dowd, J.B.; Aiello, A.E. Sociodemographic differences in population-level immunosenescence in older age. MedRxiv 2022. Preprint. [Google Scholar] [CrossRef]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: The path to the clinic. Nat. Med. 2022, 28, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- An Open-Label Intervention Trial to Reduce Senescence and Improve Frailty in Adult Survivors of Childhood Cancer. Available online: https://ncorp.cancer.gov/find-a-study/find-a-study.php?id=NCT04733534 (accessed on 18 March 2024).
- Targeting Pro-Inflammatory Cells in Idiopathic Pulmonary Fibrosis: A Human Trial (IPF). Available online: https://clinicaltrials.gov/study/NCT02874989 (accessed on 18 March 2024).
- Malavolta, M.; Giacconi, R.; Brunetti, D.; Provinciali, M.; Maggi, F. Exploring the relevance of senotherapeutics for the current SARS-CoV-2 emergency and similar future global health threats. Cells 2020, 9, 909. [Google Scholar] [CrossRef] [PubMed]
- Lelarge, V.; Capelle, R.; Oger, F.; Mathieu, T.; Le Calvé, B. Senolytics: From pharmacological inhibitors to immunotherapies, a promising future for patients’ treatment. NPJ Aging 2024, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, A.R.; Flores, M.K.; Miller, B.; Kim, S.J.; Yen, K.; Mehta, H.H.; Cohen, P. Mito-Omics and immune function: Applying novel mitochondrial omic techniques to the context of the aging immune system. Transl. Med. Aging 2020, 4, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Rafferty, E.; Paulden, M.; Buchan, S.A.; Robinson, J.L.; Bettinger, J.A.; Kumar, M.; Svenson, L.W.; MacDonald, S.E. Canadian Immunization Research Network (CIRN) investigators. Evaluating the individual healthcare costs and burden of disease associated with RSV across age groups. Pharmacoeconomics 2022, 40, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Mathiasen, S.L.; Gall-Mas, L.; Pateras, I.S.; Theodorou, S.D.P.; Namini, M.R.J.; Hansen, M.B.; Martin, O.C.B.; Vadivel, C.K.; Ntostoglou, K.; Butter, D.; et al. Bacterial genotoxins induce T cell senescence. Cell Rep. 2021, 35, 109220. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E.; Wieck, A.; Petersen, L.E.; Baptista, T.S. Neuroendocrine and viral correlates of premature immunosenescence. Ann. N. Y. Acad. Sci. 2015, 1351, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, T.T.; Dowrey, T.W.; Villacorta-Martin, C.; Montano, M.; Reed, E.; Belkina, A.C.; Andersen, S.L.; Perls, T.T.; Monti, S.; Murphy, G.J.; et al. Multi-modal profiling of peripheral blood cells across the human lifespan reveals distinct immune cell signatures of aging and longevity. EBioMedicine 2023, 90, 104514. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, S.K.; Manoharan, M.S.; Lee, G.C.; McKinnon, L.R.; Meunier, J.A.; Steri, M.; Harper, N.; Fiorillo, E.; Smith, A.M.; Restrepo, M.I.; et al. Immune resilience despite inflammatory stress promotes longevity and favorable health outcomes including resistance to infection. Nat. Commun. 2023, 14, 3286. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, M.J.; Melos, K.I.; Angelini, L.; Burd, C.E.; Robbins, P.D.; Niedernhofer, L.J. Mouse models of accelerated cellular senescence. Methods Mol. Biol. 2019, 1896, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.L.; Bakula, D.; Scheibye-Knudsen, M. Clinical Trials Targeting Aging. Front. Aging 2022, 3, 820215. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Infective Agents | Innate Immunity | Adaptive Immunity | |||
|---|---|---|---|---|---|
| References | References | ||||
| viral | HIV | IL-6 and TNF-α elevation | [116,118] | decreased thymic output | [37,119,120] |
| IFN-α increase and IL-2 reduction | expansion of TEMRA cells | [88] | |||
| expansion of non-functional CD3-CD56-CD16+ NK cells | [125] | elevated KRECs in patients not needing therapy | [121] | ||
| decreased T-cell diversity | [123] | ||||
| diminished CD8+ T-cell response in aged mice | [135] | ||||
| reduced public CD8+ TCRαβ clonotypes and TCRαβ diversity | [137] | ||||
| Influenza virus | decrease in macrophage peritoneal phagocytic function | [131,132] | |||
| reduced uptake of bacteria by monocytes | [133] | ||||
| decreased abundance, activity and migration of DCs | [134] | ||||
| RSV | low levels of serum neutralizing antibody and IFN-γ | [141] | RSV-specific effector memory T cells prevent symptomatic infection | [142] | |
| CMV | increased CD8+ CD244+ effector T cells | [147] | |||
| decreased CD16-/CD16+CD56bright and increase in CD56−CD16+ NK cells percentage | [146] | lower number of naïve CD4+ and increased effector memory CD4+ and CD8+ T cells | [148] | ||
| reduced TCR repertoire diversity | [151] | ||||
| EBV | expansion of CD56- NK cells with reduced cytotoxic capacity and IFN-γ production | [152] | increase in terminally differentiated T cells and decrease in TCR repertoire diversity | [153] | |
| expansion of viral-specific exhausted, senescent CD8+ CD28− T cells | [154] | ||||
| HCV | high plasma levels of SASP proteins | [155] | increase in intrahepatic senescent, not functional T cells | [156,157] | |
| HZV | interferes with the type 1 IFN pathway and the production of pro-inflammatory cytokines | [160] | reduced frequency of virus-specific effector memory T cells | [161] | |
| increases in CD57+ NK cells | [159] | ||||
| Measles virus and parvovirus | induction of pro-inflammatory secretome-related factors | [164] | |||
| WWNV | impairment of neutrophils, monocyte/macrophages, DCs, and NK cells | [169] | |||
| JCV | CD4+ T-cell lymphocytopenia, low production of TRECs and KRECs and TCR repertoire restrictions in natalizumab-treated patients | [172,173,174] | |||
| bacterial | Mycobacterium tuberculosis | alterations in monocyte proportion and phenotype | [186] | impaired adaptive T-cell immunity | [118,186] |
| reduction in IFN-γ/IL-4 ratio and other pro-inflammatory, such as IL-17A, IL-2, TNF-α | [187] | reduction in regulatory T cells and polyfunctional IFN-γ+TNF-α+ T cells | [187] | ||
| imbalanced pro- and anti-inflammatory factor pattern and changes in IL-2 and TNF-α production in the lung | [184] | ||||
| Streptococcus pneumoniae | low opsonic activities of antibodies and phagocytic killing of neutrophils | [191] | changes in CD27+IgM+ B cells | [194] | |
| increase in senescence markers (IL-1α/β, TNF-α, IL-6, and CXCL1) | [192] | ||||
| Escherichia coli and other bacteria inducing urinary tract infections | high levels of pro-inflammatory cytokines (IL-6, IL-1β, and TNF-α) | [195] | formation of bladder tertiary lymphoid tissues and redistribution of B-cell pools from the periphery to mucosal surface that alter the mucosal landscape | [195] | |
| aged bladder CXCL13+ macrophages may be responsible for inhibiting development of the adaptive immune responses | [198] | ||||
| decreased macrophage phagocytosis | [199] | ||||
| Gram-positive and Gram-negative intestinal bacteria disequilibrium | activation of DCs | [205] | |||
| release of pro-inflammatory cytokines, mainly IL-6 and IL-17 | [206] | ||||
| Porphyromonas gingivalis | senescent cellular markers in DCs | [214] | |||
| Gram-positive and Gram-negative induced sepsis | expansion of myeloid-derived suppressor cells, inhibiting the function of DCs and macrophages in cirrhosis patients | [222] | inhibition of Th1 response and induction of Th2 and regulatory T-cell productions | [222] | |
| parasitic | Leishmania | expansion of senescent CD56+ CD57+ NK cells | [231] | expansion of CD57+ CD4+ lymphocytes | [230] |
| expansion of effector memory CD8+ T cells that re-express CD45RA marker | [231] | ||||
| increased transcriptions of senescence-associated genes in the cutaneous lesions | [232] | ||||
| Trypanosoma cruzi | compromised capacity to control the magnitude of inflammation | [233] | increase in antigen-experienced IFN-γ-producing CD4+ T cells | [234] | |
| Helminths | compromised Th2 function in mice | [235] | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiros-Roldan, E.; Sottini, A.; Natali, P.G.; Imberti, L. The Impact of Immune System Aging on Infectious Diseases. Microorganisms 2024, 12, 775. https://doi.org/10.3390/microorganisms12040775
Quiros-Roldan E, Sottini A, Natali PG, Imberti L. The Impact of Immune System Aging on Infectious Diseases. Microorganisms. 2024; 12(4):775. https://doi.org/10.3390/microorganisms12040775
Chicago/Turabian StyleQuiros-Roldan, Eugenia, Alessandra Sottini, Pier Giorgio Natali, and Luisa Imberti. 2024. "The Impact of Immune System Aging on Infectious Diseases" Microorganisms 12, no. 4: 775. https://doi.org/10.3390/microorganisms12040775
APA StyleQuiros-Roldan, E., Sottini, A., Natali, P. G., & Imberti, L. (2024). The Impact of Immune System Aging on Infectious Diseases. Microorganisms, 12(4), 775. https://doi.org/10.3390/microorganisms12040775