
Diversity and Antibiotic Resistance of Triticale Seed-Borne Bacteria on the Tibetan Plateau


Abstract
1. Introduction
2. Materials and Methods
2.1. Triticale Seeds
2.2. Antibiotics and Media
2.3. Separation, Purification and Preservation of Bacteria from Triticale Seeds
2.4. Extraction and Appraisal of 16S RRNA from Triticale Seedborne Bacteria
2.5. Determination of Bacterium Isolate Motility
2.6. Determination of the Biofilm Formation Ability of Bacteria
2.7. Determination of Antibiotic Resistance of Bacteria
3. Results
3.1. The Characteristics of Triticale Seedborne Bacteria
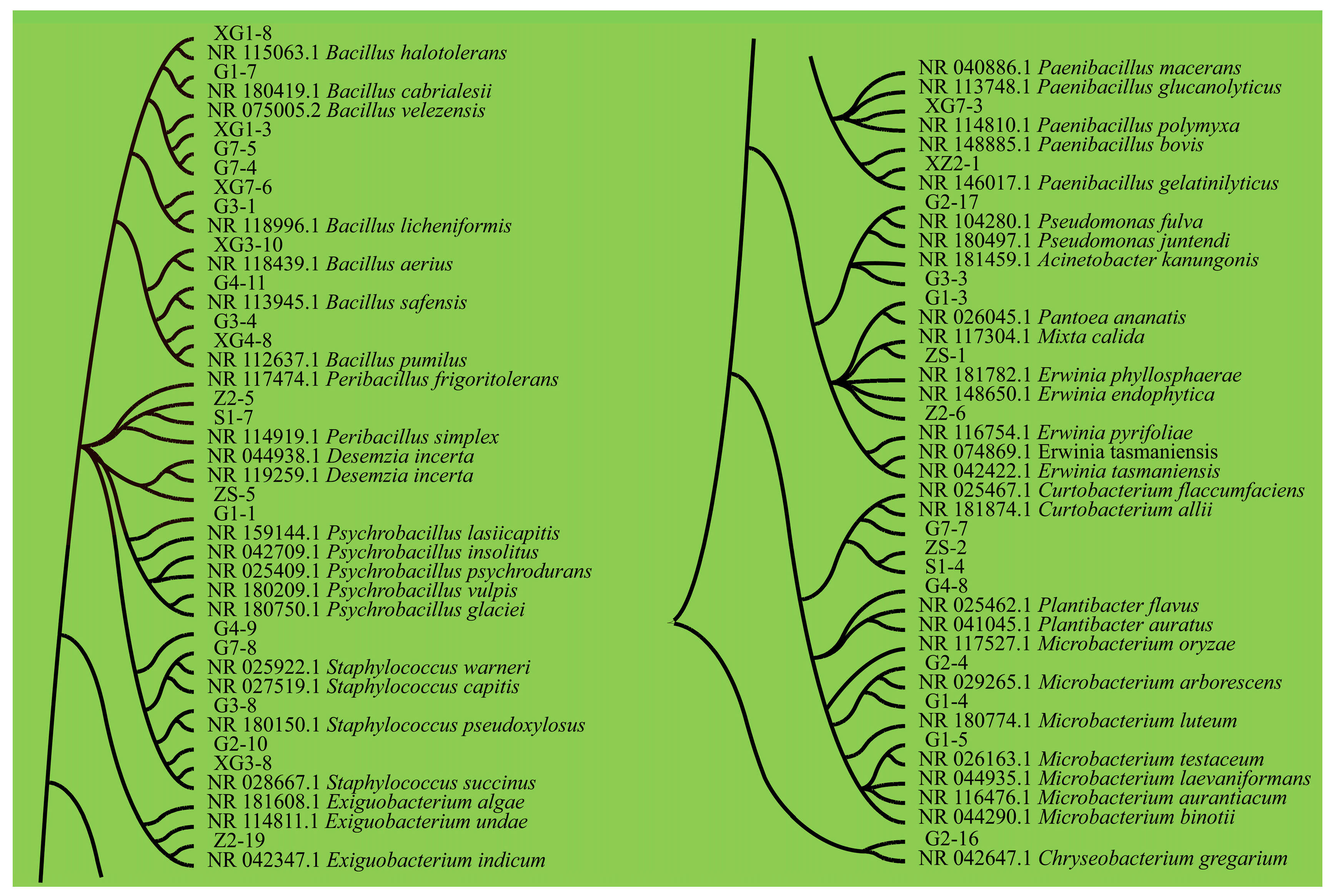
3.1.1. Isolation and Identification
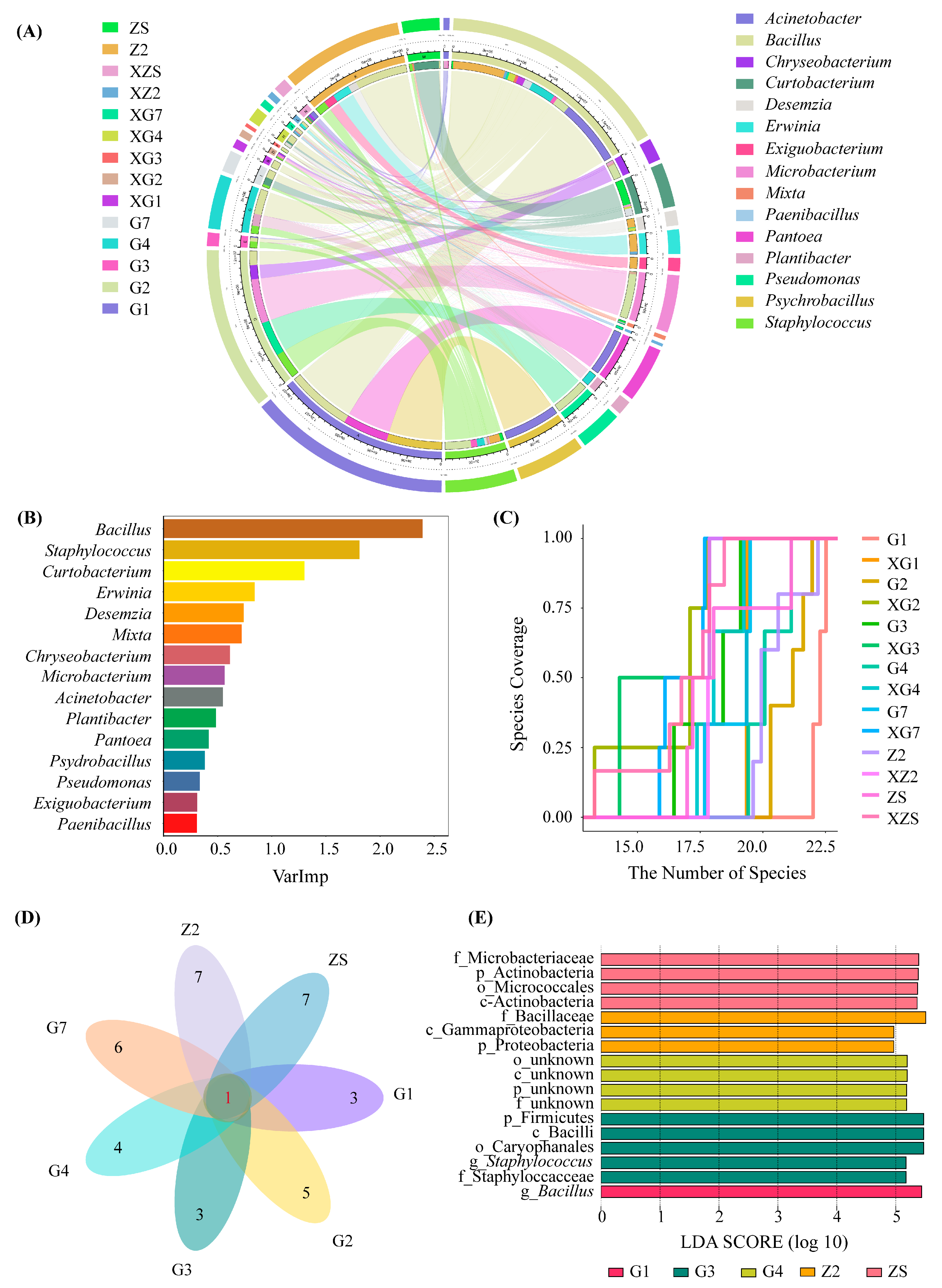
3.1.2. Bacterial Community Abundance Composition Analysis
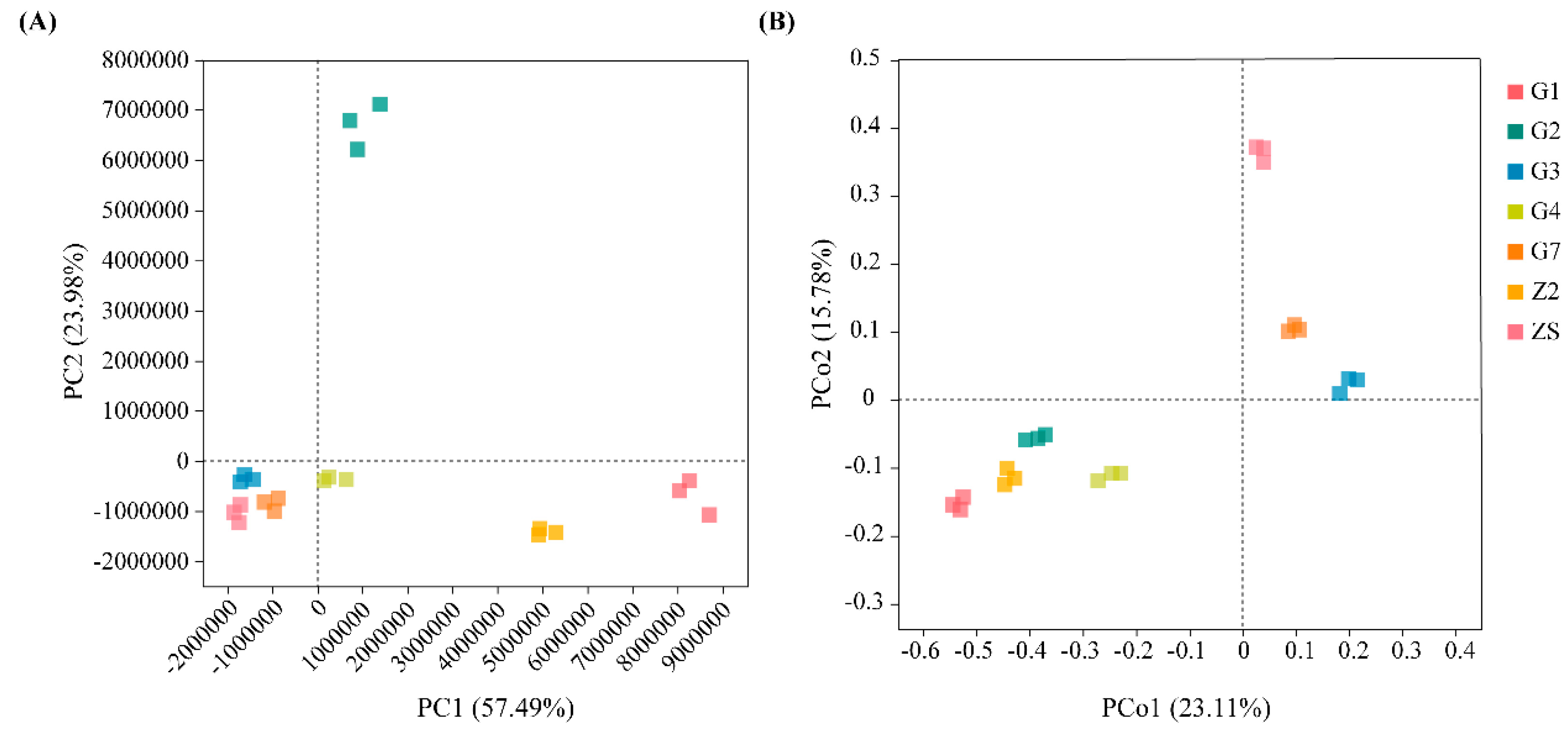
3.1.3. Differences in Culturable Triticale Seedborne Bacterial Communities
3.1.4. Ecological Niche Analysis of Culturable Triticale Seedborne Bacteria
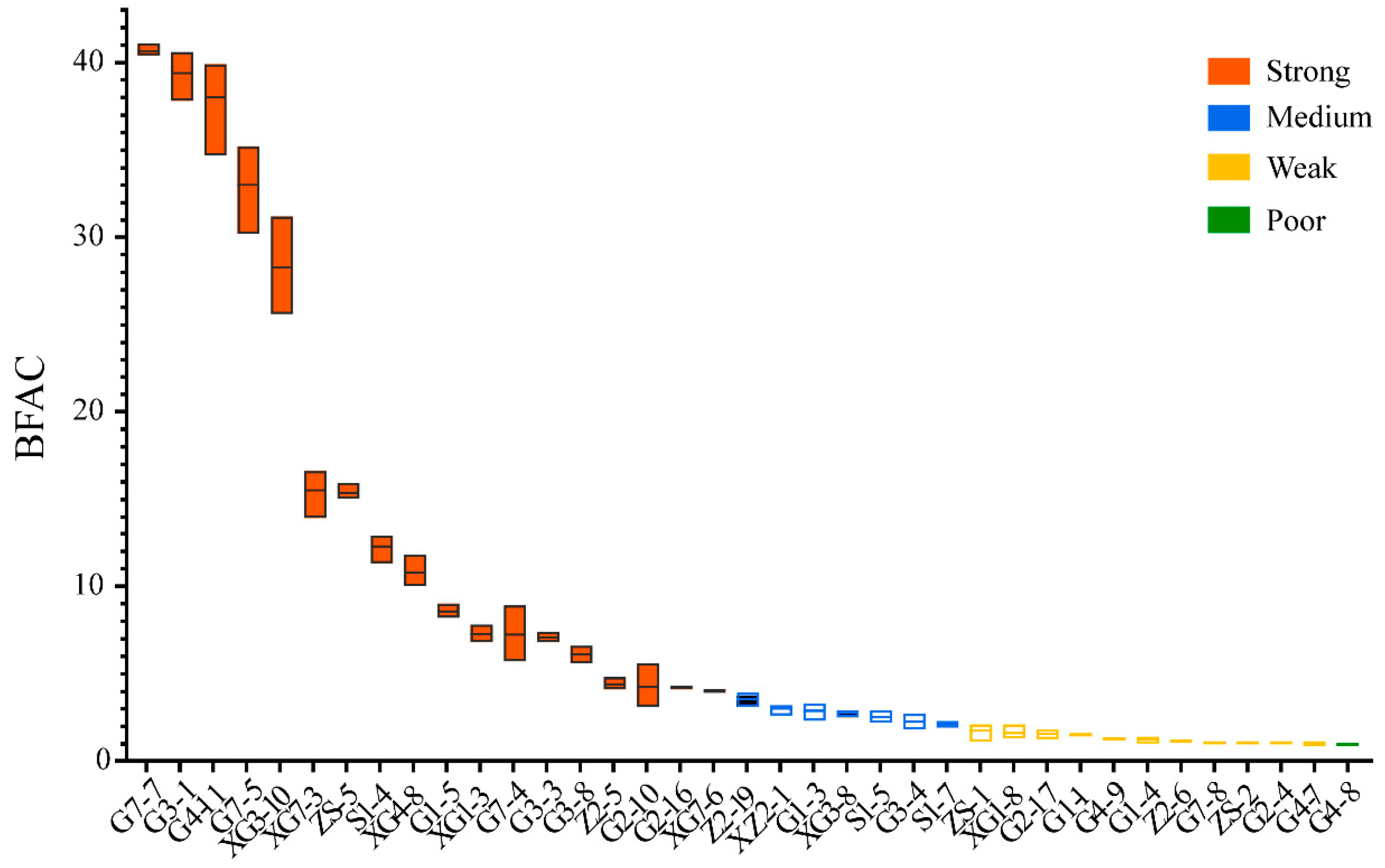
3.2. Biofilm Formation Ability
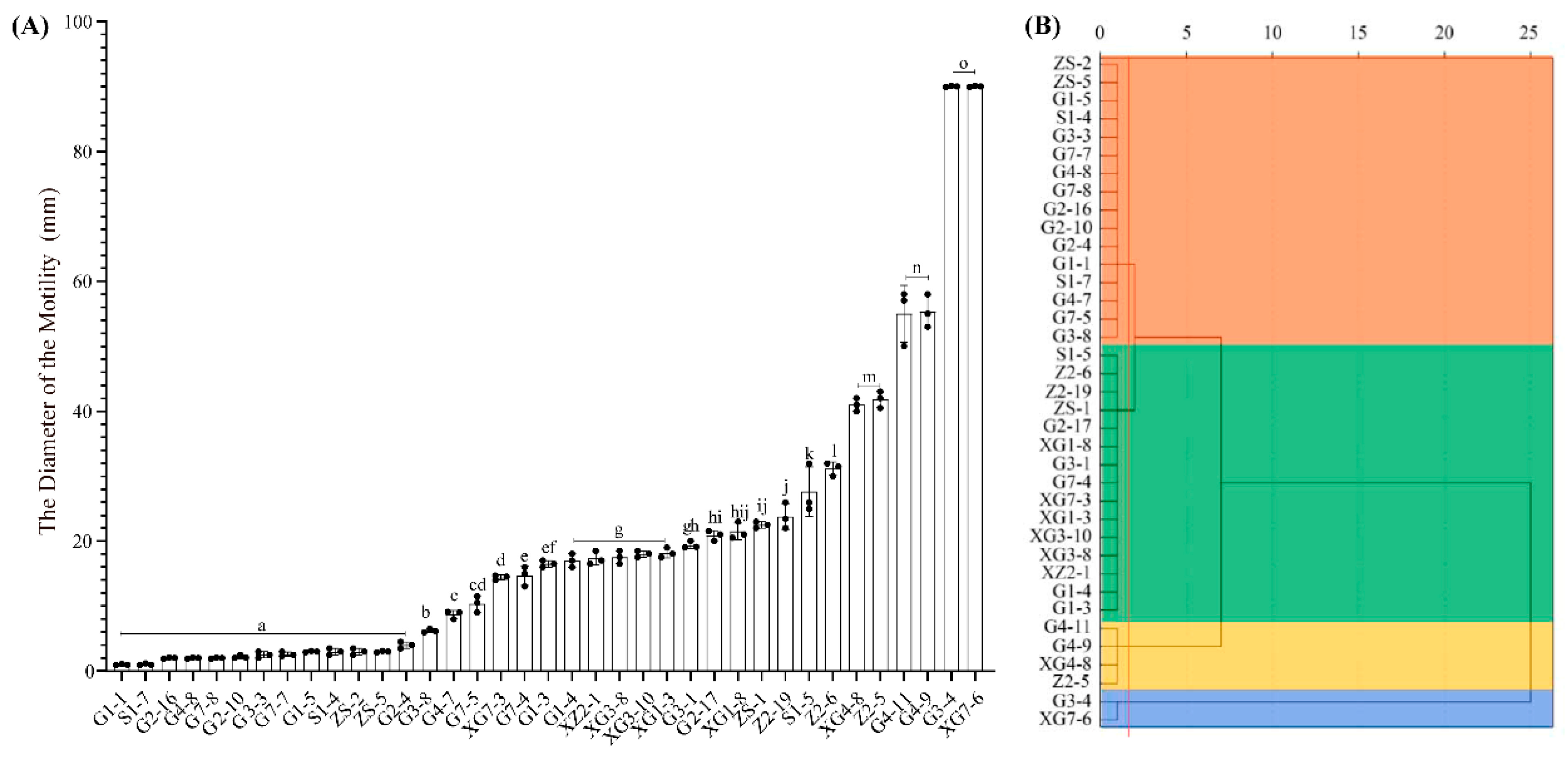
3.3. Motility
3.4. Bacterial Resistance to Antibiotics
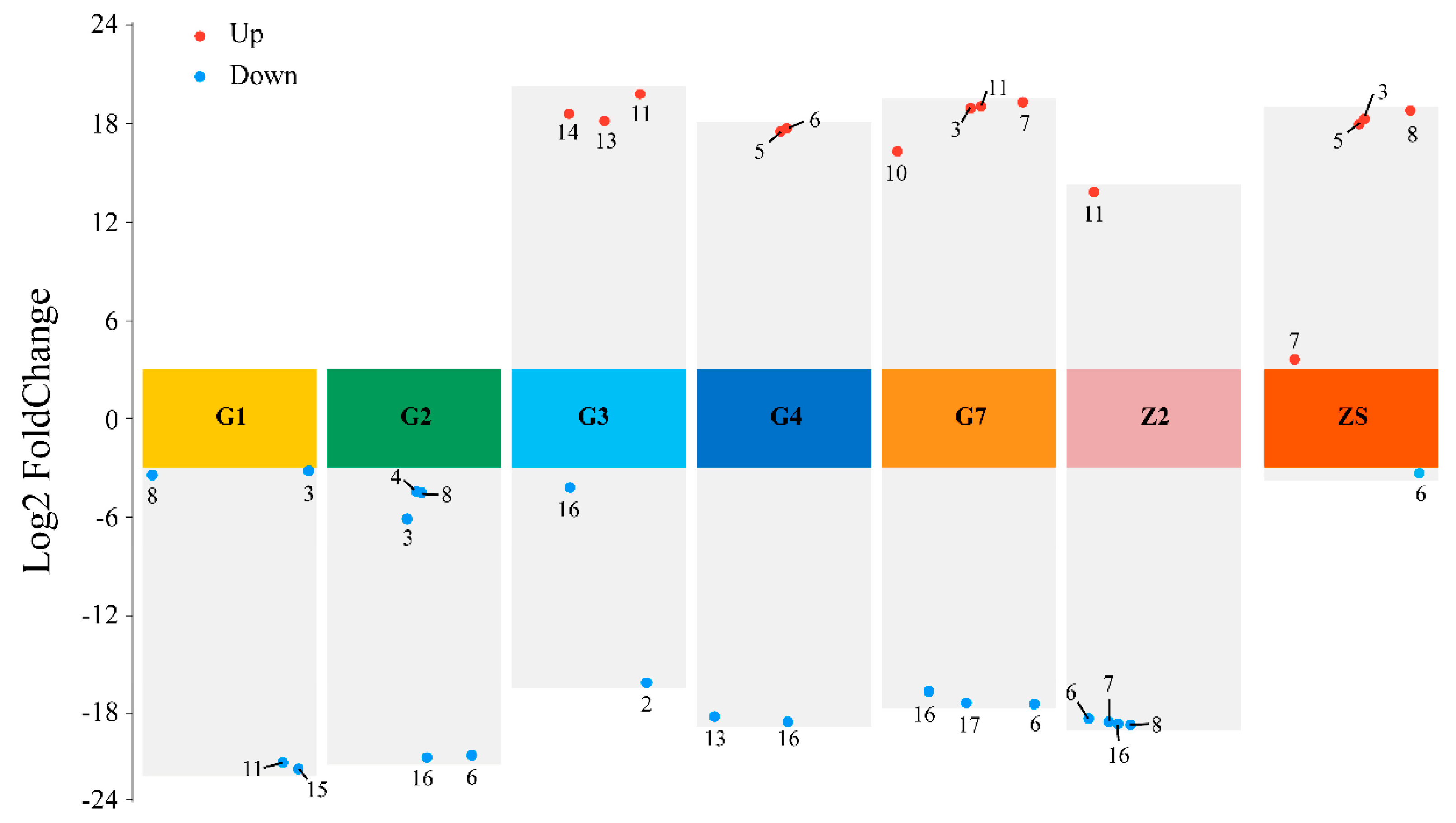
3.4.1. Qualitative Screening
3.4.2. Quantitative Screening
3.4.3. Quantitative Analysis of the IC50
3.4.4. Visualisation of the Resistance Analysis Results
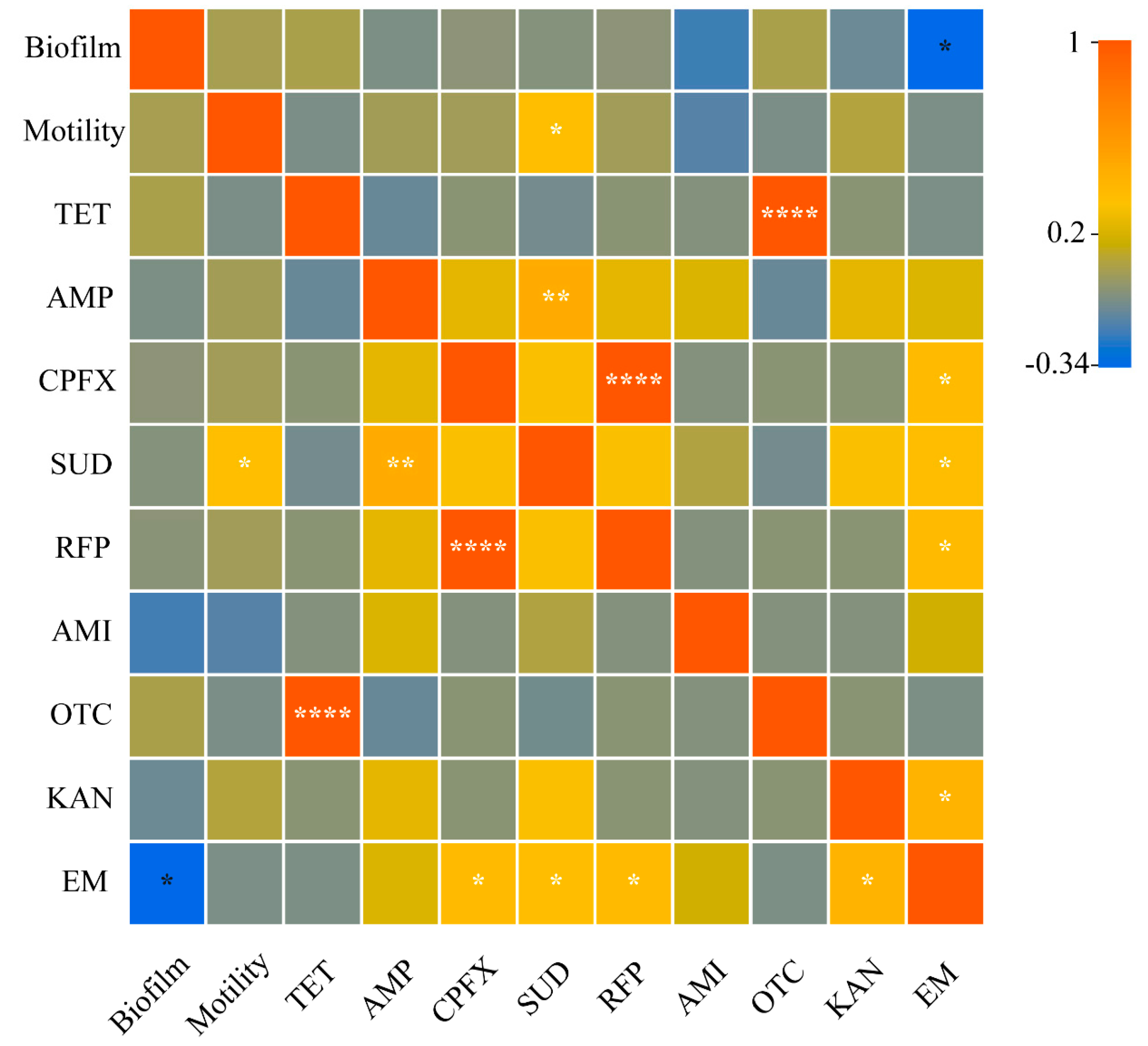
3.5. Correlations between Resistance and Motility and Biofilm Formation Ability
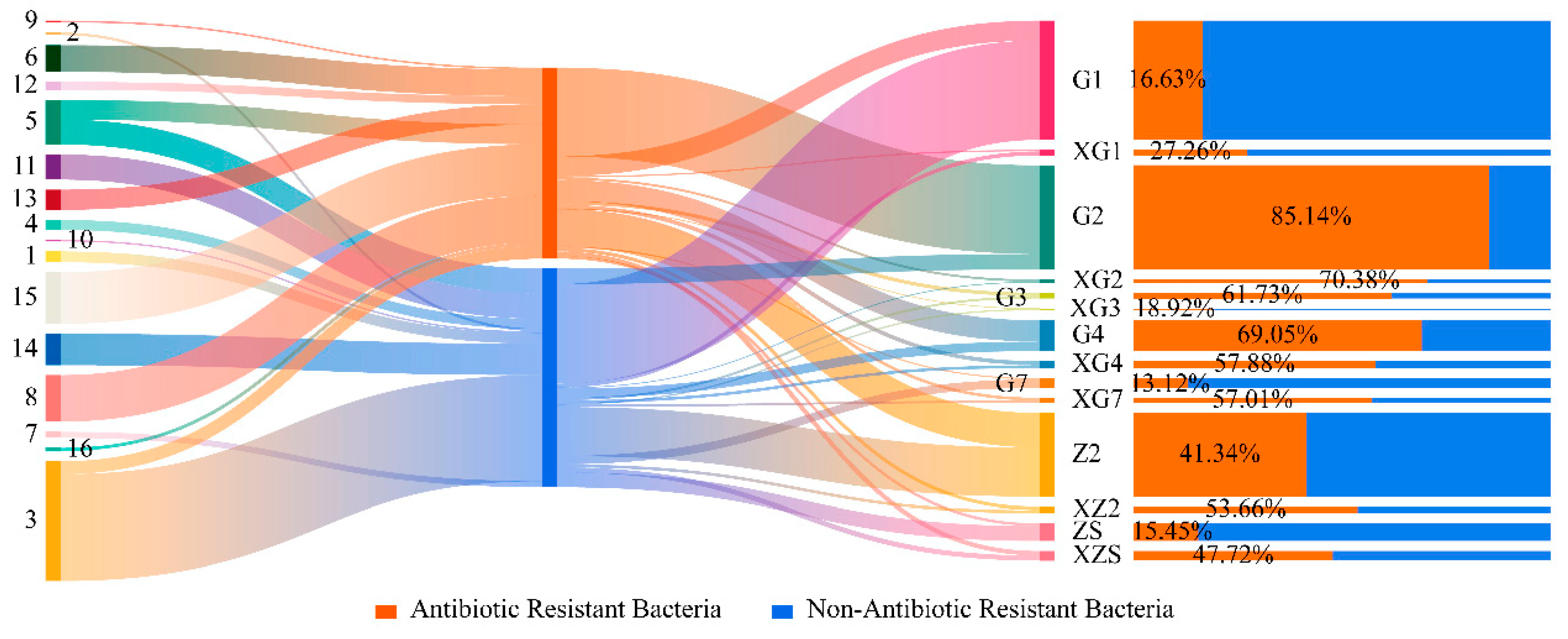
3.6. Analysis of Bacterial Resistance and Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mulchandani, R.; Wang, Y.; Gilbert, M.; Van Boeckel, T.P. Global Trends in Antimicrobial Use in Food-Producing Animals: 2020 to 2030. PLoS Glob. Public Health 2023, 3, e0001305. [Google Scholar] [CrossRef]
- Aarestrup, F.M. The Livestock Reservoir for Antimicrobial Resistance: A Personal View on Changing Patterns of Risks, Effects of Interventions and the Way Forward. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140085. [Google Scholar] [CrossRef]
- Niemi, M.; Backman, J.T.; Fromm, M.F.; Neuvonen, P.J.; Kivistö, K.T. Pharmacokinetic Interactions with Rifampicin: Clinical Relevance. Clin. Pharmacokinet. 2003, 42, 819–850. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The Antimicrobial Resistance Crisis: Causes, Consequences, and Management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; Arena, F.; Pecile, P.; Pollini, S. Update on the Antibiotic Resistance Crisis. Curr. Opin. Pharmacol. 2014, 18, 56–60. [Google Scholar] [CrossRef]
- Lee, C.-R.; Lee, J.H.; Kang, L.-W.; Jeong, B.C.; Lee, S.H. Educational Effectiveness, Target, and Content for Prudent Antibiotic Use. BioMed Res. Int. 2015, 2015, 214021. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in Agriculture and the Risk to Human Health: How Worried Should We Be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, J. Detection and Diversity Evaluation of Tetracycline Resistance Genes in Grassland-Based Production Systems in Colombia, South America. Front. Microbiol. 2011, 2, 15715. [Google Scholar] [CrossRef] [PubMed]
- Jechalke, S.; Kopmann, C.; Rosendahl, I.; Groeneweg, J.; Weichelt, V.; Krögerrecklenfort, E.; Brandes, N.; Nordwig, M.; Ding, G.-C.; Siemens, J.; et al. Increased Abundance and Transferability of Resistance Genes after Field Application of Manure from Sulfadiazine-Treated Pigs. Appl. Environ. Microbiol. 2013, 79, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Nõlvak, H.; Truu, M.; Kanger, K.; Tampere, M.; Espenberg, M.; Loit, E.; Raave, H.; Truu, J. Inorganic and Organic Fertilizers Impact the Abundance and Proportion of Antibiotic Resistance and Integron-Integrase Genes in Agricultural Grassland Soil. Sci. Total Environ. 2016, 562, 678–689. [Google Scholar] [CrossRef]
- Liu, Y.; Zuo, S.; Xu, L.; Zou, Y.; Song, W. Study on Diversity of Endophytic Bacterial Communities in Seeds of Hybrid Maize and Their Parental Lines. Arch. Microbiol. 2012, 194, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Shi, X.-Y. Taxonomic Structure and Function of Seed-Inhabiting Bacterial Microbiota from Common Reed (Phragmites australis) and Narrowleaf Cattail (Typha angustifolia L.). Arch. Microbiol. 2018, 200, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Chesneau, G.; Torres-Cortes, G.; Briand, M.; Darrasse, A.; Preveaux, A.; Marais, C.; Jacques, M.-A.; Shade, A.; Barret, M. Temporal Dynamics of Bacterial Communities during Seed Development and Maturation. FEMS Microbiol. Ecol. 2020, 96, fiaa190. [Google Scholar] [CrossRef] [PubMed]
- Dai, H. Comparisons on the Productivity and Quality of Nutrition and Silage for Triticale, Rye and Oat in Gannan Plateau. Master’s Thesis, Gansu Agricultural University, Gansu, China, 2020. [Google Scholar]
- Zhang, Z. Seed-Borne Bacteria of Lucerne (Medicago sativa L.) and Their Pathogenicity. Ph.D. Thesis, Lanzhou University, Gansu, China, 2013. [Google Scholar]
- Mahmud, S.A.; Qureshi, M.A.; Sapkota, M.; Pellegrino, M.W. A Pathogen Branched-Chain Amino Acid Catabolic Pathway Subverts Host Survival by Impairing Energy Metabolism and the Mitochondrial UPR. PLoS Pathog. 2020, 16, e1008918. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Smith, L.; Liu, A.; Lu, S.-E. Genetic and Biochemical Map for the Biosynthesis of Occidiofungin, an Antifungal Produced by Burkholderia Contaminans Strain MS14. Appl. Environ. Microbiol. 2011, 77, 6189–6198. [Google Scholar] [CrossRef] [PubMed]
- GB/T 38483-2020; Determination of Antibacterial Activity for Microbial Secondary Metabolites-INHIBITION Zone Metho. China Jiliang University: Beijing, China; China National Institute of Standardization: Beijing, China; Standards Press of China: Beijing, China, 2020; Volume 11.
- GB/T 20287-2006; Microbial Inoculants in Agriculture. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration. Standards Press of China: Beijing, China, 2006; Volume 09.
- Rossolini, G.M.; Franceschini, N.; Riccio, M.L.; Mercuri, P.S.; Perilli, M.; Galleni, M.; Frere, J.-M.; Amicosante, G. Characterization and Sequence of the Chryseobacterium (Flavobacterium) Meningosepticum Carbapenemase: A New Molecular Class B β-Lactamase Showing a Broad Substrate Profile. Biochem. J. 1998, 332, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Grum, M.; Camloh, M.; Rudolph, K.; Ravnikar, M. Elimination of Bean Seed-Borne Bacteria by Thermotherapy and Meristem Culture. In Pathogen and Microbial Contamination Management in Micropropagation; Cassells, A.C., Ed.; Developments in Plant Pathology; Springer: Dordrecht, The Netherlands, 1997; Volume 12, pp. 225–231. ISBN 978-90-481-4932-2. [Google Scholar]
- Rahman, M.M.; Flory, E.; Koyro, H.-W.; Abideen, Z.; Schikora, A.; Suarez, C.; Schnell, S.; Cardinale, M. Consistent Associations with Beneficial Bacteria in the Seed Endosphere of Barley (Hordeum vulgare L.). Syst. Appl. Microbiol. 2018, 41, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Furuya, T. Diversity and Characteristics of Culturable Endophytic Bacteria from Passiflora edulis Seeds. MicrobiologyOpen 2021, 10, e1226. [Google Scholar] [CrossRef]
- Tkalec, V.; Mahnic, A.; Gselman, P.; Rupnik, M. Analysis of Seed-Associated Bacteria and Fungi on Staple Crops Using the Cultivation and Metagenomic Approaches. Folia Microbiol. 2022, 67, 351–361. [Google Scholar] [CrossRef]
- Newcombe, G.; Marlin, M.; Barge, E.; Heitmann, S.; Ridout, M.; Busby, P.E. Plant Seeds Commonly Host Bacillus Spp., Potential Antagonists of Phytopathogens. Microb. Ecol. 2023, 85, 1356–1366. [Google Scholar] [CrossRef]
- Cottyn, B.; Debode, J.; Regalado, E.; Mew, T.W.; Swings, J. Phenotypic and Genetic Diversity of Rice Seed-Associated Bacteria and Their Role in Pathogenicity and Biological Control. J. Appl. Microbiol. 2009, 107, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; William Costerton, J. Antibiotic Resistance of Bacteria in Biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Hol, F.J.H.; Hubert, B.; Dekker, C.; Keymer, J.E. Density-Dependent Adaptive Resistance Allows Swimming Bacteria to Colonize an Antibiotic Gradient. ISME J. 2016, 10, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Cutugno, L.; Mc Cafferty, J.; Pané-Farré, J.; O’Byrne, C.; Boyd, A. rpoB Mutations Conferring Rifampicin-Resistance Affect Growth, Stress Response and Motility in Vibrio Vulnificus. Microbiol. Read. Engl. 2020, 166, 1160–1170. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental Pollution by Antibiotics and by Antibiotic Resistance Determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- White, D.G.; McDermott, P.F. Emergence and Transfer of Antibacterial Resistance. J. Dairy Sci. 2001, 84, E151–E155. [Google Scholar] [CrossRef]
- Liu, K.; Ding, X.; Wang, J. Soil Metabolome Correlates with Bacterial Diversity and Co-Occurrence Patterns in Root-Associated Soils on the Tibetan Plateau. Sci. Total Environ. 2020, 735, 139572. [Google Scholar] [CrossRef]
- Fu, H.; Zhang, L.; Fan, C.; Li, W.; Liu, C.; Zhang, H.; Cheng, Q.; Zhang, Y. Sympatric Yaks and Plateau Pikas Promote Microbial Diversity and Similarity by the Mutual Utilization of Gut Microbiota. Microorganisms 2021, 9, 1890. [Google Scholar] [CrossRef]
- Huang, J.; Li, Y.; Luo, Y. Bacterial Community in the Rumen of Tibetan Sheep and Gansu Alpine Fine-Wool Sheep Grazing on the Qinghai-Tibetan Plateau, China. J. Gen. Appl. Microbiol. 2017, 63, 122–130. [Google Scholar] [CrossRef]
- Guo, X.; Sha, Y.; Lv, W.; Pu, X.; Liu, X.; Luo, Y.; Hu, J.; Wang, J.; Li, S.; Zhao, Z. Sex Differences in Rumen Fermentation and Microbiota of Tibetan Goat. Microb. Cell Factories 2022, 21, 55. [Google Scholar] [CrossRef]
- Zeng, B.; Zhang, S.; Xu, H.; Kong, F.; Yu, X.; Wang, P.; Yang, M.; Li, D.; Zhang, M.; Ni, Q.; et al. Gut Microbiota of Tibetans and Tibetan Pigs Varies between High and Low Altitude Environments. Microbiol. Res. 2020, 235, 126447. [Google Scholar] [CrossRef]
- Dong, K.; Wu, K.; Zheng, T.; Yue, J.; Wang, W.; Luo, R.; You, L.; He, X.; Li, J.; Hong, Z.; et al. Comparative Study of Oral Bacteria and Fungi Microbiota in Tibetan and Chinese Han Living at Different Altitude. Tohoku J. Exp. Med. 2021, 254, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-Resistant Enterococci: A Review of Antimicrobial Resistance Mechanisms and Perspectives of Human and Animal Health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, T.; Li, X.; Wang, G.; Lin, Q.; Qu, J. Gut Microbiota in Tibetan Herdsmen Reflects the Degree of Urbanization. Front. Microbiol. 2018, 9, 1745. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, J.; Shi, D.; Yang, Z.; Zhou, S.; Yang, D.; Chen, T.; Li, J.; Li, H.; Jin, M. Landscape of Antibiotic Resistance Genes and Bacterial Communities in Groundwater on the Tibetan Plateau, and Distinguishing Their Difference with Low-Altitude Counterparts. J. Hazard. Mater. 2023, 459, 132300. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Zhao, R.; Su, G.; Liu, B.; Liu, W.; Xu, J.; Li, Q.; Meng, J. Metagenomic Surveillance of Antibiotic Resistome in Influent and Effluent of Wastewater Treatment Plants Located on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2023, 870, 162031. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, M.; Yu, C.; Wu, Q.; Mao, Z.; Li, H.; Ren, L.; Zeng, J.; Xing, P.; Zhou, L.-J.; et al. Unexpected Enrichment of Antibiotic Resistance Genes and Organic Remediation Genes in High-Altitude Lakes at Eastern Tibetan Plateau. Sci. Total Environ. 2023, 874, 162554. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Antibiotics | Inhibition Rate (I) | ||||
|---|---|---|---|---|---|---|
| Name | Antibiotic Concentration (μg/mL) | 40 | 640 | |||
| CK | 40 | 640 | ||||
| G2-17 | KAN | 8.00 | 9.38 ± 0.166 | 12.60 ± 0.100 | 17.3% | 57.5% |
| G3-8 | TET | 8.00 | 9.70 ± 0.200 | 13.10 ± 0.400 | 21.3% | 63.8% |
| XG3-8 | CPFX | 10.30 ± 0.224 | 10.92 ± 0.287 | 18.72 ± 0.185 | 6.0% | 81.7% |
| XG3-8 | EM | 8.00 | 8.50 | 13.10 ± 0.400 | 6.3% | 63.8% |
| Strain | Antibiotics | Linear Regression Analysis | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Name | Bacteriostatic Rate Corresponding to Antibiotic Concentration | Regression Equation | R2 | IC50 | |||||
| 40 | 80 | 160 | 320 | 640 | |||||
| G2-17 | KAN | 17.30% | 31.50% | 38.80% | 47.50% | 57.50% | Y = 32.023X − 32.063 | 0.9881 | 365 |
| G3-8 | TET | 21.30% | 21.30% | 33.80% | 41.30% | 63.80% | Y = 32.973X − 35.757 | 0.8703 | 399 |
| XG3-8 | EM | 6.30% | 15.00% | 27.00% | 39.30% | 63.80% | Y = 46.274X − 71.714 | 0.9586 | 427 |
| XG3-8 | CPFX | 6.02% | 24.47% | 53.59% | 69.90% | 81.75% | Y = 65.405X − 97.015 | 0.9784 | 177 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Yang, J.; Xie, J.; Zhu, S.; Zhang, Z. Diversity and Antibiotic Resistance of Triticale Seed-Borne Bacteria on the Tibetan Plateau. Microorganisms 2024, 12, 650. https://doi.org/10.3390/microorganisms12040650
Hou X, Yang J, Xie J, Zhu S, Zhang Z. Diversity and Antibiotic Resistance of Triticale Seed-Borne Bacteria on the Tibetan Plateau. Microorganisms. 2024; 12(4):650. https://doi.org/10.3390/microorganisms12040650
Chicago/Turabian StyleHou, Xuan, Jie Yang, Jinjing Xie, Shaowei Zhu, and Zhenfen Zhang. 2024. "Diversity and Antibiotic Resistance of Triticale Seed-Borne Bacteria on the Tibetan Plateau" Microorganisms 12, no. 4: 650. https://doi.org/10.3390/microorganisms12040650
APA StyleHou, X., Yang, J., Xie, J., Zhu, S., & Zhang, Z. (2024). Diversity and Antibiotic Resistance of Triticale Seed-Borne Bacteria on the Tibetan Plateau. Microorganisms, 12(4), 650. https://doi.org/10.3390/microorganisms12040650