
The Complete Genome of a Novel Typical Species Thiocapsa bogorovii and Analysis of Its Central Metabolic Pathways

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Genome Sequencing, Assembly, and Annotation
3. Results and Discussion
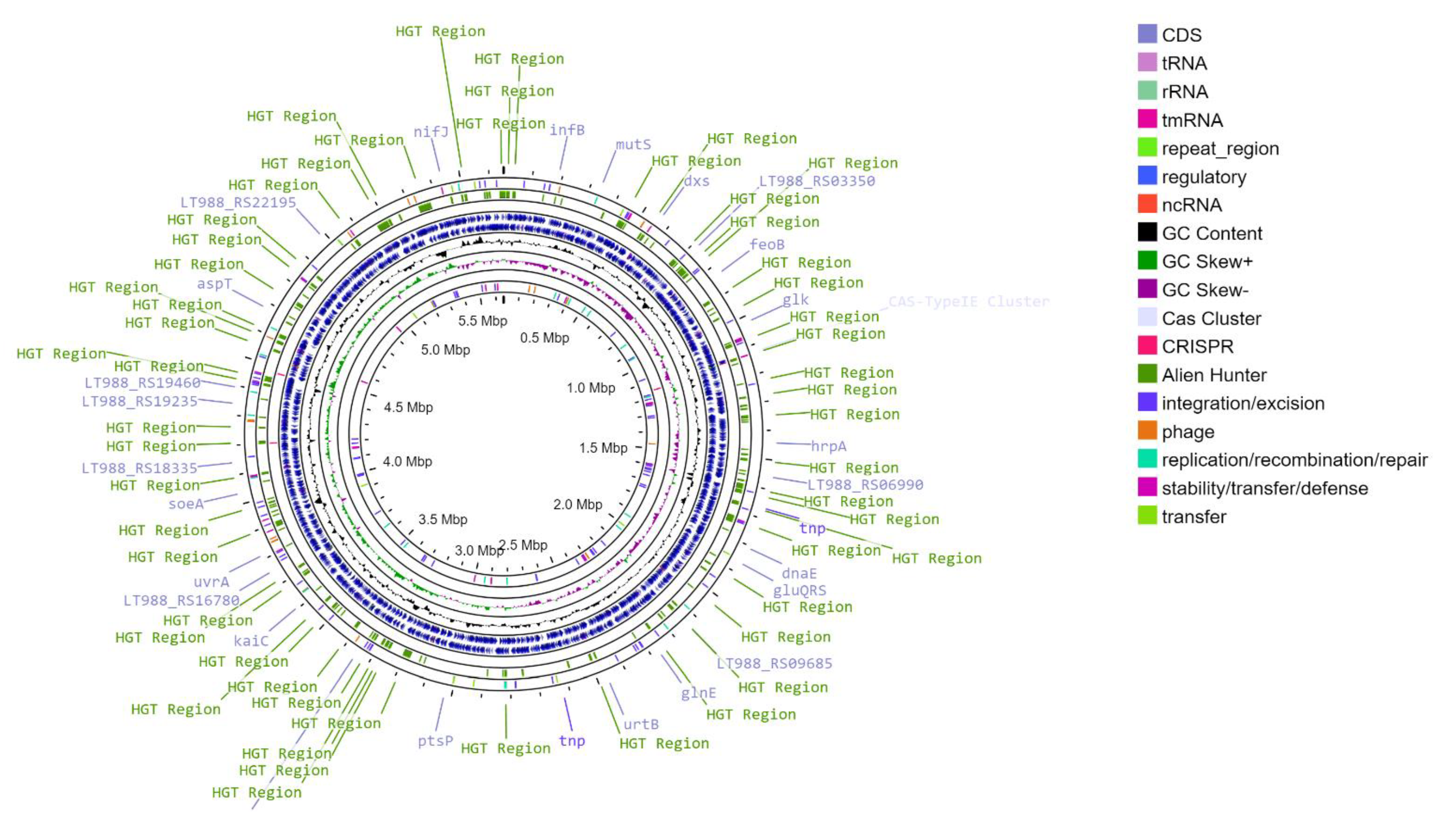
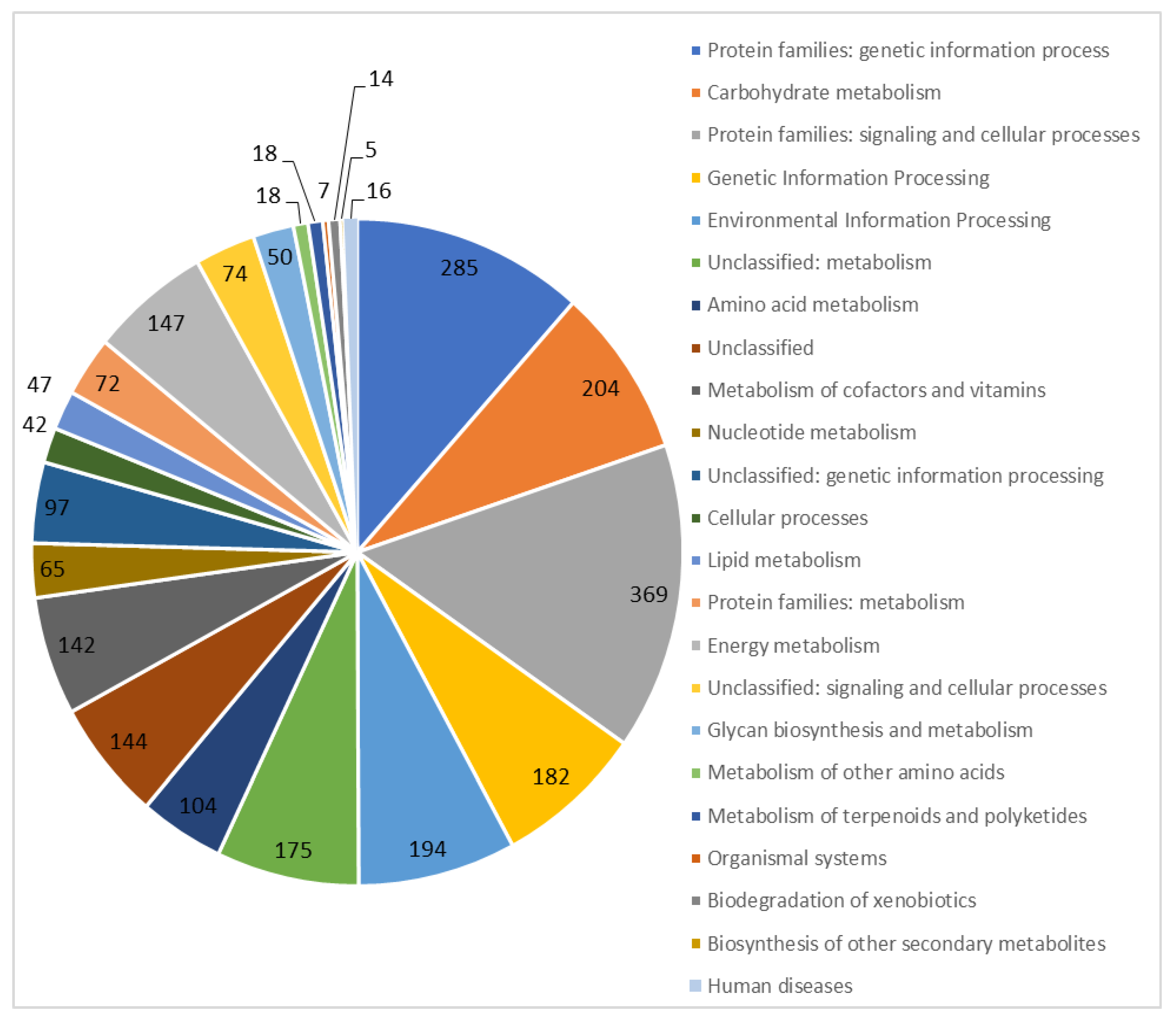
3.1. General Characteristics of the T. roseopersicina BBS Genome
3.2. Validating the Taxonomic Position of the BBS Strain
3.3. A Comparison of the Whole Genomes of T. bogorovii BBS and Alc. Vinosum DSM 180T
3.3.1. Pigment Biosynthesis and Photocomplexes
3.3.2. Autotrophy and RuBisCO
3.3.3. Heterotrophy
3.3.4. Hydrogenases
3.3.5. Chemotrophic Metabolism
3.3.6. Nitrogen Metabolism
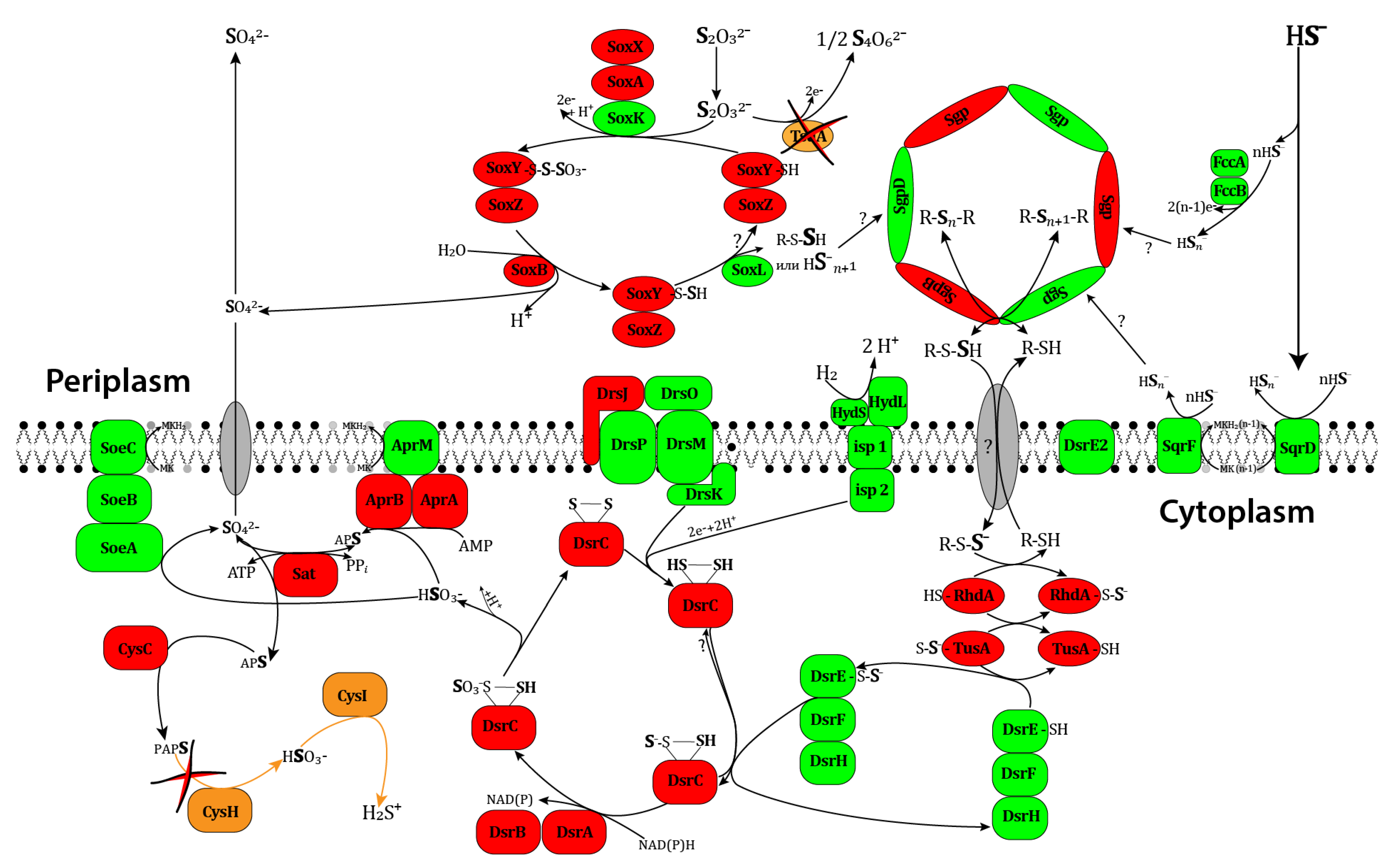
3.3.7. Sulfur Metabolism
Thiosulfate Oxidation
Sulfide Oxidation
Oxidizing Elemental Sulfur
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| Aa, aa—amino acids |
| Alc. vinosum—Allochromatium vinosum |
| ANI—Average Nucleotide Identity |
| ATCC—American Type Culture Collection |
| ATP—adenosine triphosphate |
| bp—base pairs |
| CRISPR—clustered regularly interspaced short palindromic repeats |
| DDH—digital DNA-DNA hybridization parameter |
| DNA—deoxyribonucleic acid |
| e-value—the expectation value |
| FAD—flavin adenine dinucleotide |
| FccAB—flavocytochrome c sulfide dehydrogenase |
| FCSD—flavocytochrome c sulfide dehydrogenase |
| GC content—guanine-cytosine content |
| HiPIP—High potential iron–sulfur protein |
| kbp—kilobase pairs |
| kDa—kilodalton |
| KEGG—Kyoto Encyclopedia of Genes and Genomes |
| LH—light-harvesting antenna complex |
| LPSN—List of Prokaryotic names with Standing in Nomenclature |
| Mb—megabase |
| Mo-containing—molybdenum-containing |
| NAD—nicotinamide adenine dinucleotide |
| NAD+—the oxidized form |
| NADH—the reduced form |
| NADP—nicotinamide adenine dinucleotide phosphate |
| NAD(P)H—the reduced form |
| ncRNAs—non-coding RNA |
| ONT—Oxford Nanopore Technologies |
| PEP—phosphoenolpyruvate |
| PRK—Phosphoribulokinase |
| PTS—phosphotransferase system |
| PufX—intrinsic membrane protein |
| RC—reaction center |
| Rhd—rhodanese-like protein |
| RLP—RubisCO-like protein |
| RNA—ribonucleic acid |
| rRNA—ribosomal RNA |
| rTCA—reductive tricarboxylic acid cycle |
| RuBisCO—Ribulose-1,5-bisphosphate carboxylase/oxygenase |
| SBP—Sedoheptulose-bisphosphatase |
| Sgp—sulfur globule proteins |
| Sox system—sulfur oxidation system |
| SQR—sulfide/quinone oxidoreductase |
| T. bogorovii—Thiocapsa bogorovii |
| T. imhoffii—Thiocapsa imhoffii |
| T. marina—Thiocapsa marina |
| T. rosea—Thiocapsa rosea |
| T. roseopersicina—Thiocapsa roseopersicina |
| TCA cycle—the tricarboxylic acid cycle |
| tRNA—transfer RNA |
| TsdA—tetrathionate reductase |
References
- Bogorov, L.V. The Properties of Thiocapsa Roseopersicina, Strain BBS, Isolated from an Estuary of the White Sea. Mikrobiologiia 1974, 43, 326–332. [Google Scholar] [PubMed]
- Kondratieva, E.N.; Zhukov, V.G.; Ivanovsky, R.N.; Petushkova, Y.P.; Monosov, E.Z. The Capacity of Phototrophic Sulfur Bacterium Thiocapsa Roseopersicina for Chemosynthesis. Arch. Microbiol. 1976, 108, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Zhukov, V.G. Formation of Ribuloso-1,5-Diphosphate Carboxylase by Thiocapsa Roseopersicina under Different Growth Conditions. Mikrobiologiia 1976, 45, 915–917. [Google Scholar] [PubMed]
- Petushkova, I.P.; Ivanovskiĭ, R.N. Enzymes Involved in Thiosulfate Metabolism in Thiocapsa Roseopersicina under Various Conditions of Growth. Mikrobiologiia 1976, 45, 960–965. [Google Scholar] [PubMed]
- Petushkova, Y.P.; Ivanovsky, R.N. Sulphite Oxidation by Thiocapsa Roseopersicina (Russian). Mikrobiologiya 1976, 45, 592–597. [Google Scholar]
- Gogotov, I.N. Hydrogenases of Phototrophic Microorganisms. Biochimie 1986, 68, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Kondratieva, E.N.; Ivanovsky, R.N.; Krasilnikova, E.N. Light and Dark Metabolism in Purpule Sulfur Bacteria. Sov. Sci. Rev. 1981, 13, 325–364. [Google Scholar]
- Kovács, A.T.; Rákhely, G.; Kovács, K.L. Genes Involved in the Biosynthesis of Photosynthetic Pigments in the Purple Sulfur Photosynthetic Bacterium Thiocapsa Roseopersicina. Appl. Environ. Microbiol. 2003, 69, 3093–3102. [Google Scholar] [CrossRef]
- Kovács, K.L.; Kovács, Á.T.; Maróti, G.; Mészáros, L.S.; Balogh, J.; Latinovics, D.; Fülöp, A.; Dávid, R.; Dorogházi, E.; Rákhely, G. The Hydrogenases of Thiocapsa roseopersicina. Biochem. Soc. Trans. 2005, 33, 61–63. [Google Scholar] [CrossRef]
- Laurinavichene, T.V.; Rákhely, G.; Kovács, K.L.; Tsygankov, A.A. The Effect of Sulfur Compounds on H2 Evolution/Consumption Reactions, Mediated by Various Hydrogenases, in the Purple Sulfur Bacterium, Thiocapsa Roseopersicina. Arch. Microbiol. 2007, 188, 403–410. [Google Scholar] [CrossRef]
- Palágyi-Mészáros, L.S.; Maróti, J.; Latinovics, D.; Balogh, T.; Klement, E.; Medzihradszky, K.F.; Rákhely, G.; Kovács, K.L. Electron-Transfer Subunits of the NiFe Hydrogenases in Thiocapsa Roseopersicina BBS. FEBS J. 2009, 276, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Maróti, G.; Fodor, B.D.; Rákhely, G.; Kovács, Á.T.; Arvani, S.; Kovács, K.L. Accessory Proteins Functioning Selectively and Pleiotropically in the Biosynthesis of [Nife] Hydrogenases in Thiocapsa Roseoporsicina. Eur. J. Biochem. 2003, 270, 2218–2227. [Google Scholar] [CrossRef] [PubMed]
- Maróti, J.; Farkas, A.; Nagy, I.K.; Maróti, G.; Kondorosi, É.; Rákhely, G.; Kovács, K.L. A Second Soluble Hox-Type NiFe Enzyme Completes the Hydrosenase Set in Thiocapsa Roseopersicina BBS. Appl. Environ. Microbiol. 2010, 76, 5113–5123. [Google Scholar] [CrossRef] [PubMed]
- Tengölics, R.; Mészáros, L.; Gyori, E.; Doffkay, Z.; Kovács, K.L.; Rákhely, G. Connection between the Membrane Electron Transport System and Hyn Hydrogenase in the Purple Sulfur Bacterium, Thiocapsa Roseopersicina BBS. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shawand, P.D.; Marshall, D. Using Tablet for Visual Exploration of Second-Generation Sequencing Data. Brief. Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; Van Nimwegen, E. Automated Reconstruction of Whole-Genome Phylogenies from Short-Sequence Reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and Linear Interactive Genome Visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for Integration and Interpretation of Large-Scale Molecular Data Sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.S.; Parkhill, J. Interpolated Variable Order Motifs for Identification of Horizontally Acquired DNA: Revisiting the Salmonella Pathogenicity Islands. Bioinformatics 2006, 22, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an Update of CRISRFinder, Includes a Portable Version, Enhanced Performance and Integrates Search for Cas Proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: A Hub for Protein Information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Tourova, T.P.; Keppen, O.I.; Kovaleva, O.L.; Slobodova, N.V.; Berg, I.A.; Ivanovsky, R.N. Phylogenetic Characterization of the Purple Sulfur Bacterium Thiocapsa Sp. BBS by Analysis of the 16S RRNA, CbbL, and NifH Genes and Its Description as Thiocapsa bogorovii sp. nov., a New Species. Microbiology 2009, 78, 339–349. [Google Scholar] [CrossRef]
- Weissgerber, T.; Zigann, R.; Bruce, D.; Chang, Y.J.; Detter, J.C.; Han, C.; Hauser, L.; Jeffries, C.D.; Land, M.; Munk, C.; et al. Complete Genome Sequence of Allochromatium Vinosum DSM 180T. Stand. Genom. Sci. 2011, 5, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Ouchane, S.; Picaud, M.; Vernotte, C.; Astier, C. Photooxidative Stress Stimulates Illegitimate Recombination and Mutability in Carotenoid-Less Mutants of Rubrivivax Gelatinosus. EMBO J. 1997, 16, 4777–4787. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, B.; Lunceford, C.; Lin, S.; Woronowicz, K.; Niederman, R.A.; Woodbury, N.W. Energy Transfer Properties of Rhodobacter Sphaeroides Chromatophores during Adaptation to Low Light Intensity. Phys. Chem. Chem. Phys. 2014, 16, 17133–17141. [Google Scholar] [CrossRef] [PubMed]
- Cogdell, R.J.; Roszak, A.W. Structural Biology: The Purple Heart of Photosynthesis. Nature 2014, 508, 196–197. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.; Papiz, M.Z.; Jackson, P.J.; Brindley, A.A.; Ng, I.W.; Olsen, J.D.; Dickman, M.J.; Bullough, P.A.; Hunter, C.N. Three-Dimensional Structure of the Rhodobacter Sphaeroides Rc-Lh1-Pufx Complex: Dimerization and Quinone Channels Promoted by PufX. Biochemistry 2013, 52, 7575–7585. [Google Scholar] [CrossRef] [PubMed]
- Roszak, A.W.; Howard, T.D.; Southall, J.; Gardiner, A.T.; Law, C.J.; Isaacs, N.W.; Cogdell, R.J. Crystal Structure of the RC-LH1 Core Complex from Rhodopseudomonas Palustris. Science 2003, 302, 1969–1972. [Google Scholar] [CrossRef]
- Niwa, S.; Yu, L.J.; Takeda, K.; Hirano, Y.; Kawakami, T.; Wang-Otomo, Z.Y.; Miki, K. Structure of the LH1-RC Complex from Thermochromatium Tepidum at 3.0 Å. Nature 2014, 508, 228–232. [Google Scholar] [CrossRef]
- Liu, L.N.; Bracun, L.; Li, M. Structural Diversity and Modularity of Photosynthetic RC−LH1 Complexes. Trends Microbiol. 2023, 32, 38–52. [Google Scholar] [CrossRef]
- Tabita, F.R. The Biochemistry and Metabolic Regulation of Carbon Metabolism and CO2 Fixation in Purple Bacteria. In Anoxygenic Photosynthetic Bacteria; Kluwer Academic Publishers: Dordrecht, Germany, 1995; pp. 885–914. [Google Scholar]
- McFadden, B.A.; Shively, L.M. Variations in Autotrophic Life; Academic Press: London, UK, 1991; pp. 25–49. [Google Scholar]
- Tabita, F.R.; Hanson, T.E.; Li, H.; Satagopan, S.; Singh, J.; Chan, S. Function, Structure, and Evolution of the RubisCO-Like Proteins and Their RubisCO Homologs. Microbiol. Mol. Biol. Rev. 2007, 71, 576–599. [Google Scholar] [CrossRef]
- Hanson, T.E.; Tabita, F.R. Insights into the Stress Response and Sulfur Metabolism Revealed by Proteome Analysis of a Chlorobium Tepidum Mutant Lacking the Rubisco-like Protein. Photosynth. Res. 2003, 78, 231–248. [Google Scholar] [CrossRef]
- Hanson, T.E.; Tabita, F.R. A Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase (RubisCO)-like Protein from Chlorobium Tepidum That Is Involved with Sulfur Metabolism and the Response to Oxidative Stress. Proc. Natl. Acad. Sci. USA 2001, 98, 4397–4402. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Bek, E.J. Multiple Rubisco Forms in Proteobacteria: Their Functional Significance in Relation to CO2 Acquisition by the CBB Cycle. J. Exp. Bot. 2008, 59, 1525–1541. [Google Scholar] [CrossRef] [PubMed]
- Purohit, K.; McFadden, B.A. Ribulose 1,5-Bisphosphate Carboxylase and Oxygenase from Thiocapsa Roseopersicina: Activation and Catalysis. Arch. Biochem. Biophys. 1979, 194, 101–106. [Google Scholar] [CrossRef]
- Badger, M.R.; Price, G.D. CO2 Concentrating Mechanisms in Cyanobacteria: Molecular Components, Their Diversity and Evolution. J. Exp. Bot. 2003, 54, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Kerfeld, C.A.; Sawaya, M.R.; Tanaka, S.; Nguyen, C.V.; Phillips, M.; Beeby, M.; Yeates, T.O. Microbiology: Protein Structures Forming the Shell of Primitive Bacterial Organelles. Science 2005, 309, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Kondratieva, E.N. Interrelation between Modes of Carbon Assimilation and Energy Production in Phototropic Purple and Green Bacteria. Int. Rev. Biochem. 1979, 21, 117–175. [Google Scholar]
- Takai, K.; Campbell, B.J.; Cary, S.C.; Suzuki, M.; Oida, H.; Nunoura, T.; Hirayama, H.; Nakagawa, S.; Suzuki, Y.; Inagaki, F.; et al. Enzymatic and Genetic Characterization of Carbon and Energy Metabolisms by Deep-Sea Hydrothermal Chemolithoautotrophic Isolates of Epsilonproteobacteria. Appl. Environ. Microbiol. 2005, 71, 7310–7320. [Google Scholar] [CrossRef]
- Aoshima, M.; Ishii, M.; Igarashi, Y. A Novel Enzyme, Citryl-CoA Synthetase, Catalysing the First Step of the Citrate Cleavage Reaction in Hydrogenobacter Thermophilus TK-6. Mol. Microbiol. 2004, 52, 751–761. [Google Scholar] [CrossRef]
- Vignais, P.M.; Billoud, B.; Meyer, J. Classification and Phylogeny of Hydrogenases. FEMS Microbiol. Rev. 2001, 25, 455–501. [Google Scholar] [CrossRef]
- Adams, M.W.W.; Mortenson, L.E.; Chen, J.-S. Hydrogenase. Biochim. Biophys. Acta (BBA)—Rev. Bioenerg. 1980, 594, 105–176. [Google Scholar] [CrossRef]
- Søndergaard, D.; Pedersen, C.N.S.; Greening, C. HydDB: A Web Tool for Hydrogenase Classification and Analysis. Sci. Rep. 2016, 6, srep34212. [Google Scholar] [CrossRef]
- Colbeau, A.; Kovacs, K.L.; Chabert, J.; Vignais, P.M. Cloning and Sequences of the Structural (HupSLC) and Accessory (HupDHI) Genes for Hydrogenase Biosynthesis in Thiocapsa Roseopersicina. Gene 1994, 140, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Fodor, B.; Rákhely, G.; Kovács, Á.T.; Kovács, K.L. Transposon Mutagenesis in Purple Sulfur Photosynthetic Bacteria: Identification of HypF, Encoding a Protein Capable of Processing [NiFe] Hydrogenases in α, β, and γ Subdivisions of the Proteobacteria. Appl. Environ. Microbiol. 2001, 67, 2476–2483. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kovács, K.L.; Fodor, B.; Kovács, Á.T.; Csanádi, G.; Maróti, G.; Balogh, J.; Arvani, S.; Rákhely, G. Hydrogenases, Accessory Genes and the Regulation of [NiFe] Hydrogenase Biosynthesis in Thiocapsa Roseopersicina. Int. J. Hydrogen Energy 2002, 27, 1463–1469. [Google Scholar] [CrossRef]
- Gogotov, I.N.; Zorin, N.A.; Serebriakova, L.T.; Kondratieva, E.N. The Properties of Hydrogenase from Thiocapsa Roseopersicina. BBA—Enzymol. 1978, 523, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Bagyinka, C.; Zorin, N.A.; Kovács, K.L. Unconsidered Factors Affecting Hydrogenase Activity Measurement. Anal. Biochem. 1984, 142, 7–15. [Google Scholar] [CrossRef]
- Kovács, Á.T.; Rákhely, G.; Balogh, J.; Maróti, G.; Cournac, L.; Carrier, P.; Mészáros, L.S.; Peltier, G.; Kovàcs, K.L. Hydrogen Independent Expression of HupSL Genes in Thiocapsa Roseopersicina BBS. FEBS J. 2005, 272, 4807–4816. [Google Scholar] [CrossRef]
- Rákhely, G.; Kovács, Á.T.; Maróti, G.; Fodor, B.D.; Csanádi, G.; Latinovics, D.; Kovács, K.L. Cyanobacterial-Type, Heteropentameric, NAD+-Reducing NiFe Hydrogenase in the Purple Sulfur Photosynthetic Bacterium Thiocapsa Roseopersicina. Appl. Environ. Microbiol. 2004, 70, 722–728. [Google Scholar] [CrossRef]
- Rákhely, G.; Laurinavichene, T.V.; Tsygankov, A.A.; Kovács, K.L. The Role of Hox Hydrogenase in the H2 Metabolism of Thiocapsa Roseopersicina. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 671–676. [Google Scholar] [CrossRef][Green Version]
- Meuer, J.; Bartoschek, S.; Koch, J.; Künkel, A.; Hedderich, R. Purification and Catalytic Properties of Ech Hydrogenase from Methanosarcina Barkeri. Eur. J. Biochem. 1999, 265, 325–335. [Google Scholar] [CrossRef]
- Dincturk, H.B.; Demir, V.; Aykanat, T. Bd Oxidase Homologue of Photosynthetic Purple Sulfur Bacterium Allochromatium Vinosum Is Co-Transcribed with a Nitrogen Fixation Related Gene. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 99, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T. Microbiology of nitrogen fixation by anoxygenic photosynthetic bacteria. In Anoxygenic Photosynthetic Bacteria. Advances in Photosynthesis and Respiration; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Springer: Dordrecht, Germany, 1995; Volume 2, pp. 915–928. [Google Scholar]
- Dahl, C. Sulfur Metabolism in Phototrophic Bacteria. In Modern Topics in the Phototrophic Prokaryotes; Springer International Publishing: Cham, Switzerland, 2017; pp. 27–66. [Google Scholar]
- Brito, J.A.; Denkmann, K.; Pereira, I.S.A.C.; Archer, M.; Dahl, C. Thiosulfate Dehydrogenase (TsdA) from Allochromatium Vinosum: Structural and Functional Insights into Thiosulfate Oxidation. J. Biol. Chem. 2015, 290, 9222–9238. [Google Scholar] [CrossRef] [PubMed]
- Brune, D.C. Isolation and Characterization of Sulfur Globule Proteins from Chromatium Vinosum and Thiocapsa Roseopersicina. Arch. Microbiol. 1995, 163, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Prange, A.; Dahl, C.; Trüper, H.G.; Chauvistré, R.; Modrow, H.; Hormes, J. X-Ray Absorption Spectroscopy of Bacterial Sulfur Globules: A Detailed Reply. Microbiology 2002, 148, 2268–2270. [Google Scholar] [CrossRef]
- Frigaard, N.U.; Dahl, C. Sulfur metabolism in phototrophic sulfur bacteria. Adv. Microb. Physiol. 2009, 54, 103–200. [Google Scholar] [PubMed]
- Stockdreher, Y.; Sturm, M.; Josten, M.; Sahl, H.G.; Dobler, N.; Zigann, R.; Dahl, C. New Proteins Involved in Sulfur Trafficking in the Cytoplasm of Allochromatium Vinosum. J. Biol. Chem. 2014, 289, 12390–12403. [Google Scholar] [CrossRef] [PubMed]
- Tsygankov, A.A.; Zorin, N.A.; Starodubov, A.S.; Khasimov, M.K.; Melnikova, M.S.; Doronin, I.A.; Vasilov, R.G. A Simple Method for Oriented Immobilization of HydSL Hydrogenase of Thiocapsa Bogorovii on Carbon Electrodes. Int. J. Hydrog. Energy 2023, 48, 39989–39999. [Google Scholar] [CrossRef]
- Tsygankov, A.A.; Khusnutdinova, A.N. Hydrogen in Metabolism of Purple Bacteria and Prospects of Practical Application. Microbiology 2015, 84, 1–22. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | T. roseopersicina BBS | Alc. vinosum DT (DSM 180T) |
|---|---|---|
| GenBank accession number(s) | NZ_CP089309.1 | CP001896; CP001897; CP001898 |
| Genome size (bp) | 5,649,927 | 3,669,074 |
| Number of replicons | 1 | 3 |
| Extrachromosomal elements (size) | 0 | 2 (102 kbp; 40 kbp) |
| Percent G + C content % | 63.94 | 64.2 |
| Total number of genes | 5036 | 3327 |
| Protein-encoding genes (% of total) | 4854 (96.4) | 3217 (96.7) |
| Total RNA genes | 58 | 64 |
| tRNAs | 48 | 51 |
| rRNA genes/operons | 2, 2, 2 (5S, 16S, 23S) | 9/3 |
| ncRNAs | 4 | 4 |
| Putative pseudogenes | 124 | 46 |
| CRISPR arrays | 1 | 3 |
| Name of Organism | ANI Value, % | DDH, % |
|---|---|---|
| T. marina 5811T | 89.03 | 50.9 |
| T. rosea DSM235T | 86.92 | 38.3 |
| T. imhoffii DSM21303 | 75.58 | 14.6 |
| T. roseopersicina DSM217T | 87.22 | 43.2 |
| Category | T. bogorovii BBS | Alc. vinosum DSM 180 T | ||
|---|---|---|---|---|
| No. of Genes | % Genome Content | No. of Genes | % Genome Content | |
| Protein metabolism | 316 | 10.33 | 264 | 10.85 |
| Vitamins, cofactors, prosthetic groups, and photosynthetic pigments | 284 | 9.28 | 249 | 10.23 |
| Amino acids and derivatives | 307 | 10.03 | 235 | 9.66 |
| RNA metabolism | 191 | 6.24 | 162 | 6.66 |
| Respiration | 196 | 6.41 | 178 | 7.31 |
| Carbohydrate metabolism | 288 | 9.41 | 183 | 7.52 |
| Motility and chemotaxis | 0 | 0 | 161 | 6.62 |
| Cell wall structure | 182 | 5.95 | 161 | 6.62 |
| DNA metabolism | 183 | 5.98 | 76 | 3.12 |
| Membrane transport | 164 | 5.36 | 102 | 4.19 |
| Fatty acids, lipids, and isoprenoids | 103 | 3.37 | 88 | 3.62 |
| Stress response | 154 | 5.03 | 107 | 4.4 |
| Virulence (disease and defense) | 103 | 3.37 | 77 | 3.2 |
| Nucleosides and nucleotides | 79 | 2.58 | 61 | 2.51 |
| Phosphorus metabolism | 54 | 1.77 | 43 | 1.8 |
| Cell division | 37 | 1.21 | 39 | 1.6 |
| Sulfur metabolism | 64 | 2.1 | 52 | 2.14 |
| Nitrogen metabolism | 89 | 2.91 | 39 | 1.6 |
| Potassium metabolism | 37 | 1.21 | 20 | 0.82 |
| № | Gene Designation and Function According to Kovács et al. [8] | Function Number, Enzyme Annotation, and Number (According to KEGG Orthology Database) | Gene ID | Gene Name and Annotation Presented in GenBank |
|---|---|---|---|---|
| 1 | -* | K08926 light-harvesting complex 1 alpha chain | LT988_08850 | light-harvesting protein |
| 2 | - | K08927 light-harvesting complex 1 beta chain | LT988_08855 | light-harvesting protein |
| 3 | - | K08926 light-harvesting complex 1 alpha chain | LT988_08860 | light-harvesting protein |
| 4 | - | K13992 photosynthetic reaction center cytochrome c subunit | LT988_08865 | photosynthetic reaction center cytochrome c subunit |
| 5 | - | K08929 photosynthetic reaction center M subunit | LT988_08870 | pufM; photosynthetic reaction center subunit M |
| 6 | - | K08928 photosynthetic reaction center L subunit | LT988_08875 | pufL; photosynthetic reaction center subunit L |
| 7 | - | K08926 light-harvesting complex 1 alpha chain | LT988_08880 | light-harvesting protein |
| 8 | - | K08927 light-harvesting complex 1 beta chain | LT988_08885 | light-harvesting protein |
| 9 | - | no KO assigned | LT988_08890 | pseudogene |
| 10 | - | K11335 3,8-divinyl chlorophyllide a/chlorophyllide a reductase subunit Z [EC:1.3.7.14 1.3.7.15] | LT988_08895 | bchZ; chlorophyllide a reductase subunit Z |
| 11 | - | K11334 3,8-divinyl chlorophyllide a/chlorophyllide a reductase subunit Y [EC:1.3.7.14 1.3.7.15] | LT988_08900 | bchY; chlorophyllide a reductase subunit Y |
| 12 | bchX, Bacteriochlorophyllide reductase subunit | K11333 3,8-divinyl chlorophyllide a/chlorophyllide a reductase subunit X [EC:1.3.7.14 1.3.7.15] | LT988_08905 | chlorophyllide a reductase iron protein subunit X |
| 13 | bchC(T), 2-α-Hydroxyethyl bacteriochlorophyllide oxidase, 317 aa | K11337 bacteriochlorophyllide a dehydrogenase [EC:1.1.1.396] | LT988_08910 317 aa | bchC; chlorophyll synthesis pathway protein BchC |
| 14 | crtF(G), Hydroxyneurosporene methyltransferase, 371 aa | K09846 demethylspheroidene O-methyltransferase [EC:2.1.1.210] | LT988_08915 371 aa | acetylserotonin O-methyltransferase |
| 15 | crtE, Geranylgeranyl pyrophosphate synthase, 288 aa | K13789 geranylgeranyl diphosphate synthase, type II [EC:2.5.1.1 2.5.1.10 2.5.1.29] | LT988_08920 288 aa | polyprenyl synthetase family protein |
| 16 | crtD, Methoxyneurosporene dehydrogenase, 498 aa | K09845 1-hydroxycarotenoid 3,4-desaturase [EC:1.3.99.27] | LT988_08925 498 aa | crtI; phytoene desaturase |
| crtC, Hydroxyneurosporene dehydrogenase, 405 aa 1962948–1961731 | ||||
| 17 | - | K09844 carotenoid 1,2-hydratase [EC:4.2.1.131] | LT988_08930 277 aa | carotenoid 1,2-hydratase |
| 18 | orf495 (G), o-Succinyl-benzoic acid CoA ligase, 495 aa | K01911 o-succinylbenzoate---CoA ligase [EC:6.2.1.26] | LT988_08935 468 aa | AMP-binding protein |
| 19 | bchJ (G), 4-Vinyl reductase, 208 aa | K04036 divinyl protochlorophyllide a 8-vinyl-reductase [EC:1.-.-.-] | LT988_08940 201 aa | bchJ; bacteriochlorophyll 4-vinyl reductase |
| 20 | hemN, O2-independent coproporphyrinogen III oxidase, 453 aa | K02495 oxygen-independent coproporphyrinogen III oxidase [EC:1.3.98.3] | LT988_08945 453 aa | hemN; oxygen-independent coproporphyrinogen III oxidase |
| 21 | bchE, Mg-protoporphyrin IX monomethylester oxidative cyclase subunit, 551 aa | K04034 anaerobic magnesium-protoporphyrin IX monomethyl ester cyclase [EC:1.21.98.3] | LT988_08950 551 aa | bchE; magnesium-protoporphyrin IX monomethyl ester anaerobic oxidative cyclase |
| 22 | orf312, Hypothetical membrane protein, 312 aa | no KO assigned | LT988_08955 312 aa | DUF3623 domain-containing protein |
| 23 | orf139, Hypothetical protein, 139 aa | no KO assigned | LT988_08960 151 aa | phosphonoacetaldehyde hydrolase |
| 24 | orf218(G), Hypothetical membrane protein, 218 aa | no KO assigned | LT988_08965 218 aa | PH domain-containing protein |
| 25 | puhA, 255 aa, Photosynthetic reaction center H subunit | K13991 photosynthetic reaction center H subunit | LT988_08970 255 aa | puhA; photosynthetic reaction center subunit H |
| 26 | bchM, Mg-protoporphyrin methyltranserase, 233 aa | K03428 magnesium-protoporphyrin O-methyltransferase [EC:2.1.1.11] | LT988_08975 233 aa | bchM; magnesium-protoporphyrin IX methyltransferase |
| 27 | bchL, 294 aa, Light-independent prochlorophyllide reductase iron–sulfur-ATP-binding subunit | K04037 light-independent protochlorophyllide reductase subunit L [EC:1.3.7.7] | LT988_08980 294 aa | bchL; ferredoxin:protochlorophyllide reductase (ATP-dependent) iron–sulfur- ATP-binding protein |
| 28 | bchH, AF528191.1, 1245 aa, Mg-protoporphyrin IX chelatase H subunit | K03403 magnesium chelatase subunit H [EC:6.6.1.1] | LT988_08985 1245 aa | magnesium chelatase subunit H |
| 29 | bchB, AF528191.1, Light-independent prochlorophyllide reductase b subunit, 258 aa, | K04039 light-independent protochlorophyllide reductase subunit B [EC:1.3.7.7] | LT988_08990 534 aa | ferredoxin:protochlorophyllide reductase (ATP-dependent) subunit B |
| 30 | - | K04038 light-independent protochlorophyllide reductase subunit N [EC:1.3.7.7] | LT988_08995 | ferredoxin:protochlorophyllide reductase (ATP-dependent) subunit N |
| 31 | - | K11336 3-vinyl bacteriochlorophyllide hydratase [EC:4.2.1.165] | LT988_09000 | bchF; 2-vinyl bacteriochlorophyllide hydratase |
| 32 | - | K04040 chlorophyll/bacteriochlorophyll a synthase [EC:2.5.1.62 2.5.1.133] | LT988_09010 | chlG; chlorophyll synthase ChlG |
| 33 | - | K10960 geranylgeranyl diphosphate/geranylgeranyl-bacteriochlorophyllide a reductase [EC:1.3.1.83 1.3.1.111] | LT988_09015 | geranylgeranyl diphosphate reductase |
| 34 | - | no KO assigned | LT988_21295 | photosynthetic reaction center cytochrome c subunit |
| № | Function Number, Enzyme Annotation, and Number (According to KEGG Orthology Database) | Gene ID | Gene Name and Annotation Presented in GenBank |
|---|---|---|---|
| 1 | K01647 citrate synthase [EC:2.3.3.1] | LT988_13005 | citrate synthase |
| 1 | K01647 citrate synthase [EC:2.3.3.1] | LT988_17095 | citrate synthase |
| 2, 3 | K01681 aconitate hydratase [EC:4.2.1.3] | LT988_17115 | acnA; aconitate hydratase AcnA |
| 4, 5 | K00031 isocitrate dehydrogenase [EC:1.1.1.42] | LT988_17130 | icd; NADP-dependent isocitrate dehydrogenase |
| 6, 7 | K00164 2-oxoglutarate dehydrogenase E1 component [EC:1.2.4.2] | LT988_19210 | 2-oxoglutarate dehydrogenase E1 component |
| 8 | K00658 2-oxoglutarate dehydrogenase E2 component (dihydrolipoamide succinyltransferase) [EC:2.3.1.61] | LT988_19205 | odhB; 2-oxoglutarate dehydrogenase complex dihydrolipoyllysine-residue succinyltransferase |
| 9 | K00382 dihydrolipoyl dehydrogenase [EC:1.8.1.4] | LT988_13640 | dihydrolipoyl dehydrogenase |
| 9 | K00382 dihydrolipoyl dehydrogenase [EC:1.8.1.4] | LT988_22720 | lpdA; dihydrolipoyl dehydrogenase |
| 9 | K00382 dihydrolipoyl dehydrogenase [EC:1.8.1.4] | LT988_05285 | dihydrolipoyl dehydrogenase |
| 6a | K00174 2-oxoglutarate/2-oxoacid ferredoxin oxidoreductase subunit alpha [EC:1.2.7.3 1.2.7.11] | LT988_18060 | 2-oxoacid:acceptor oxidoreductase subunit alpha |
| 6b | K00175 2-oxoglutarate/2-oxoacid ferredoxin oxidoreductase subunit beta [EC:1.2.7.3 1.2.7.11] | LT988_18065 | 2-oxoacid:ferredoxin oxidoreductase subunit beta |
| 10 | K01902 succinyl-CoA synthetase alpha subunit [EC:6.2.1.5] | LT988_08455 | sucD; succinate--CoA ligase subunit alpha |
| K01903 succinyl-CoA synthetase beta subunit [EC:6.2.1.5] | LT988_08460 | sucC; ADP-forming succinate--CoA ligase subunit beta | |
| 11 | K00240 succinate dehydrogenase iron–sulfur subunit [EC:1.3.5.1] | LT988_09960 | sdhB; succinate dehydrogenase iron–sulfur subunit |
| K00239 succinate dehydrogenase flavoprotein subunit [EC:1.3.5.1] | LT988_09965 | sdhA; succinate dehydrogenase (quinone) flavoprotein subunit [KO:K00239] [EC:1.3.5.1] | |
| K00241 succinate dehydrogenase cytochrome b subunit | LT988_09970 | succinate dehydrogenase [KO:K00241] | |
| 11 | K00240 succinate dehydrogenase iron–sulfur subunit [EC:1.3.5.1 1.3.5.4] | LT988_04620 | succinate dehydrogenase iron–sulfur subunit |
| K00239 succinate dehydrogenase flavoprotein subunit [EC:1.3.5.1] | LT988_04625 | sdhA; succinate dehydrogenase flavoprotein subunit | |
| K00242 succinate dehydrogenase membrane anchor subunit | LT988_04630 | sdhD; succinate dehydrogenase, hydrophobic membrane anchor protein | |
| K00241 succinate dehydrogenase cytochrome b subunit | LT988_04635 | sdhC; succinate dehydrogenase, cytochrome b556 subunit | |
| 12 | K01679 fumarate hydratase, class II [EC:4.2.1.2] | LT988_23020 | fumC; class II fumarate hydratase |
| 12 | K01676 fumarate hydratase, class I [EC:4.2.1.2] | LT988_22530 | fumarate hydratase |
| 13 | K00024 malate dehydrogenase [EC:1.1.1.37] | LT988_13010 | mdh; malate dehydrogenase |
| 13 | K00116 malate dehydrogenase (quinone) [EC:1.1.5.4] | LT988_13045 | FAD-dependent oxidoreductase [KO:K00116] [EC:1.1.5.4] |
| 14 | K00174 2-oxoglutarate/2-oxoacid ferredoxin oxidoreductase subunit alpha [EC:1.2.7.3 1.2.7.11] | LT988_18060 | 2-oxoacid:acceptor oxidoreductase subunit alpha |
| K00175 2-oxoglutarate/2-oxoacid ferredoxin oxidoreductase subunit beta [EC:1.2.7.3 1.2.7.11] | LT988_18065 | 2-oxoacid:ferredoxin oxidoreductase subunit beta | |
| 15 | K15232 citryl-CoA synthetase large subunit [EC: 6.2.1.18] | - ** | - |
| 16 | K15234 citryl-CoA lyase (EC: 4.1.3.34) | - | - |
| 17 | K01637 isocitrate lyase [EC:4.1.3.1] | LT988_08090 | aceA; isocitrate lyase |
| 18 | K01638 malate synthase [EC:2.3.3.9] | LT988_06350 | malate synthase G |
| 19 | K01895 acetyl-CoA synthetase [EC:6.2.1.1] | LT988_16650 | acs; acetate--CoA ligase |
| № | Function Number, Enzyme Annotation, and Number (According to KEGG Orthology Database) | Gene ID | Gene Name and Annotation Presented in GenBank |
|---|---|---|---|
| 1 | K00845 glucokinase [EC:2.7.1.2] | LT988_04670 | glk; glucokinase |
| 2 | K01810 glucose-6-phosphate isomerase [EC:5.3.1.9] | LT988_04445 | pgi; glucose-6-phosphate isomerase |
| 2 | K01810 glucose-6-phosphate isomerase [EC:5.3.1.9] | LT988_25180 | pgi; glucose-6-phosphate isomerase |
| 3 | K21071 ATP-dependent phosphofructokinase/diphosphate-dependent phosphofructokinase [EC:2.7.1.11; 2.7.1.90] | LT988_14085 | 6-phosphofructokinase |
| 4 | K01623 fructose-bisphosphate aldolase, class I [EC:4.1.2.13] | LT988_25065 | fructose-bisphosphate aldolase class I |
| 4 | K01624 fructose-bisphosphate aldolase, class II [EC:4.1.2.13] | LT988_08070 | fba; fructose-bisphosphate aldolase class II |
| 5 | K01803 triose-phosphate isomerase (TIM) [EC:5.3.1.1] | LT988_02660 | tpiA; triose-phosphate isomerase |
| 6 | K00134 glyceraldehyde 3-phosphate dehydrogenase (phosphorylating) [EC:1.2.1.12] | LT988_08050 | gap; type I glyceraldehyde-3-phosphate dehydrogenase |
| 7 | K00927 phosphoglycerate kinase [EC:2.7.2.3] | LT988_08060 | phosphoglycerate kinase |
| 8 | K15634 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase [EC:5.4.2.11] | LT988_13795 | histidine phosphatase family protein |
| 8 | K15633 2,3-bisphosphoglycerate-independent phosphoglycerate mutase [EC:5.4.2.12] | LT988_06575 | gpmI; 2,3-bisphosphoglycerate-independent phosphoglycerate mutase |
| 9 | K01689 enolase [EC:4.2.1.11] | LT988_19115 | eno; phosphopyruvate hydratase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_01025 | pyk; pyruvate kinase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_08065 | pyk; pyruvate kinase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_22860 | pyruvate kinase |
| 11 | K03841 fructose-1,6-bisphosphatase I [EC:3.1.3.11] | LT988_07425 | class 1 fructose-bisphosphatase |
| 12 | K00172 pyruvate-ferredoxin oxidoreductase gamma subunit [EC:1.2.7.1] | LT988_13550 | 2-oxoacid:acceptor oxidoreductase family protein |
| K00169 pyruvate-ferredoxin oxidoreductase alpha subunit [EC:1.2.7.1] | LT988_13555 | hypothetical protein | |
| K00170 pyruvate-ferredoxin oxidoreductase beta subunit [EC:1.2.7.1] | LT988_13560 | thiamine pyrophosphate-dependent enzyme | |
| 12 | K03737 pyruvate-ferredoxin/flavodoxin oxidoreductase [EC:1.2.7.1 1.2.7.-] | LT988_24065 | 2-oxoacid:acceptor oxidoreductase family protein |
| K03737 pyruvate-ferredoxin/flavodoxin oxidoreductase [EC:1.2.7.1 1.2.7.-] | LT988_24180 | nifJ; pyruvate:ferredoxin (flavodoxin) oxidoreductase | |
| 13 | K00163 pyruvate dehydrogenase E1 component [EC:1.2.4.1] | LT988_22730 | aceE; pyruvate dehydrogenase (acetyl-transferring), homodimeric type |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_05285 | dihydrolipoyl dehydrogenase |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_13640 | dihydrolipoyl dehydrogenase |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_22720 | lpdA; dihydrolipoyl dehydrogenase |
| 15 | K00627 pyruvate dehydrogenase E2 component (dihydrolipoamide acetyltransferase) [EC:2.3.1.12] | LT988_22725 | aceF; dihydrolipoyllysine-residue acetyltransferase |
| № | Function Number, Enzyme Annotation, and Number (According to KEGG Orthology Database) | Gene ID | Gene Name and Annotation Presented in GenBank |
|---|---|---|---|
| 1 | K00845 glucokinase [EC:2.7.1.2] | LT988_04670 | glk; glucokinase |
| 2 | K01810 glucose-6-phosphate isomerase [EC:5.3.1.9] | LT988_04445 | pgi; glucose-6-phosphate isomerase |
| 2 | K01810 glucose-6-phosphate isomerase [EC:5.3.1.9] | LT988_25180 | pgi; glucose-6-phosphate isomerase |
| 3 | K21071 ATP-dependent phosphofructokinase/diphosphate-dependent phosphofructokinase [EC:2.7.1.11; 2.7.1.90] | LT988_14085 | 6-phosphofructokinase |
| 4 | K01623 fructose-bisphosphate aldolase, class I [EC:4.1.2.13] | LT988_25065 | fructose-bisphosphate aldolase class I |
| 4 | K01624 fructose-bisphosphate aldolase, class II [EC:4.1.2.13] | LT988_08070 | fba; fructose-bisphosphate aldolase class II |
| 5 | K01803 triose-phosphate isomerase (TIM) [EC:5.3.1.1] | LT988_02660 | tpiA; triose-phosphate isomerase |
| 6 | K00134 glyceraldehyde 3-phosphate dehydrogenase (phosphorylating) [EC:1.2.1.12] | LT988_08050 | gap; type I glyceraldehyde-3-phosphate dehydrogenase |
| 7 | K00927 phosphoglycerate kinase [EC:2.7.2.3] | LT988_08060 | phosphoglycerate kinase |
| 8 | K15634 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase [EC:5.4.2.11] | LT988_13795 | histidine phosphatase family protein |
| 8 | K15633 2,3-bisphosphoglycerate-independent phosphoglycerate mutase [EC:5.4.2.12] | LT988_06575 | gpmI; 2,3-bisphosphoglycerate-independent phosphoglycerate mutase |
| 9 | K01689 enolase [EC:4.2.1.11] | LT988_19115 | eno; phosphopyruvate hydratase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_01025 | pyk; pyruvate kinase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_08065 | pyk; pyruvate kinase |
| 10 | K00873 pyruvate kinase [EC:2.7.1.40] | LT988_22860 | pyruvate kinase |
| 11 | K03841 fructose-1,6-bisphosphatase I [EC:3.1.3.11] | LT988_07425 | class 1 fructose-bisphosphatase |
| 12 | K00172 pyruvate-ferredoxin oxidoreductase gamma subunit [EC:1.2.7.1] | LT988_13550 | 2-oxoacid:acceptor oxidoreductase family protein |
| K00169 pyruvate-ferredoxin oxidoreductase alpha subunit [EC:1.2.7.1] | LT988_13555 | hypothetical protein | |
| K00170 pyruvate-ferredoxin oxidoreductase beta subunit [EC:1.2.7.1] | LT988_13560 | thiamine pyrophosphate-dependent enzyme | |
| 12 | K03737 pyruvate-ferredoxin/flavodoxin oxidoreductase [EC:1.2.7.1 1.2.7.-] | LT988_24065 | 2-oxoacid:acceptor oxidoreductase family protein |
| K03737 pyruvate-ferredoxin/flavodoxin oxidoreductase [EC:1.2.7.1 1.2.7.-] | LT988_24180 | nifJ; pyruvate:ferredoxin (flavodoxin) oxidoreductase | |
| 13 | K00163 pyruvate dehydrogenase E1 component [EC:1.2.4.1] | LT988_22730 | aceE; pyruvate dehydrogenase (acetyl-transferring), homodimeric type |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_05285 | dihydrolipoyl dehydrogenase |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_13640 | dihydrolipoyl dehydrogenase |
| 14 | K00382 dihydrolipoamide dehydrogenase [EC:1.8.1.4] | LT988_22720 | lpdA; dihydrolipoyl dehydrogenase |
| 15 | K00627 pyruvate dehydrogenase E2 component (dihydrolipoamide acetyltransferase) [EC:2.3.1.12] | LT988_22725 | aceF; dihydrolipoyllysine-residue acetyltransferase |
| № | Function Number, Enzyme Annotation, and Number (According to KEGG Orthology Database) | Gene ID | Gene Name and Annotation Presented in GenBank |
|---|---|---|---|
| 1 | K17222 L-cysteine S-thiosulfotransferase [EC:2.8.5.2] | LT988_12625 | soxA; sulfur oxidation c-type cytochrome SoxA |
| 1 | K17222 L-cysteine S-thiosulfotransferase [EC:2.8.5.2] | LT988_24245 | soxA; sulfur oxidation c-type cytochrome SoxA |
| 2 | K17223 L-cysteine S-thiosulfotransferase [EC:2.8.5.2] | LT988_12630 | soxX; sulfur oxidation c-type cytochrome SoxX |
| 2 | K17223 L-cysteine S-thiosulfotransferase [EC:2.8.5.2] | LT988_24240 | soxX; sulfur oxidation c-type cytochrome SoxX |
| 3 | K17224 S-sulfosulfanyl-L-cysteine sulfohydrolase [EC:3.1.6.20] | LT988_24235 | soxB; thiosulfohydrolase SoxB |
| 4 | K17226 sulfur-oxidizing protein SoxY | LT988_05335 | SoxY; thiosulfate oxidation carrier protein SoxY |
| 4 | K17226 sulfur-oxidizing protein SoxY | LT988_16035 | SoxY; thiosulfate oxidation carrier protein SoxY |
| 4 | K17226 sulfur-oxidizing protein SoxY | LT988_21280 | hypothetical protein |
| 5 | K17227 sulfur-oxidizing protein SoxZ | LT988_05330 | soxZ; thiosulfate oxidation carrier complex protein SoxZ |
| 5 | K17227 sulfur-oxidizing protein SoxZ | LT988_16040 | soxZ; thiosulfate oxidation carrier complex protein SoxZ |
| 5 | K17227 sulfur-oxidizing protein SoxZ | LT988_21285 | soxZ; thiosulfate oxidation carrier complex protein SoxZ |
| 6 | K17218 sulfide:quinone oxidoreductase [EC:1.8.5.4] | LT988_02950 | FAD-dependent oxidoreductase |
| 6 | K17218 sulfide:quinone oxidoreductase [EC:1.8.5.4] | LT988_15980 | FAD-dependent oxidoreductase |
| 7 | K17230 cytochrome subunit of sulfide dehydrogenase | LT988_00620 | cytochrome c4 |
| 8 | K17229 sulfide dehydrogenase [flavocytochrome c] flavoprotein chain [EC:1.8.2.3] | LT988_00625 | FCSD flavin-binding domain-containing protein |
| № | ID and Name of Alc. vinosum DSM 180T Gene | Function Number, Enzyme Annotation, and Number of T. bogorovii BBS (According to KEGG Orthology Database) | Gene ID of T. bogorovii BBS | Gene name and Annotation of T. bogorovii BBS Presented in GenBank |
|---|---|---|---|---|
| 1 | Alvin_1265 dsrS | no KO assigned | LT988_06595 | sulfur reduction protein DsrS |
| 2 | Alvin_1264 dsrR | K13628 iron–sulfur cluster assembly protein | LT988_06600 | iron–sulfur cluster assembly accessory protein |
| 3 | Alvin_1263 dsrN | K02224 cobyrinic acid a,c-diamide synthase [EC:6.3.5.9 6.3.5.11] | LT988_06605 | cobyrinate a,c-diamide synthase |
| 4 | Alvin_1262 dsrP | no KO assigned | LT988_06610 | nrfD; polysulfide reductase NrfD |
| 5 | Alvin_1261 dsrO | no KO assigned | LT988_06615 | 4Fe-4S dicluster domain-containing protein |
| 6 | Alvin_1260 dsrJ | no KO assigned | LT988_06620 | sulfur reduction protein DsrJ |
| 7 | Alvin_1259 dsrL | no KO assigned | LT988_06625 | NAD(P)-binding protein |
| 8 | Alvin_1258 dsrK | no KO assigned | LT988_06630 | (Fe-S)-binding protein |
| 9 | Alvin_1257 dsrM | K00374 nitrate reductase gamma subunit [EC:1.7.5.1 1.7.99.-] | LT988_06635 | respiratory nitrate reductase subunit gamma |
| 10 | Alvin_1256 dsrC | K23077 dissimilatory sulfite reductase related protein | LT988_06640 | TusE/DsrC/DsvC family sulfur relay protein |
| 11 | Alvin_1255 dsrH | K07237 tRNA 2-thiouridine synthesizing protein B | LT988_06645 | tusB; sulfurtransferase complex subunit TusB |
| 12 | Alvin_1254 dsrF | K07236 tRNA 2-thiouridine synthesizing protein C | LT988_06650 | tusC; sulfurtransferase complex subunit TusC |
| 13 | Alvin_1253 dsrE | K07235 tRNA 2-thiouridine synthesizing protein D [EC:2.8.1.-] | LT988_06655 | tusD; sulfurtransferase complex subunit TusD |
| 14 | Alvin_1252 dsrB | K11181 dissimilatory sulfite reductase beta subunit [EC:1.8.99.5] | LT988_06660 | dsrB; dissimilatory-type sulfite reductase subunit beta |
| 15 | Alvin_1251 dsrA | K11180 dissimilatory sulfite reductase alpha subunit [EC:1.8.99.5] | LT988_06665 | dsrA; dissimilatory-type sulfite reductase subunit alpha |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petushkova, E.; Khasimov, M.; Mayorova, E.; Delegan, Y.; Frantsuzova, E.; Bogun, A.; Galkina, E.; Tsygankov, A. The Complete Genome of a Novel Typical Species Thiocapsa bogorovii and Analysis of Its Central Metabolic Pathways. Microorganisms 2024, 12, 391. https://doi.org/10.3390/microorganisms12020391
Petushkova E, Khasimov M, Mayorova E, Delegan Y, Frantsuzova E, Bogun A, Galkina E, Tsygankov A. The Complete Genome of a Novel Typical Species Thiocapsa bogorovii and Analysis of Its Central Metabolic Pathways. Microorganisms. 2024; 12(2):391. https://doi.org/10.3390/microorganisms12020391
Chicago/Turabian StylePetushkova, Ekaterina, Makhmadyusuf Khasimov, Ekaterina Mayorova, Yanina Delegan, Ekaterina Frantsuzova, Alexander Bogun, Elena Galkina, and Anatoly Tsygankov. 2024. "The Complete Genome of a Novel Typical Species Thiocapsa bogorovii and Analysis of Its Central Metabolic Pathways" Microorganisms 12, no. 2: 391. https://doi.org/10.3390/microorganisms12020391
APA StylePetushkova, E., Khasimov, M., Mayorova, E., Delegan, Y., Frantsuzova, E., Bogun, A., Galkina, E., & Tsygankov, A. (2024). The Complete Genome of a Novel Typical Species Thiocapsa bogorovii and Analysis of Its Central Metabolic Pathways. Microorganisms, 12(2), 391. https://doi.org/10.3390/microorganisms12020391