

How Does Airway Surface Liquid Composition Vary in Different Pulmonary Diseases, and How Can We Use This Knowledge to Model Microbial Infections?


Abstract
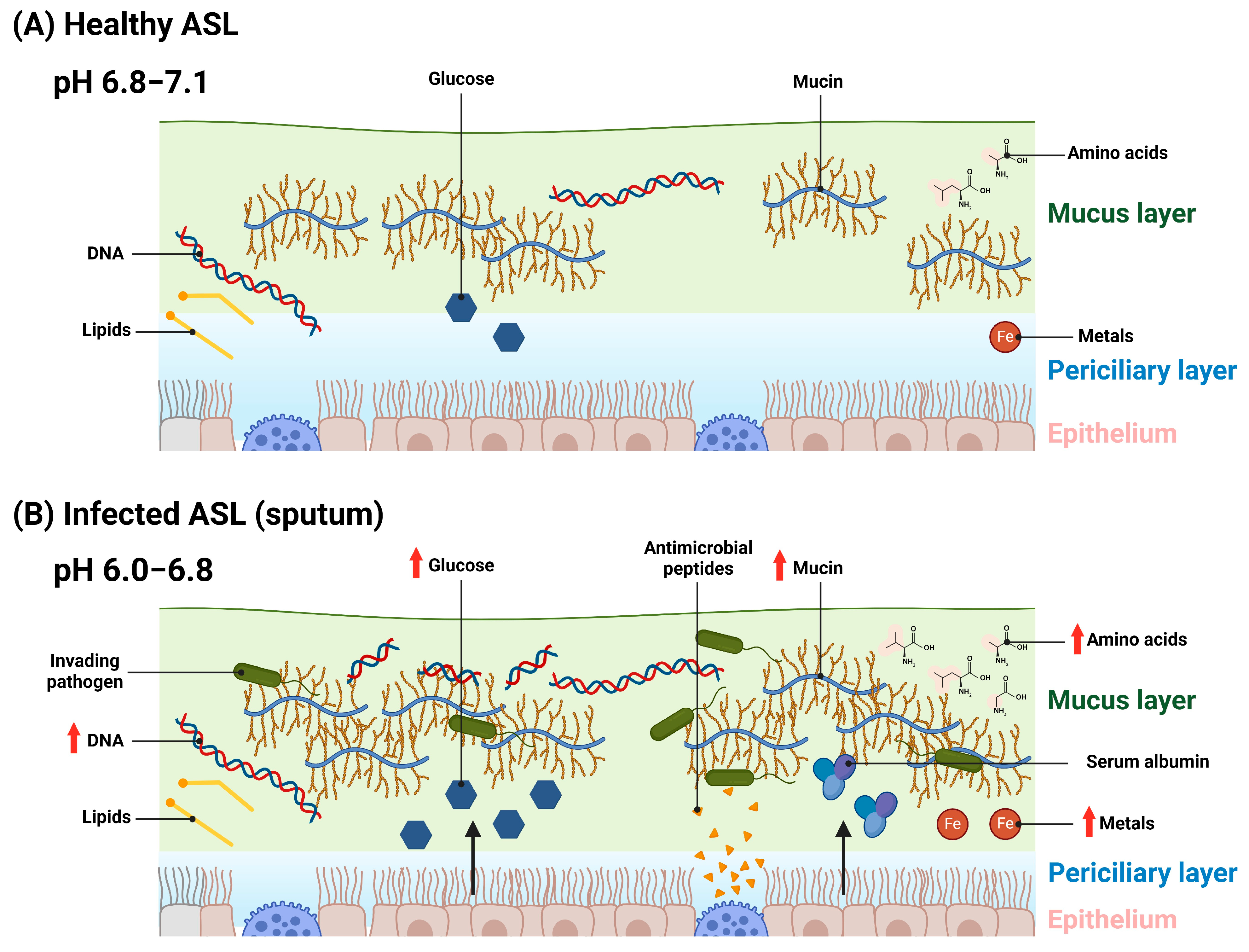
1. Introduction
2. Mucin
3. DNA
4. pH and Ion Concentrations
5. Sugars and Other Carbon Sources
6. Amino Acids
7. Lipids
8. Antimicrobial Peptides and Enzymes
9. Metals
10. Polyamines
11. Serum Albumin
12. Airway Surface Liquid Component Interactions
13. Lung Microbiome
14. Future Developments and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASL | Airway surface liquid |
| PCL | Periciliary layer |
| MCL | Mucus layer |
| CF | Cystic fibrosis |
| COPD | Chronic obstructive pulmonary disease |
| VAP | Ventilator-associated pneumonia |
| ETT | Endotracheal tube |
| COVID-19 | Coronavirus disease 2019 |
| ICU | Intensive care unit |
| PGM | Porcine gastric mucin |
| BSM | Bovine submaxillary mucin |
| NET | Neutrophil extracellular trap |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| ASM | Artificial sputum medium |
| SCFM | Synthetic CF mucus media |
| ARDS | Acute respiratory distress syndrome |
| MRSA | Methicillin-resistant S. aureus |
| GlcNAc | N-acetylglucosamine |
| GalNAc | N-acetylgalactose |
| NTHi | Nontypeable Haemophilus influenzae |
| BALF | Bronchoalveolar lavage fluid |
| HNE | Human neutrophil elastase |
| MMP | Matrix metalloproteases |
| BSA | Bovine serum albumin |
References
- Widdicombe, J.H. Regulation of the depth and composition of airway surface liquid. J. Anat. 2002, 201, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M.; Dreano, E.; Edwards, A.; Planelles, G.; Sermet-Gaudelus, I. Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge. Int. J. Mol. Sci. 2021, 22, 3384. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajah, J.R.; Song, Y.; Derichs, N.; Verkman, A.S. Airway surface liquid depth imaged by surface laser reflectance microscopy. J. Gen. Physiol. 2010, 136, 353–362. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Atanasova, K.R.; Reznikov, L.R. Strategies for measuring airway mucus and mucins. Respir. Res. 2019, 20, 261. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, K.R.; O’Sullivan, D.D.; Seltzer, J.; Rossing, T.H.; Drazen, J.M.; Reid, L.M. Density gradient study of bronchial mucus aspirates from healthy volunteers (smokers and nonsmokers) and from patients with tracheostomy. Exp. Lung Res. 1985, 9, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.R.; Boucher, R.C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Investig. 2002, 109, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Rostami, M.R.; LeBlanc, M.G.; Strulovici-Barel, Y.; Zuo, W.; Mezey, J.G.; O’Beirne, S.L.; Kaner, R.J.; Leopold, P.L.; Crystal, R.G. Smoking shifts human small airway epithelium club cells toward a lesser differentiated population. NPJ. Genom. Med. 2021, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Wanner, A.; Salathe, M.; O’Riordan, T.G. Mucociliary clearance in the airways. Am. J. Respir. Crit. Care. Med. 1996, 154, 1868–1902. [Google Scholar] [CrossRef] [PubMed]
- Roomans, G.M.; Kozlova, I.; Nilsson, H.; Vanthanouvong, V.; Button, B.; Tarran, R. Measurements of airway surface liquid height and mucus transport by fluorescence microscopy, and of ion composition by X-ray microanalysis. J. Cyst. Fibros. 2004, 3 (Suppl. S2), 135–139. [Google Scholar] [CrossRef]
- King, M.; Zahm, J.M.; Pierrot, D.; Vaquez-Girod, S.; Puchelle, E. The role of mucus gel viscosity, spinnability, and adhesive properties in clearance by simulated cough. Biorheology 1989, 26, 737–745. [Google Scholar] [CrossRef]
- Amatngalim, G.D.; Hiemstra, P.S. Airway Epithelial Cell Function and Respiratory Host Defense in Chronic Obstructive Pulmonary Disease. Chin. Med. J. 2018, 131, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Mager, S.; Sloan, J. Possible role of amino acids, peptides, and sugar transporter in protein removal and innate lung defense. Eur. J. Pharmacol. 2003, 479, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, A.A.; Gutiérrez, J.; Duschner, K.S.; McConnell, K.S.; Taft, P.J.; Ernst, S.E.; Yahr, T.L.; Rahmouni, K.; Klesney-Tait, J.; Stoltz, D.A.; et al. Glucose Depletion in the Airway Surface Liquid Is Essential for Sterility of the Airways. PLoS ONE 2011, 6, e16166. [Google Scholar] [CrossRef] [PubMed]
- Vargas Buonfiglio, L.G.; Borcherding, J.A.; Frommelt, M.; Parker, G.J.; Duchman, B.; Vanegas Calderon, O.G.; Fernandez-Ruiz, R.; Noriega, J.E.; Stone, E.A.; Gerke, A.K.; et al. Airway surface liquid from smokers promotes bacterial growth and biofilm formation via iron-lactoferrin imbalance. Respir. Res. 2018, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.; Garnett, J.P.; Mather, M.W.; Cooles, F.A.H.; Nelson, A.; Verdon, B.; Scott, J.; Jiwa, K.; Ruchaud-Sparagano, M.H.; Cummings, S.P.; et al. Excess Mucin Impairs Subglottic Epithelial Host Defense in Mechanically Ventilated Patients. Am. J. Respir. Crit. Care Med. 2018, 198, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Palmer, K.L.; Aye, L.M.; Whiteley, M. Nutritional cues control Pseudomonas aeruginosa multicellular behavior in cystic fibrosis sputum. J. Bacteriol. 2007, 189, 8079–8087. [Google Scholar] [CrossRef] [PubMed]
- Stickler, D.J.; Morris, N.S.; Winters, C. Simple physical model to study formation and physiology of biofilms on urethral catheters. Methods. Enzymol. 1999, 310, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Pratten, J.; Smith, A.W.; Wilson, M. Response of single species biofilms and microcosm dental plaques to pulsing with chlorhexidine. J. Antimicrob. Chemother. 1998, 42, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Werthen, M.; Henriksson, L.; Jensen, P.O.; Sternberg, C.; Givskov, M.; Bjarnsholt, T. An in vitro model of bacterial infections in wounds and other soft tissues. APMIS 2010, 118, 156–164. [Google Scholar] [CrossRef]
- Maierl, M.; Jorger, M.; Rosker, P.; Reisner, A. In vitro Dynamic Model of a Catheterized Bladder and Biofilm Assay. Bio. Protoc. 2015, 5, e1381. [Google Scholar] [CrossRef] [PubMed]
- Millhouse, E.; Jose, A.; Sherry, L.; Lappin, D.F.; Patel, N.; Middleton, A.M.; Pratten, J.; Culshaw, S.; Ramage, G. Development of an in vitroperiodontal biofilm model for assessing antimicrobial and host modulatory effects of bioactive molecules. BMC. Oral Health 2014, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Furner-Pardoe, J.; Anonye, B.O.; Cain, R.; Moat, J.; Ortori, C.A.; Lee, C.; Barrett, D.A.; Corre, C.; Harrison, F. Anti-biofilm efficacy of a medieval treatment for bacterial infection requires the combination of multiple ingredients. Sci. Rep. 2020, 10, 12687. [Google Scholar] [CrossRef] [PubMed]
- Harrington, N.E.; Sweeney, E.; Harrison, F. Building a better biofilm-Formation of in vivo-like biofilm structures by Pseudomonas aeruginosa in a porcine model of cystic fibrosis lung infection. Biofilm 2020, 2, 100024. [Google Scholar] [CrossRef] [PubMed]
- Fung, C.; Naughton, S.; Turnbull, L.; Tingpej, P.; Rose, B.; Arthur, J.; Hu, H.; Harmer, C.; Harbour, C.; Hassett, D.J.; et al. Gene expression of Pseudomonas aeruginosa in a mucin-containing synthetic growth medium mimicking cystic fibrosis lung sputum. J. Med. Microbiol. 2010, 59, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Hare, N.J.; Soe, C.Z.; Rose, B.; Harbour, C.; Codd, R.; Manos, J.; Cordwell, S.J. Proteomics of Pseudomonas aeruginosa Australian epidemic strain 1 (AES-1) cultured under conditions mimicking the cystic fibrosis lung reveals increased iron acquisition via the siderophore pyochelin. J. Proteome Res. 2012, 11, 776–795. [Google Scholar] [CrossRef] [PubMed]
- Ruhluel, D.; O’Brien, S.; Fothergill, J.L.; Neill, D.R. Development of liquid culture media mimicking the conditions of sinuses and lungs in cystic fibrosis and health. F1000Research 2022, 11, 1007. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, S.D. Artificial Sputum Medium. Protoc. Exch. 2010. [Google Scholar] [CrossRef]
- Ghani, M.; Soothill, J.S. Ceftazidime, gentamicin, and rifampicin, in combination, kill biofilms of mucoid Pseudomonas aeruginosa. Can. J. Microbiol. 1997, 43, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, S.; Fothergill, J.L.; Wright, E.A.; James, C.E.; Mowat, E.; Winstanley, C. Use of artificial sputum medium to test antibiotic efficacy against Pseudomonas aeruginosa in conditions more relevant to the cystic fibrosis lung. J. Vis. Exp. 2012, 64, e3857. [Google Scholar] [CrossRef]
- Quinn, R.A.; Whiteson, K.; Lim, Y.W.; Salamon, P.; Bailey, B.; Mienardi, S.; Sanchez, S.E.; Blake, D.; Conrad, D.; Rohwer, F. A Winogradsky-based culture system shows an association between microbial fermentation and cystic fibrosis exacerbation. ISME 2015, 9, 1024–1038. [Google Scholar] [CrossRef]
- Sriramulu, D.D.; Lunsdorf, H.; Lam, J.S.; Romling, U. Microcolony formation: A novel biofilm model of Pseudomonas aeruginosa for the cystic fibrosis lung. J. Med. Microbiol. 2005, 54, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Serisier, D.J.; Carroll, M.P.; Shute, J.K.; Young, S.A. Macrorheology of cystic fibrosis, chronic obstructive pulmonary disease & normal sputum. Respir. Res. 2009, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V.; Kim, K.W.; Liu, J.; Boushey, H.A. Prominent neutrophilic inflammation in sputum from subjects with asthma exacerbation. J. Allergy. Clin. Immunol. 1995, 95, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Imam, J.S.; Duarte, A.G. Non-CF bronchiectasis: Orphan disease no longer. Respir. Med. 2020, 166, 105940. [Google Scholar] [CrossRef] [PubMed]
- Eickmeier, O.; Huebner, M.; Herrmann, E.; Zissler, U.; Rosewich, M.; Baer, P.C.; Buhl, R.; Schmitt-Grohé, S.; Zielen, S.; Schubert, R. Sputum biomarker profiles in cystic fibrosis (CF) and chronic obstructive pulmonary disease (COPD) and association between pulmonary function. Cytokine 2010, 50, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.K. Mucus, phlegm, and sputum in cystic fibrosis. Respir. Care 2009, 54, 726–732, discussion 732. [Google Scholar] [CrossRef] [PubMed]
- Koenig, S.M.; Truwit, J.D. Ventilator-associated pneumonia: Diagnosis, treatment, and prevention. Clin. Microbiol. Rev. 2006, 19, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Charles, P.M.V.; Kali, A.; Easow, J.M.; Joseph, N.M.; Ravishankar, M.; Srinivasan, S.; Kumar, S.; Umadevi, S. Ventilator-associated pneumonia. Australas. Med. J. 2014, 7, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Bhagat, R. Preventing Ventilator-Associated Infections. Clin. Chest. Med. 2016, 37, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Nora, D.; Povoa, P. Antibiotic consumption and ventilator-associated pneumonia rates, some parallelism but some discrepancies. Ann. Transl. Med. 2017, 5, 450. [Google Scholar] [CrossRef]
- Camara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Economic significance of biofilms: A multidisciplinary and cross-sectoral challenge. NPJ Biofilms Microbiomes 2022, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Gragueb-Chatti, I.; Hyvernat, H.; Leone, M.; Agard, G.; Peres, N.; Guervilly, C.; Boucekine, M.; Hamidi, D.; Papazian, L.; Dellamonica, J.; et al. Incidence, Outcomes and Risk Factors of Recurrent Ventilator Associated Pneumonia in COVID-19 Patients: A Retrospective Multicenter Study. J. Clin. Med. 2022, 11, 7097. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.; Grainha, T.; Pereira, M.O.; Lopes, S.P. Antimicrobial materials for endotracheal tubes: A review on the last two decades of technological progress. Acta Biomater. 2023, 158, 32–55. [Google Scholar] [CrossRef] [PubMed]
- Papazian, L.; Klompas, M.; Luyt, C.E. Ventilator-associated pneumonia in adults: A narrative review. Intensive Care Med. 2020, 46, 888–906. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Yamamoto, Y.; Yanagihara, K.; Araki, N.; Harada, Y.; Morinaga, Y.; Izumikawa, K.; Kakeya, H.; Hasegawa, H.; Kohno, S.; et al. In vivo efficacy and pharmacokinetics of biapenem in a murine model of ventilator-associated pneumonia with Pseudomonas aeruginosa. J. Infect. Chemother. 2012, 18, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.M.; Sibila, O.; Agusti, C.; Torres, A. Animal models of ventilator-associated pneumonia. Eur. Respir. J. 2009, 33, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Benahmed, M.A.; Elbayed, K.; Daubeuf, F.; Santelmo, N.; Frossard, N.; Namer, I.J. NMR HRMAS spectroscopy of lung biopsy samples: Comparison study between human, pig, rat, and mouse metabolomics. Magn. Reson. Med. 2014, 71, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, I.; Vanthanouvong, V.; Almgren, B.; Hogman, M.; Roomans, G.M. Elemental composition of airway surface liquid in the pig determined by x-ray microanalysis. Am. J. Respir. Cell Mol. Biol. 2005, 32, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Cowley, E.A.; Govindaraju, K.; Guilbault, C.; Radzioch, D.; Eidelman, D.H. Airway surface liquid composition in mice. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2000, 278, L1213–L1220. [Google Scholar] [CrossRef]
- Robinson, N.P.; Kyle, H.; Webber, S.E.; Widdicombe, J.G. Electrolyte and other chemical concentrations in tracheal airway surface liquid and mucus. J. Appl. Physiol. 1989, 66, 2129–2135. [Google Scholar] [CrossRef]
- Boucher, R.C.; Stutts, M.J.; Bromberg, P.A.; Gatzy, J.T. Regional differences in airway surface liquid composition. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Thiagarajah, J.; Verkman, A.S. Sodium and chloride concentrations, pH, and depth of airway surface liquid in distal airways. J. Gen. Physiol. 2003, 122, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef] [PubMed]
- Boat, T.F.; Cheng, P.W.; Iyer, R.N.; Carlson, D.M.; Polony, I. Human respiratory tract secretion: Mucous glycoproteins of nonpurulent tracheobronchial secretions, and sputum of patients with bronchitis and cystic fibrosis. Arch. Biochem. Biophys. 1977, 177, 97–104. [Google Scholar]
- Henke, M.O.; John, G.; Germann, M.; Lindemann, H.; Rubin, B.K. MUC5AC and MUC5B mucins increase in cystic fibrosis airway secretions during pulmonary exacerbation. Am. J. Respir. Crit. Care Med. 2007, 175, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, S.; Sheehan, J.K.; Knight, D.; Richardson, P.S.; Thornton, D.J. Heterogeneity of airways mucus: Variations in the amounts and glycoforms of the major oligomeric mucins MUC5AC and MUC5B. Biochem. J. 2002, 361, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Wickstrom, C.; Davies, J.R.; Eriksen, G.V.; Veerman, E.C.; Carlstedt, I. MUC5B is a major gel-forming, oligomeric mucin from human salivary gland, respiratory tract and endocervix: Identification of glycoforms and C-terminal cleavage. Biochem. J. 1998, 334, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Royall, J.A.; Damera, G.; Sachdev, G.P.; Cummings, R.D. Altered O-glycosylation and sulfation of airway mucins associated with cystic fibrosis. Glycobiology 2005, 15, 747–775. [Google Scholar] [CrossRef] [PubMed]
- Patarin, J.; Ghiringhelli, E.; Darsy, G.; Obamba, M.; Bochu, P.; Camara, B.; Quetant, S.; Cracowski, J.L.; Cracowski, C.; Robert de Saint Vincent, M. Rheological analysis of sputum from patients with chronic bronchial diseases. Sci. Rep. 2020, 10, 15685. [Google Scholar] [CrossRef]
- Henke, M.O.; Renner, A.; Huber, R.M.; Seeds, M.C.; Rubin, B.K. MUC5AC and MUC5B Mucins Are Decreased in Cystic Fibrosis Airway Secretions. Am. J. Respir. Cell Mol. Biol. 2004, 31, 86–91. [Google Scholar] [CrossRef]
- Henderson, A.G.; Ehre, C.; Button, B.; Abdullah, L.H.; Cai, L.H.; Leigh, M.W.; DeMaria, G.C.; Matsui, H.; Donaldson, S.H.; Davis, C.W.; et al. Cystic fibrosis airway secretions exhibit mucin hyperconcentration and increased osmotic pressure. J. Clin. Investig. 2014, 124, 3047–3060. [Google Scholar] [CrossRef] [PubMed]
- Hellyer, T.P.; Morris, A.C.; McAuley, D.F.; Walsh, T.S.; Anderson, N.H.; Singh, S.; Dark, P.; Roy, A.I.; Baudouin, S.V.; Wright, S.E.; et al. Diagnostic accuracy of pulmonary host inflammatory mediators in the exclusion of ventilator-acquired pneumonia. Thorax 2015, 70, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Gernez, Y.; Tirouvanziam, R.; Chanez, P. Neutrophils in chronic inflammatory airway diseases: Can we target them and how? Eur. Respir. J. 2010, 35, 467–469. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, S.; Kolsum, U.; Rousseau, K.; Singh, D.; Vestbo, J.; Thornton, D.J. MUC5B Is the Major Mucin in the Gel Phase of Sputum in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2008, 178, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Bassi, G.L.; Zanella, A.; Cressoni, M.; Stylianou, M.; Kolobow, T. Following tracheal intubation, mucus flow is reversed in the semirecumbent position: Possible role in the pathogenesis of ventilator-associated pneumonia. Crit. Care Med. 2008, 36, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, C.L.; Lalsiamthara, J.; Aballay, A. Host Mucin Is Exploited by Pseudomonas aeruginosa to Provide Monosaccharides Required for a Successful Infection. mBio 2020, 11, e00060-20. [Google Scholar] [CrossRef] [PubMed]
- Yesilkaya, H.; Manco, S.; Kadioglu, A.; Terra, V.S.; Andrew, P.W. The ability to utilize mucin affects the regulation of virulence gene expression in Streptococcus pneumoniae. FEMS Microbiol. Lett. 2008, 278, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, V.; Packer, N.H.; Thaysen-Andersen, M. Host mucin glycosylation plays a role in bacterial adhesion in lungs of individuals with cystic fibrosis. Expert Rev. Respir. Med. 2013, 7, 553–576. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, K.M.; Carcamo-Oyarce, G.; Turner, B.S.; Dellos-Nolan, S.; Co, J.Y.; Lehoux, S.; Cummings, R.D.; Wozniak, D.J.; Ribbeck, K. Mucin glycans attenuate the virulence of Pseudomonas aeruginosa in infection. Nat. Microbiol. 2019, 4, 2146–2154. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.X.; Wheeler, K.M.; Cady, K.C.; Lehoux, S.; Cummings, R.D.; Laub, M.T.; Ribbeck, K. Mucin Glycans Signal through the Sensor Kinase RetS to Inhibit Virulence-Associated Traits in Pseudomonas aeruginosa. Curr. Biol. 2021, 31, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.E.; Wheeler, K.M.; Ribbeck, K. Mucins and Their Role in Shaping the Functions of Mucus Barriers. Annu. Rev. Cell Dev. Biol. 2018, 34, 189–215. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.L.; Sloane, A.J.; Robinson, L.J.; Prasad, S.S.; Lindner, R.A.; Robinson, M.; Bye, P.T.; Nielson, D.W.; Harry, J.L.; Packer, N.H.; et al. Glycosylation of sputum mucins is altered in cystic fibrosis patients. Glycobiology 2007, 17, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.K.; Wang, Y.Y.; Wirtz, D.; Hanes, J. Micro- and macrorheology of mucus. Adv. Drug Deliv. Rev. 2009, 61, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.M.; Niccum, D.; Dunitz, J.M.; Hunter, R.C. Evidence and Role for Bacterial Mucin Degradation in Cystic Fibrosis Airway Disease. PLoS Pathog. 2016, 12, e1005846. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Schaefer, A.L.; Parsek, M.R.; Moninger, T.O.; Welsh, M.J.; Greenberg, E.P. Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 2000, 407, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The Interaction between Respiratory Pathogens and Mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Paharik, A.E.; Salgado-Pabon, W.; Meyerholz, D.K.; White, M.J.; Schlievert, P.M.; Horswill, A.R. The Spl Serine Proteases Modulate Staphylococcus aureus Protein Production and Virulence in a Rabbit Model of Pneumonia. mSphere 2016, 1, e00208-16. [Google Scholar] [CrossRef] [PubMed]
- Takagi, J.; Aoki, K.; Turner, B.S.; Lamont, S.; Lehoux, S.; Kavanaugh, N.; Gulati, M.; Valle Arevalo, A.; Lawrence, T.J.; Kim, C.Y.; et al. Mucin O-glycans are natural inhibitors of Candida albicans pathogenicity. Nat. Chem. Biol. 2022, 18, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Ahearn, C.P.; Gallo, M.C.; Murphy, T.F. Insights on persistent airway infection by non-typeable Haemophilus influenzae in chronic obstructive pulmonary disease. Pathog. Dis. 2017, 75, ftx042. [Google Scholar] [CrossRef]
- Turner, K.H.; Wessel, A.K.; Palmer, G.C.; Murray, J.L.; Whiteley, M. Essential genome of Pseudomonas aeruginosa in cystic fibrosis sputum. Proc. Natl. Acad. Sci. USA 2015, 112, 4110–4115. [Google Scholar] [CrossRef] [PubMed]
- Padra, M.; Adamczyk, B.; Benktander, J.; Flahou, B.; Skoog, E.C.; Padra, J.T.; Smet, A.; Jin, C.; Ducatelle, R.; Samuelsson, T.; et al. Helicobacter suis binding to carbohydrates on human and porcine gastric mucins and glycolipids occurs via two modes. Virulence 2018, 9, 898–918. [Google Scholar] [CrossRef] [PubMed]
- Barmpatsalou, V.; Dubbelboer, I.R.; Rodler, A.; Jacobson, M.; Karlsson, E.; Pedersen, B.L.; Bergstrom, C.A.S. Physiological properties, composition and structural profiling of porcine gastrointestinal mucus. Eur. J. Pharm. Biopharm. 2021, 169, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, K.; Kirkham, S.; Johnson, L.; Fitzpatrick, B.; Howard, M.; Adams, E.J.; Rogers, D.F.; Knight, D.; Clegg, P.; Thornton, D.J. Proteomic analysis of polymeric salivary mucins: No evidence for MUC19 in human saliva. Biochem. J. 2008, 413, 545–552. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lieleg, O.; Lieleg, C.; Bloom, J.; Buck, C.B.; Ribbeck, K. Mucin biopolymers as broad-spectrum antiviral agents. Biomacromolecules 2012, 13, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.; Holbein, B.E.; Zhou, H.; Xu, H.H.; Chen, W. Potential Mechanisms of Mucin-Enhanced Acinetobacter baumannii Virulence in the Mouse Model of Intraperitoneal Infection. Infect. Immun. 2019, 87, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Neve, R.L.; Carrillo, B.D.; Phelan, V.V. Impact of Artificial Sputum Medium Formulation on Pseudomonas aeruginosa Secondary Metabolite Production. J. Bacteriol. 2021, 203, e0025021. [Google Scholar] [CrossRef] [PubMed]
- Nisizawa, K.; Pigman, W. The composition and properties of the mucin clot from cattle submaxillary glands. Arch. Oral Biol. 1959, 1, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Tettamanti, G.; Pigman, W. Purification and characterization of bovine and ovine submaxillary mucins. Arch. Biochem. Biophys. 1968, 124, 41–50. [Google Scholar] [CrossRef]
- Schömig, V.J.; Käsdorf, B.T.; Scholz, C.; Bidmon, K.; Lieleg, O.; Berensmeier, S. An optimized purification process for porcine gastric mucin with preservation of its native functional properties. RSC Adv. 2016, 6, 44932–44943. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit. Care 2018, 22, 358. [Google Scholar] [CrossRef] [PubMed]
- Radermecker, C.; Detrembleur, N.; Guiot, J.; Cavalier, E.; Henket, M.; d’Emal, C.; Vanwinge, C.; Cataldo, D.; Oury, C.; Delvenne, P.; et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J. Exp. Med. 2020, 217, e20201012. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Dassler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J. Exp. Med. 2020, 217, e20200652. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.N.; Stein, R.T. Neutrophil Extracellular Traps in Pulmonary Diseases: Too Much of a Good Thing? Front. Immunol. 2016, 7, 311. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, C.; Palaniyar, N.; Otulakowski, G.; Khan, M.A.; Post, M.; Kuebler, W.M.; Tanswell, K.; Belcastro, R.; Masood, A.; Engelberts, D.; et al. Mechanical ventilation induces neutrophil extracellular trap formation. Anesthesiology 2015, 122, 864–875. [Google Scholar] [CrossRef] [PubMed]
- De Rose, V. Mechanisms and markers of airway inflammation in cystic fibrosis. Eur. Respir. J. 2002, 19, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Rosenecker, J.; Naundorf, S.; Rudolph, C. Airway surface liquid contains endogenous DNase activity which can be activated by exogenous magnesium. Eur. J. Med. Res. 2009, 14, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Lethem, M.I.; James, S.L.; Marriott, C.; Burke, J.F. The origin of DNA associated with mucus glycoproteins in cystic fibrosis sputum. Eur. Respir. J. 1990, 3, 19–23. [Google Scholar] [CrossRef]
- Lewenza, S.; Johnson, L.; Charron-Mazenod, L.; Hong, M.; Mulcahy-O’Grady, H. Extracellular DNA controls expression of Pseudomonas aeruginosa genes involved in nutrient utilization, metal homeostasis, acid pH tolerance and virulence. J. Med. Microbiol. 2020, 69, 895–905. [Google Scholar] [CrossRef]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA Acidifies Biofilms and Induces Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Herzog, S.; Dach, F.; de Buhr, N.; Niemann, S.; Schlagowski, J.; Chaves-Moreno, D.; Neumann, C.; Goretzko, J.; Schwierzeck, V.; Mellmann, A.; et al. High Nuclease Activity of Long Persisting Staphylococcus aureus Isolates Within the Airways of Cystic Fibrosis Patients Protects Against NET-Mediated Killing. Front. Immunol. 2019, 10, 2552. [Google Scholar] [CrossRef] [PubMed]
- Brandt, T.; Breitenstein, S.; von der Hardt, H.; Tummler, B. DNA concentration and length in sputum of patients with cystic fibrosis during inhalation with recombinant human DNase. Thorax 1995, 50, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, K.K.; Wagener, J.S.; Khan, T.Z.; Copenhaver, S.C.; Accurso, F.J. Increased DNA levels in bronchoalveolar lavage fluid obtained from infants with cystic fibrosis. Am. J. Respir. Crit. Care Med. 1996, 154, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Marcos, V.; Zhou-Suckow, Z.; Onder Yildirim, A.; Bohla, A.; Hector, A.; Vitkov, L.; Krautgartner, W.D.; Stoiber, W.; Griese, M.; Eickelberg, O.; et al. Free DNA in cystic fibrosis airway fluids correlates with airflow obstruction. Mediat. Inflamm. 2015, 2015, 408935. [Google Scholar] [CrossRef] [PubMed]
- McShane, D.; Davies, J.C.; Davies, M.G.; Bush, A.; Geddes, D.M.; Alton, E.W. Airway surface pH in subjects with cystic fibrosis. Eur. Respir. J. 2003, 21, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Song, Y.; Vetrivel, L.; Shankar, L.; Verkman, A.S. Noninvasive in vivo fluorescence measurement of airway-surface liquid depth, salt concentration, and pH. J. Clin. Investig. 2001, 107, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Salinas, D.; Nielson, D.W.; Verkman, A.S. Hyperacidity of secreted fluid from submucosal glands in early cystic fibrosis. Am. J. Physiol.-Cell Physiol. 2006, 290, C741–C749. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Joo, N.S.; Reitz, B.; Wine, J.J.; Verkman, A.S. Submucosal gland secretions in airways from cystic fibrosis patients have normal [Na+] and pH but elevated viscosity. Proc. Natl. Acad. Sci. USA 2001, 98, 8119–8123. [Google Scholar] [CrossRef]
- Bodem, C.R.; Lampton, L.M.; Miller, D.P.; Tarka, E.F.; Everett, E.D. Endobronchial pH. Relevance of aminoglycoside activity in gram-negative bacillary pneumonia. Am. Rev. Respir. Dis. 1983, 127, 39–41. [Google Scholar] [CrossRef]
- Karnad, D.R.; Mhaisekar, D.G.; Moralwar, K.V. Respiratory mucus pH in tracheostomized intensive care unit patients: Effects of colonization and pneumonia. Crit. Care Med. 1990, 18, 699–701. [Google Scholar] [CrossRef] [PubMed]
- Lozo Vukovac, E.; Mise, K.; Gudelj, I.; Peric, I.; Duplancic, D.; Vukovic, I.; Vucinovic, Z.; Lozo, M. Bronchoalveolar pH and inflammatory biomarkers in patients with acute exacerbation of chronic obstructive pulmonary disease. J. Int. Med. Res. 2019, 47, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.H.; Clark, N.; Brennan, A.L.; Fisher, D.A.; Gyi, K.M.; Hodson, M.E.; Philips, B.J.; Baines, D.L.; Wood, D.M. Hyperglycemia and cystic fibrosis alter respiratory fluid glucose concentrations estimated by breath condensate analysis. J. Appl. Physiol. 2007, 102, 1969–1975. [Google Scholar] [CrossRef] [PubMed]
- Philips, B.J.; Meguer, J.X.; Redman, J.; Baker, E.H. Factors determining the appearance of glucose in upper and lower respiratory tract secretions. Intensive Care Med. 2003, 29, 2204–2210. [Google Scholar] [CrossRef] [PubMed]
- Philips, B.J.; Redman, J.; Brennan, A.; Wood, D.; Holliman, R.; Baines, D.; Baker, E.H. Glucose in bronchial aspirates increases the risk of respiratory MRSA in intubated patients. Thorax 2005, 60, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Barth, A.L.; Pitt, T.L. The high amino-acid content of sputum from cystic fibrosis patients promotes growth of auxotrophic Pseudomonas aeruginosa. J. Med. Microbiol. 1996, 45, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.R.; Ray, A.; Hodson, M.E.; Pitt, T.L. Increased sputum amino acid concentrations and auxotrophy of Pseudomonas aeruginosa in severe cystic fibrosis lung disease. Thorax 2000, 55, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Sagel, S.D.; Sontag, M.K.; Accurso, F.J. Relationship Between Antimicrobial Proteins and AirwayInflammation and Infection in Cystic Fibrosis. Pediatr. Pulmonol. 2009, 44, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.W.; Lam, Q.T.; Schneider, H.; Walters, E.H. Airway iron and iron-regulatory cytokines in cystic fibrosis. Eur. Respir. J. 2004, 24, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Grasemann, H.; Shehnaz, D.; Enomoto, M.; Leadley, M.; Belik, J.; Ratjen, F. L-ornithine derived polyamines in cystic fibrosis airways. PLoS ONE 2012, 7, e46618. [Google Scholar] [CrossRef]
- Sagel, S.D.; Kapsner, R.; Osberg, I.; Sontag, M.K.; Accurso, F.J. Airway inflammation in children with cystic fibrosis and healthy children assessed by sputum induction. Am. J. Respir. Crit. Care Med. 2001, 164, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Olivença, D.V.; Fonseca, L.L.; Voit, E.O.; Pinto, F.R. Thickness of the airway surface liquid layer in the lung is affected in cystic fibrosis by compromised synergistic regulation of the ENaC ion channel. J. R. Soc. Interface 2019, 16, 20190187. [Google Scholar] [CrossRef] [PubMed]
- Berkebile, A.R.; McCray, P.B., Jr. Effects of airway surface liquid pH on host defense in cystic fibrosis. Int. J. Biochem. Cell Biol. 2014, 52, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.S.; Coakley, R.; Lau, G.W.; Lymar, S.V.; Gaston, B.; Karabulut, A.C.; Hennigan, R.F.; Hwang, S.H.; Buettner, G.; Schurr, M.J.; et al. Anaerobic killing of mucoid Pseudomonas aeruginosa by acidified nitrite derivatives under cystic fibrosis airway conditions. J. Clin. Investig. 2006, 116, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Clary-Meinesz, C.; Mouroux, J.; Cosson, J.; Huitorel, P.; Blaive, B. Influence of external pH on ciliary beat frequency in human bronchi and bronchioles. Eur. Respir. J. 1998, 11, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Simonin, J.; Bille, E.; Crambert, G.; Noel, S.; Dreano, E.; Edwards, A.; Hatton, A.; Pranke, I.; Villeret, B.; Cottart, C.H.; et al. Airway surface liquid acidification initiates host defense abnormalities in Cystic Fibrosis. Sci. Rep. 2019, 9, 6516. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, Y.Z.; Shi, Y.H.; Yang, L.; Chen, X.Y.; Wang, L.W.; Qu, J.M.; Weng, D.; Wang, X.J.; Liu, H.P.; et al. Airway acidification impaired host defense against Pseudomonas aeruginosa infection by promoting type 1 interferon beta response. Emerg. Microbes Infect. 2022, 11, 2132–2146. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, F.; Koutsokera, A.; Bernasconi, E.; Junier, P.; von Garnier, C.; Ubags, N. Recent Advances in Fungal Infections: From Lung Ecology to Therapeutic Strategies With a Focus on Aspergillus spp. Front. Med. 2022, 9, 832510. [Google Scholar] [CrossRef] [PubMed]
- Joris, L.; Dab, I.; Quinton, P.M. Elemental composition of human airway surface fluid in healthy and diseased airways. Am. Rev. Respir. Dis. 1993, 148, 1633–1637. [Google Scholar] [CrossRef]
- Goldman, M.J.; Anderson, G.M.; Stolzenberg, E.D.; Kari, U.P.; Zasloff, M.; Wilson, J.M. Human beta-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cystic fibrosis. Cell 1997, 88, 553–560. [Google Scholar] [CrossRef]
- Garnett, J.P.; Nguyen, T.T.; Moffatt, J.D.; Pelham, E.R.; Kalsi, K.K.; Baker, E.H.; Baines, D.L. Pro-inflammatory mediators disrupt glucose homeostasis in airway surface liquid. J. Immunol. 2012, 189, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.H.; Janaway, C.H.; Philips, B.J.; Brennan, A.L.; Baines, D.L.; Wood, D.M.; Jones, P.W. Hyperglycaemia is associated with poor outcomes in patients admitted to hospital with acute exacerbations of chronic obstructive pulmonary disease. Thorax 2006, 61, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Mirabella, S.; Gomez-Paz, S.; Lam, E.; Gonzalez-Mosquera, L.; Fogel, J.; Rubinstein, S. Glucose dysregulation and its association with COVID-19 mortality and hospital length of stay. Diabetes Metab. Syndr. 2022, 16, 102439. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shen, M.; Tao, Y.; Fairley, C.K.; Zhong, Q.; Li, Z.; Chen, H.; Ong, J.J.; Zhang, D.; Zhang, K.; et al. Elevated glucose level leads to rapid COVID-19 progression and high fatality. BMC Pulm. Med. 2021, 21, 64. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk Factors Associated With Acute Respiratory Distress Syndrome and Death in Patients With Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.H.; Baines, D.L. Airway Glucose Homeostasis: A New Target in the Prevention and Treatment of Pulmonary Infection. Chest 2018, 153, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.K.; Hui, K.; Farne, H.; Garnett, J.P.; Baines, D.L.; Moore, L.S.; Holmes, A.H.; Filloux, A.; Tregoning, J.S. Increased airway glucose increases airway bacterial load in hyperglycaemia. Sci. Rep. 2016, 6, 27636. [Google Scholar] [CrossRef] [PubMed]
- McAlister, F.A.; Majumdar, S.R.; Blitz, S.; Rowe, B.H.; Romney, J.; Marrie, T.J. The relation between hyperglycemia and outcomes in 2471 patients admitted to the hospital with community-acquired pneumonia. Diabetes Care 2005, 28, 810–815. [Google Scholar] [CrossRef]
- Brennan, A.L.; Gyi, K.M.; Wood, D.M.; Johnson, J.; Holliman, R.; Baines, D.L.; Philips, B.J.; Geddes, D.M.; Hodson, M.E.; Baker, E.H. Airway glucose concentrations and effect on growth of respiratory pathogens in cystic fibrosis. J. Cyst. Fibros. 2007, 6, 101–109. [Google Scholar] [CrossRef]
- Garnett, J.P.; Braun, D.; McCarthy, A.J.; Farrant, M.R.; Baker, E.H.; Lindsay, J.A.; Baines, D.L. Fructose transport-deficient Staphylococcus aureus reveals important role of epithelial glucose transporters in limiting sugar-driven bacterial growth in airway surface liquid. Cell. Mol. Life. Sci. 2014, 71, 4665–4673. [Google Scholar] [CrossRef] [PubMed]
- Dennesen, P.; Veerman, E.; van Nieuw Amerongen, A.; Jacobs, J.; Kessels, A.; van der Keybus, P.; Ramsay, G.; van der Ven, A. High levels of sulfated mucins in bronchoalveolar lavage fluid of ICU patients with ventilator-associated pneumonia. Intensive Care Med. 2003, 29, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Davril, M.; Degroote, S.; Humbert, P.; Galabert, C.; Dumur, V.; Lafitte, J.; Lamblin, G.; Roussel, P. The sialylation of bronchial mucins secreted by patients suffering from cystic fibrosis or from chronic bronchitis is related to the severity of airway infection. Glycobiology 1999, 9, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Aprianto, R.; Slager, J.; Holsappel, S.; Veening, J.W. High-resolution analysis of the pneumococcal transcriptome under a wide range of infection-relevant conditions. Nucleic Acids Res. 2018, 46, 9990–10006. [Google Scholar] [CrossRef] [PubMed]
- Blanchette, K.A.; Shenoy, A.T.; Milner, J., 2nd; Gilley, R.P.; McClure, E.; Hinojosa, C.A.; Kumar, N.; Daugherty, S.C.; Tallon, L.J.; Ott, S.; et al. Neuraminidase A-Exposed Galactose Promotes Streptococcus pneumoniae Biofilm Formation during Colonization. Infect. Immun. 2016, 84, 2922–2932. [Google Scholar] [CrossRef] [PubMed]
- Korgaonkar, A.K.; Whiteley, M. Pseudomonas aeruginosa enhances production of an antimicrobial in response to N-acetylglucosamine and peptidoglycan. J. Bacteriol. 2011, 193, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Robbe-Masselot, C.; Fu, X.; Leonard, R.; Marsac, B.; Dauriat, C.J.G.; Lepissier, A.; Rytter, H.; Ramond, E.; Dupuis, M.; et al. Airway environment drives the selection of quorum sensing mutants and promote Staphylococcus aureus chronic lifestyle. Nat. Commun. 2023, 14, 8135. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.K.; Villarreal, A.R.; Ahmad, M.M.; Itabiyi, A.; Feddema, E.; Boyer, H.C.; Hunter, R.C. Anaerobic Microbiota Derived from the Upper Airways Impact Staphylococcus aureus Physiology. Infect. Immun. 2021, 89, e0015321. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Khatua, B.; Mandal, C. Sialic Acid-Siglec-E Interactions During Pseudomonas aeruginosa Infection of Macrophages Interferes With Phagosome Maturation by Altering Intracellular Calcium Concentrations. Front. Immunol. 2020, 11, 332. [Google Scholar] [CrossRef]
- Khatua, B.; Bhattacharya, K.; Mandal, C. Sialoglycoproteins adsorbed by Pseudomonas aeruginosa facilitate their survival by impeding neutrophil extracellular trap through siglec-9. J. Leukoc. Biol. 2012, 91, 641–655. [Google Scholar] [CrossRef]
- Jackson, M.D.; Wong, S.M.; Akerley, B.J. Sialic Acid Protects Nontypeable Haemophilus influenzae from Natural IgM and Promotes Survival in Murine Respiratory Tract. Infect. Immun. 2021, 89, e00676-20. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.M.; Jackson, M.D.; Akerley, B.J. Suppression of Alternative Lipooligosaccharide Glycosyltransferase Activity by UDP-Galactose Epimerase Enhances Murine Lung Infection and Evasion of Serum IgM. Front. Cell. Infect. Microbiol. 2019, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.C.; Nickola, T.J.; Voynow, J.A. Airway mucus obstruction: Mucin glycoproteins, MUC gene regulation and goblet cell hyperplasia. Am. J. Respir. Cell Mol. Biol. 2001, 25, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Green, A.E.; Pottenger, S.; Monshi, M.S.; Barton, T.E.; Phelan, M.; Neill, D.R. Airway metabolic profiling during Streptococcus pneumoniae infection identifies branched chain amino acids as signatures of upper airway colonisation. PLoS Pathog. 2023, 19, e1011630. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F. Carbon catabolite repression in Pseudomonas: Optimizing metabolic versatility and interactions with the environment. FEMS Microbiol. Rev. 2010, 34, 658–684. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, R.; Johansen, H.K.; Molin, S. Adapting to the Airways: Metabolic Requirements of Pseudomonas aeruginosa during the Infection of Cystic Fibrosis Patients. Metabolites 2019, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, R.; Johansen, H.K.; Molin, S. Convergent Metabolic Specialization through Distinct Evolutionary Paths in Pseudomonas aeruginosa. mBio 2018, 9, e00269-18. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Palmer, L.D. Acinetobacter Metabolism in Infection and Antimicrobial Resistance. Infect. Immun. 2023, 91, e0043322. [Google Scholar] [CrossRef] [PubMed]
- Hallman, M.; Merritt, T.A.; Akino, T.; Bry, K. Surfactant protein A, phosphatidylcholine, and surfactant inhibitors in epithelial lining fluid. Correlation with surface activity, severity of respiratory distress syndrome, and outcome in small premature infants. Am. Rev. Respir. Dis. 1991, 144, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.J.; Krasnodembskaya, A.D. Therapeutic targeting of metabolic alterations in acute respiratory distress syndrome. Eur. Respir. Rev. 2020, 29, 200114. [Google Scholar] [CrossRef]
- Aiyer, A.; Manos, J. The Use of Artificial Sputum Media to Enhance Investigation and Subsequent Treatment of Cystic Fibrosis Bacterial Infections. Microorganisms 2022, 10, 1269. [Google Scholar] [CrossRef] [PubMed]
- Schwab, U.; Abdullah, L.H.; Perlmutt, O.S.; Albert, D.; Davis, C.W.; Arnold, R.R.; Yankaskas, J.R.; Gilligan, P.; Neubauer, H.; Randell, S.H.; et al. Localization of Burkholderia cepacia complex bacteria in cystic fibrosis lungs and interactions with Pseudomonas aeruginosa in hypoxic mucus. Infect. Immun. 2014, 82, 4729–4745. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, A.; Mason, M.; Foster, B.J.; Kulkarni, S.; Barlas, A.; Farooq, A.M.; Patak, P.; Liaqat, H.; Basso, R.G.; Zaman, M.S.; et al. Evidence-Based Mechanical Ventilatory Strategies in ARDS. J. Clin. Med. 2022, 11, 319. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A. Lipid mediators in the neural cell nucleus: Their metabolism, signaling, and association with neurological disorders. Neuroscientist 2009, 15, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Zehethofer, N.; Bermbach, S.; Hagner, S.; Garn, H.; Muller, J.; Goldmann, T.; Lindner, B.; Schwudke, D.; Konig, P. Lipid Analysis of Airway Epithelial Cells for Studying Respiratory Diseases. Chromatographia 2015, 78, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.; Lynn, W.S. Lipid composition of sputum from patients with asthma and patients with cystic fibrosis. Inflammation 1978, 3, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Sohlenkamp, C.; Lopez-Lara, I.M.; Geiger, O. Biosynthesis of phosphatidylcholine in bacteria. Prog. Lipid Res. 2003, 42, 115–162. [Google Scholar] [CrossRef] [PubMed]
- Cundell, D.R.; Gerard, N.P.; Gerard, C.; Idanpaan-Heikkila, I.; Tuomanen, E.I. Streptococcus pneumoniae anchor to activated human cells by the receptor for platelet-activating factor. Nature 1995, 377, 435–438. [Google Scholar] [CrossRef]
- Weiser, J.N.; Pan, N.; McGowan, K.L.; Musher, D.; Martin, A.; Richards, J. Phosphorylcholine on the lipopolysaccharide of Haemophilus influenzae contributes to persistence in the respiratory tract and sensitivity to serum killing mediated by C-reactive protein. J. Exp. Med. 1998, 187, 631–640. [Google Scholar] [CrossRef]
- Sun, Z.; Kang, Y.; Norris, M.H.; Troyer, R.M.; Son, M.S.; Schweizer, H.P.; Dow, S.W.; Hoang, T.T. Blocking Phosphatidylcholine Utilization in Pseudomonas aeruginosa, via Mutagenesis of Fatty Acid, Glycerol and Choline Degradation Pathways, Confirms the Importance of This Nutrient Source In Vivo. PLoS ONE 2014, 9, e103778. [Google Scholar] [CrossRef]
- Willsey, G.G.; Ventrone, S.; Schutz, K.C.; Wallace, A.M.; Ribis, J.W.; Suratt, B.T.; Wargo, M.J. Pulmonary Surfactant Promotes Virulence Gene Expression and Biofilm Formation in Klebsiella pneumoniae. Infect. Immun. 2018, 86, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, X.; Wang, S.; Zhang, M.; Yao, W.; Yuan, Y. Determination of related substances in egg yolk lecithin by HPLC-CAD and characterization of its profiling by HPLC-Q-TOF-MS. J. Pharm. Biomed. Anal. 2022, 221, 115079. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Antimicrobial polypeptides in host defense of the respiratory tract. J. Clin. Investig. 2002, 109, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Feingold, D.S.; Goldman, J.N.; Kuritz, H.M. Locus of the action of serum and the role of lysozyme in the serum bactericidal reaction. J. Bacteriol. 1968, 96, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.R.; Cole, M.F.; McGhee, J.R. A bactericidal effect for human lactoferrin. Science 1977, 197, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Drago-Serrano, M.E.; de la Garza-Amaya, M.; Luna, J.S.; Campos-Rodríguez, R. Lactoferrin-lipopolysaccharide (LPS) binding as key to antibacterial and antiendotoxic effects. Int. Immunopharmacol. 2012, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.S.; Morris, A.C.; Kefala, K.; O’Kane, C.M.; Moore, N.R.; Booth, N.A.; McAuley, D.F.; Dhaliwal, K.; Walsh, T.S.; Haslett, C.; et al. Ventilator-Associated Pneumonia Is Characterized by Excessive Release of Neutrophil Proteases in the Lung. Chest 2012, 142, 1425–1432. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Crapo, J.D. Superoxide dismutases in the lung and human lung diseases. Am. J. Respir. Crit. Care Med. 2003, 167, 1600–1619. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.W.; Carroll, V.; O’May, C.; Champion, A.; Kirov, S.M. Increased airway iron as a potential factor in the persistence of Pseudomonas aeruginosa infection in cystic fibrosis. Eur. Respir. J. 2007, 30, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Anderson, G.J.; Bell, S.C.; Reid, D.W. Elevated metal concentrations in the CF airway correlate with cellular injury and disease severity. J. Cyst. Fibros. 2014, 13, 289–295. [Google Scholar] [CrossRef]
- Sunder-Plassmann, G.; Patruta, S.I.; Horl, W.H. Pathobiology of the role of iron in infection. Am. J. Kidney Dis. 1999, 34, S25–S29. [Google Scholar] [CrossRef] [PubMed]
- Bussiere, F.I.; Gueux, E.; Rock, E.; Girardeau, J.P.; Tridon, A.; Mazur, A.; Rayssiguier, Y. Increased phagocytosis and production of reactive oxygen species by neutrophils during magnesium deficiency in rats and inhibition by high magnesium concentration. Br. J. Nutr. 2002, 87, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.D.; Duncan, A.; Noble, D.; Imrie, M.; O’Reilly, D.S.; Innes, J.A.; Porteous, D.J.; Greening, A.P.; Boyd, A.C. Sputum trace metals are biomarkers of inflammatory and suppurative lung disease. Chest 2010, 137, 635–641. [Google Scholar] [CrossRef]
- Stites, S.W.; Walters, B.; O’Brien-Ladner, A.R.; Bailey, K.; Wesselius, L.J. Increased iron and ferritin content of sputum from patients with cystic fibrosis or chronic bronchitis. Chest 1998, 114, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Stites, S.W.; Plautz, M.W.; Bailey, K.; O’Brien-Ladner, A.R.; Wesselius, L.J. Increased concentrations of iron and isoferritins in the lower respiratory tract of patients with stable cystic fibrosis. Am. J. Respir. Crit. Care Med. 1999, 160, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef]
- Chou, H.T.; Li, J.; Peng, Y.; Lu, C. Molecular characterization of PauR and its role in control of putrescine and cadaverine catabolism through the γ-glutamylation pathway in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2013, 195, 3906–3913. [Google Scholar] [CrossRef] [PubMed]
- Twomey, K.B.; Alston, M.; An, S.Q.; O’Connell, O.J.; McCarthy, Y.; Swarbreck, D.; Febrer, M.; Dow, J.M.; Plant, B.J.; Ryan, R.P. Microbiota and metabolite profiling reveal specific alterations in bacterial community structure and environment in the cystic fibrosis airway during exacerbation. PLoS ONE 2013, 8, e82432. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.V.; McGilvrey, M.I.; Hu, C.K.; Garcia-Mansfield, K.; Lewandoski, K.; Eftekhari, Z.; Yuan, Y.C.; Zenhausern, F.; Menashi, E.; Pirrotte, P. Molecular Profiling of Innate Immune Response Mechanisms in Ventilator-associated Pneumonia. Mol. Cell. Proteom. 2020, 19, 1688–1705. [Google Scholar] [CrossRef] [PubMed]
- Holz, O.; DeLuca, D.S.; Roepcke, S.; Illig, T.; Weinberger, K.M.; Schudt, C.; Hohlfeld, J.M. Smokers with COPD Show a Shift in Energy and Nitrogen Metabolism at Rest and During Exercise. Int. J. Chron. Obstruct. Pulmon. Dis. 2020, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hasan, C.M.; Pottenger, S.; Green, A.E.; Cox, A.A.; White, J.S.; Jones, T.; Winstanley, C.; Kadioglu, A.; Wright, M.H.; Neill, D.R.; et al. Pseudomonas aeruginosa utilises the host-derived polyamine spermidine to facilitate antimicrobial tolerance. JCI Insight 2022, 7, e158879. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Lu, C.D. Polyamine effects on antibiotic susceptibility in bacteria. Antimicrob. Agents Chemother. 2007, 51, 2070–2077. [Google Scholar] [CrossRef] [PubMed]
- Innes, A.L.; Carrington, S.D.; Thornton, D.J.; Kirkham, S.; Rousseau, K.; Dougherty, R.H.; Raymond, W.W.; Caughey, G.H.; Muller, S.J.; Fahy, J.V. Ex vivo sputum analysis reveals impairment of protease-dependent mucus degradation by plasma proteins in acute asthma. Am. J. Respir. Crit. Care Med. 2009, 180, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Sankar, S.; Yamaguchi, M.; Kawabata, S.; Ponnuraj, K. Streptococcus pneumoniae Surface Adhesin PfbA Exhibits Host Specificity by Binding to Human Serum Albumin but Not Bovine, Rabbit and Porcine Serum Albumins. Protein J. 2020, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Egesten, A.; Frick, I.M.; Mörgelin, M.; Olin, A.I.; Björck, L. Binding of Albumin Promotes Bacterial Survival at the Epithelial Surface. J. Biol. Chem. 2011, 286, 2469–2476. [Google Scholar] [CrossRef]
- Teevan-Hanman, C.; O’Shea, P. Candida albicans exhibit two classes of cell surface binding sites for serum albumin defined by their affinity, abundance and prospective role in interkingdom signalling. PLoS ONE 2021, 16, e0254593. [Google Scholar] [CrossRef]
- Austermeier, S.; Pekmezovic, M.; Porschitz, P.; Lee, S.; Kichik, N.; Moyes, D.L.; Ho, J.; Kotowicz, N.K.; Naglik, J.R.; Hube, B.; et al. Albumin Neutralizes Hydrophobic Toxins and Modulates Candida albicans Pathogenicity. mBio 2021, 12, e0053121. [Google Scholar] [CrossRef] [PubMed]
- Kruczek, C.; Wachtel, M.; Alabady, M.S.; Payton, P.R.; Colmer-Hamood, J.A.; Hamood, A.N. Serum albumin alters the expression of iron-controlled genes in Pseudomonas aeruginosa. Microbiology 2012, 158, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.; Rice, A.; Sutton, B.; Gabrilska, R.; Wessel, A.K.; Whiteley, M.; Rumbaugh, K.P. Albumin Inhibits Pseudomonas aeruginosa Quorum Sensing and Alters Polymicrobial Interactions. Infect. Immun. 2017, 85, e00116-17. [Google Scholar] [CrossRef] [PubMed]
- Park, W.K.; Chung, J.W.; Kim, Y.K.; Chung, S.C.; Kho, H.S. Influences of animal mucins on lysozyme activity in solution and on hydroxyapatite surfaces. Arch. Oral Biol. 2006, 51, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.C.; Neuberger, A.; Wilson, B.M. The dependence of lysozyme activity on pH and ionic strength. Biochim. Biophys. Acta 1969, 178, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Pahud, J.J.; Schellenberg, D.; Monti, J.C.; Scherz, J.C. Lysozyme, an abomasal enzyme in the ruminants. Ann. Rech. Vet. 1983, 14, 493–501. [Google Scholar] [PubMed]
- Chirkov, N.S.; Lin, S.-Y.; Michailov, A.V.; Miller, R.; Noskov, B.A. DNA Penetration into a Lysozyme Layer at the Surface of Aqueous Solutions. Int. J. Mol. Sci. 2022, 23, 12377. [Google Scholar] [CrossRef] [PubMed]
- Bergamo, A.; Sava, G. Lysozyme: A Natural Product with Multiple and Useful Antiviral Properties. Molecules 2024, 29, 652. [Google Scholar] [CrossRef] [PubMed]
- Curry, S.; Brick, P.; Franks, N.P. Fatty acid binding to human serum albumin: New insights from crystallographic studies. Biochim. Biophys. Acta 1999, 1441, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Vita, G.M.; De Simone, G.; De Marinis, E.; Nervi, C.; Ascenzi, P.; di Masi, A. Serum albumin and nucleic acids biodistribution: From molecular aspects to biotechnological applications. IUBMB Life 2022, 74, 866–879. [Google Scholar] [CrossRef]
- Dubeau, S.; Bourassa, P.; Thomas, T.J.; Tajmir-Riahi, H.A. Biogenic and synthetic polyamines bind bovine serum albumin. Biomacromolecules 2010, 11, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Bal, W.; Sokolowska, M.; Kurowska, E.; Faller, P. Binding of transition metal ions to albumin: Sites, affinities and rates. Biochim. Biophys. Acta 2013, 1830, 5444–5455. [Google Scholar] [CrossRef] [PubMed]
- Kakati, N.; Parashar, C.K.; Thakur, S.; Deshmukh, O.S.; Bandyopadhyay, D. Microrheology of Mucin-Albumin Assembly Using Diffusing Wave Spectroscopy. ACS Appl. Bio Mater. 2022, 5, 4118–4127. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.H.; Turner, B.; Bhaskar, K.R.; Lamont, J.T. Lipid binding to gastric mucin: Protective effect against oxygen radicals. Am. J. Physiol. 1990, 259 4 Pt 1, G681–G686. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.E.; Umbreit, J.N.; Moore, E.G. A role for mucin in the absorption of inorganic iron and other metal cations. A study in rats. Gastroenterology 1991, 100, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ghio, A.J.; Soukup, J.M.; Dailey, L.A.; Roggli, V.L. Mucus increases cell iron uptake to impact the release of pro-inflammatory mediators after particle exposure. Sci. Rep. 2023, 13, 3925. [Google Scholar] [CrossRef] [PubMed]
- Casale, A.; De Stefano, C.; Manfredi, G.; Milea, D.; Sammartano, S. Sequestration of Alkyltin(IV) compounds in aqueous solution: Formation, stability, and empirical relationships for the binding of dimethyltin(IV) cation by N- and O-donor ligands. Bioinorg. Chem. Appl. 2009, 2009, 219818. [Google Scholar] [CrossRef] [PubMed]
- Pelta, J.; Livolant, F.; Sikorav, J.L. DNA aggregation induced by polyamines and cobalthexamine. J. Biol. Chem. 1996, 271, 5656–5662. [Google Scholar] [CrossRef]
- Natalini, J.G.; Singh, S.; Segal, L.N. The dynamic lung microbiome in health and disease. Nat. Rev. Microbiol. 2023, 21, 222–235. [Google Scholar] [CrossRef]
- Li, R.; Li, J.; Zhou, X. Lung microbiome: New insights into the pathogenesis of respiratory diseases. Sig. Transduct. Target. Ther. 2024, 9, 19. [Google Scholar] [CrossRef]
- Morrison, A.G.; Sarkar, S.; Umar, S.; Lee, S.T.M.; Thomas, S.M. The Contribution of the Human Oral Microbiome to Oral Disease: A Review. Microorganisms 2023, 11, 318. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, P.A.; Cooper, P.J.; Cox, M.J.; Chico, M.; Arias, C.; Moffatt, M.F.; Cookson, W.O. Upper airways microbiota in antibiotic-naive wheezing and healthy infants from the tropics of rural Ecuador. PLoS ONE 2012, 7, e46803. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Marsland, B.J.; Bunyavanich, S.; O’Mahony, L.; Leung, D.Y.; Muraro, A.; Fleisher, T.A. The microbiome in allergic disease: Current understanding and future opportunities-2017 PRACTALL document of the American Academy of Allergy, Asthma & Immunology and the European Academy of Allergy and Clinical Immunology. J. Allergy Clin. Immunol. 2017, 139, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Lyon, J. The Lung Microbiome: Key to Respiratory Ills? JAMA 2017, 317, 1713–1714. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Sze, M.A.; Dimitriu, P.A.; Hayashi, S.; Elliott, W.M.; McDonough, J.E.; Gosselink, J.V.; Cooper, J.; Sin, D.D.; Mohn, W.W.; Hogg, J.C. The lung tissue microbiome in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2012, 185, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Boutin, S.; Graeber, S.Y.; Stahl, M.; Dittrich, S.A.; Mall, M.A.; Dalpke, A.H. Chronic but not intermittent infection with Pseudomonas aeruginosa is associated with global changes of the lung microbiome in cystic fibrosis. Eur. Respir. J. 2017, 50, 1701086. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.; Park, S.Y.; Kim, Y.; Jeon, J.P.; Lee, J.J.; Hong, J.Y. The Dynamics of Respiratory Microbiota during Mechanical Ventilation in Patients with Pneumonia. J. Clin. Med. 2020, 9, 638. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Craven, V.; Bingle, L.; Cookson, W.; Everard, M.L.; Moffatt, M.F. The impact of persistent bacterial bronchitis on the pulmonary microbiome of children. PLoS ONE 2017, 12, e0190075. [Google Scholar] [CrossRef] [PubMed]
- Madapoosi, S.S.; Cruickshank-Quinn, C.; Opron, K.; Erb-Downward, J.R.; Begley, L.A.; Li, G.; Barjaktarevic, I.; Barr, R.G.; Comellas, A.P.; Couper, D.J.; et al. Lung Microbiota and Metabolites Collectively Associate with Clinical Outcomes in Milder Stage Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2022, 206, 427–439. [Google Scholar] [CrossRef]


{kind=link}
| Component | Average Concentrations | Conditions | Detection Method |
|---|---|---|---|
| Mucin | 2.7 mg/mL [61], 6.5 mg/mL [61], ~5 mg/mL [15], ~20 mg/mL [15]. | Healthy [61], CF exacerbation [61], newly intubated elective laryngoscopy patients [15], ICU patients mechanically ventilated for at least 4 days [15]. | Size exclusion chromatography/differential refractive index [61], enzyme-linked immunosorbent assay of subglottic samples [15]. |
| DNA | 0.96 mg/mL [55], 6.7mg/mL [55], 5.2 mg/mL [55], 20 ng/mL [15,92], 40 ng/mL [15], ~100 ng/mL [92], ~250 ng/mL [92], 0.7 µg/mL [104], 3.2 µg/mL [104], 5.4 µg/mL [104], 2 µg/mL [105], 10 µg/mL [105], 416% more DNA by area in CF sputum compared to asthma and chronic bronchitis sputum [60]. | Healthy [55,92,105], stable CF [55,105], CF exacerbation [55], non-ICU patients [15], ICU patients [15], VAP [92], ARDS [92], VAP and ARDS [92], non-CF patients [104], infants with CF [104], older CF patients [104]. | Microfluorimetry [55], fluorometric assays [15], colorimetric assays [92], Hoechst dye-binding assay [104], Quant-iT PicoGreen assay [105], confocal microscopy [60]. |
| pH | 6.2–7 (nasal) [106], 7.1 (lower airways) [106], 6.78 [107], 7.18 [108], 6.57 [108], 6.97 [109], 6.58 [110], 6.62 [110], 6.72 [111], 6.61 [110], 6.89 [112]. | Non-CF [106,107,108,109,110], CF [106,108], pneumonia [110,111], chronic lung disease [110], acute exacerbation of COPD [112]. | Monocrystalline antimony catheter [106], in-gold combined pH-glass electrode [106], fluorescent indicators on freshly excised human bronchi [107,109], fluorescent indicators on nasal biopsies [108], pH electrode [110], pH strips [112]. |
| Glucose | 0.4 mM [113], 1 mM [114], 4 mM [114], 1.2 mM [113], 2 mM [113], 3.5 mM [115]. | Healthy [113], viral infection [114], hyperglycaemic diabetes [114], CF [113], CF and diabetes [113], mechanically ventilated patients [115]. | High-performance anion-exchange chromatography with pulsed amperometric detection [113], glucose oxidase sticks [115]. |
| Amino acids | 2.52 mg/mL; total [116], 5.7 mg/mL; total [116], 12.3 mM; total [117], 18.2 mM; total [117], 0.42 nmol/mg; alanine [47], 2.2 nmol/mg; asparagine [47], 0.42 nmol/mg; glutamine [47], 1.06 nmol/mg; glycine [47], 0.43 nmol/mg; lysine [47], 0.13 nmol/mg; valine [47]. | Healthy [116], CF [116,117], CF exacerbation [117], healthy tissue from lobectomies of lung cancer patients [47]. | Thin layer chromatography [116], high-performance liquid chromatography [117], nuclear magnetic resonance [47]. |
| Lysozyme | 3.9 µg/mL [118], 9.1 µg/mL [118]. | Culture-negative CF patients [118], culture-positive CF patients [118]. | Lysozyme activity assay [118]. |
| Lactoferrin | 5 µg/mL [61], 9 µg/mL [61], 3.0 µg/mL [118], 22.3 µg/mL [118]. | Non-CF [61], CF [61], culture-negative CF patients [118], culture-positive CF patients [118]. | Immunologic techniques [61], enzyme-linked immunosorbent assay [118]. |
| Ferritin | 0.2 µg/mL [119], 2.4 µg/mL [119], 3.6 µg/mL [119], 0.6 µg/mL [119]. | Healthy [119], CF [119], CF exacerbation [119], COPD [119]. | Microparticle enzyme immunoassay [119]. |
| Putrescine | 11.91 µmol/L [120], 6.18 µmol/L [120], 96.02 µmol/L [120], 20.59 µmol/L [120]. | Healthy [120], CF stable [120], CF exacerbation pre-antibiotic treatment [120], CF exacerbation post-antibiotic treatment [120]. | High-performance liquid chromatography [120]. |
| Spermine | 0.22 µmol/L [120], 1.71 µmol/L [120], 7.32 µmol/L [120], 1.35 µmol/L [120]. | Healthy [120], CF stable [120], CF exacerbation pre-antibiotic treatment [120], CF exacerbation post-antibiotic treatment [120]. | High-performance liquid chromatography [120]. |
| Spermidine | 0.88 µmol/L [120], 1.62 µmol/L [120], 0.78 µmol/L [120], 0.62 µmol/L [120]. | Healthy [120], CF stable [120], CF exacerbation pre-antibiotic treatment [120], CF exacerbation post-antibiotic treatment [120]. | High-performance liquid chromatography [120]. |
| Serum albumin | 0.1 dg/L [119], 0.4 dg/L [119], 0.7 dg/L [119], 0.2 dg/mL [119]. 127.4 µg/mL [121], 244.4 µg/mL [121]. | Healthy [119,121], CF [119,121], CF exacerbation [119], COPD [119]. | Rate immunophelometry [119], competitive radioimmunoassay [121]. |
| Metal | Healthy | Asthma | COPD | CF | Bronchiectasis | Smoker |
|---|---|---|---|---|---|---|
| Zn | 15.35 (10.4–25.6) a 179 (103–597) b 40.45 (20.99) c | 12.7 (7.2–41.4) a | 25.4 (9.8–50.7) a | 135.3 (54.2–209.6) a 1285 (678–1811) b | 111.3 (46.1–150.7) a 537 (401–838) b | 48.16 (35.06) c |
| Fe | 13.5 (8.6–21.5) a 0 (0–37) b 6.38 (9.12) c | 30 (6.9–35.3) a | 21.3 (3.1–35.6) a | 56.9 (24.3–115.3) a 797 (398–1292) b | 54.2 (22.7–91.6) a 1075 (862–1324) b | 23.37 (28.47) c |
| Mn | 0 (0–0.25) a 5 (2–9) b 0.12 (0.16) c | 0.8 (0.2–1.7) a | 0 (0–0.7) a | 0.3 (0.1–0.8) a 6 (4–17) b | 0.6 (0.2–1.3) a 6 (4–10) b | 0.21 (0.24) c |
| Cu | 8.6 (3–16.4) a 106 (55.3–196) b 4.77 (4.9) c | 15.2 (8.6–29.5) a | 15.2 (12.2–22) a | 19.5 (14.5–30.1) a 173 (128–257) b | 15.7 (10.9–33.3) a 226 (130–314) b | 4 (2.26) c |
| Ca | 45,000 (28,000–58,000) b 811.7 (181.61) c | 102,000 (76,000–123,000) b | 124,000 (78,000–156,000) b | 856.8 (183.34) c | ||
| Mg | 4000 (2000–7000) b 389.06 (66.93) c | 30,000 (19,000–44,000) b | 33,000 (27,000–39,000) b | 428.76 (110.42) c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walsh, D.; Bevan, J.; Harrison, F. How Does Airway Surface Liquid Composition Vary in Different Pulmonary Diseases, and How Can We Use This Knowledge to Model Microbial Infections? Microorganisms 2024, 12, 732. https://doi.org/10.3390/microorganisms12040732
Walsh D, Bevan J, Harrison F. How Does Airway Surface Liquid Composition Vary in Different Pulmonary Diseases, and How Can We Use This Knowledge to Model Microbial Infections? Microorganisms. 2024; 12(4):732. https://doi.org/10.3390/microorganisms12040732
Chicago/Turabian StyleWalsh, Dean, Jennifer Bevan, and Freya Harrison. 2024. "How Does Airway Surface Liquid Composition Vary in Different Pulmonary Diseases, and How Can We Use This Knowledge to Model Microbial Infections?" Microorganisms 12, no. 4: 732. https://doi.org/10.3390/microorganisms12040732
APA StyleWalsh, D., Bevan, J., & Harrison, F. (2024). How Does Airway Surface Liquid Composition Vary in Different Pulmonary Diseases, and How Can We Use This Knowledge to Model Microbial Infections? Microorganisms, 12(4), 732. https://doi.org/10.3390/microorganisms12040732