

Seasonality and Genotype Diversity of Human Rhinoviruses during an Eight-Year Period in Slovenia

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Period
2.2. Sample Collection and Data Source
2.3. Molecular Identification of Rhinoviruses
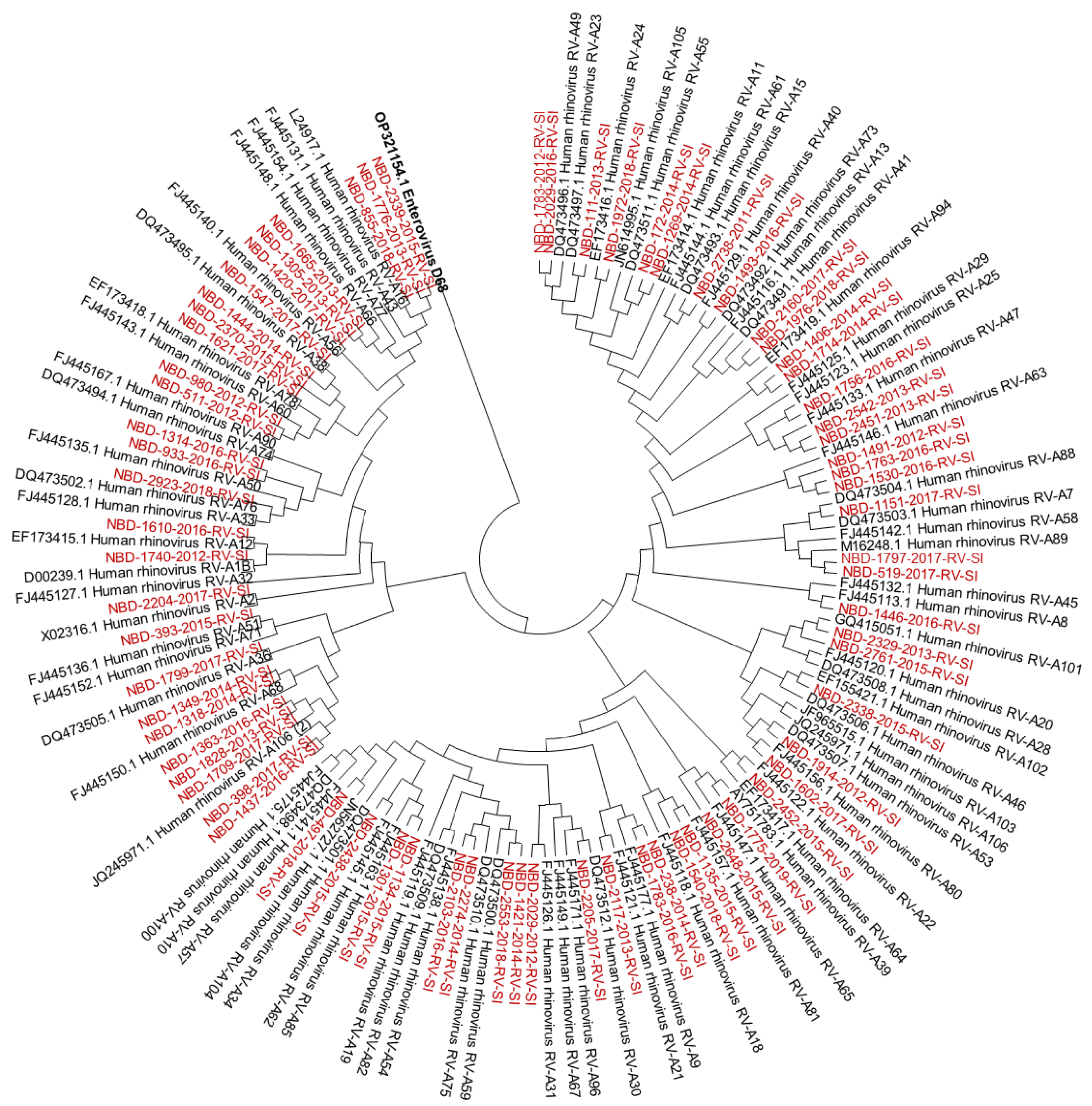
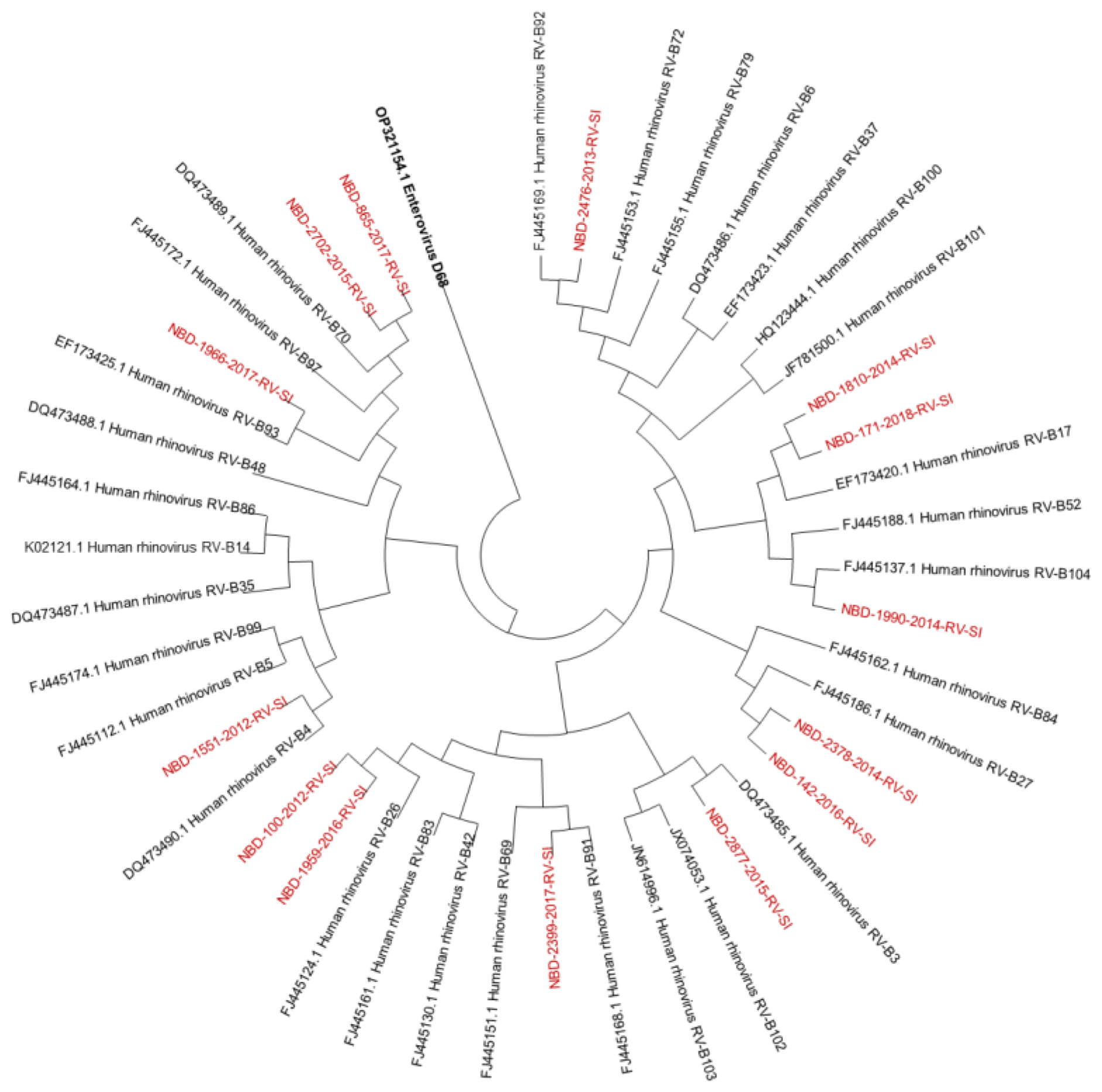
2.4. Genotyping of Rhinoviruses
2.5. Statistical Analyses
2.6. Ethical Considerations
3. Results
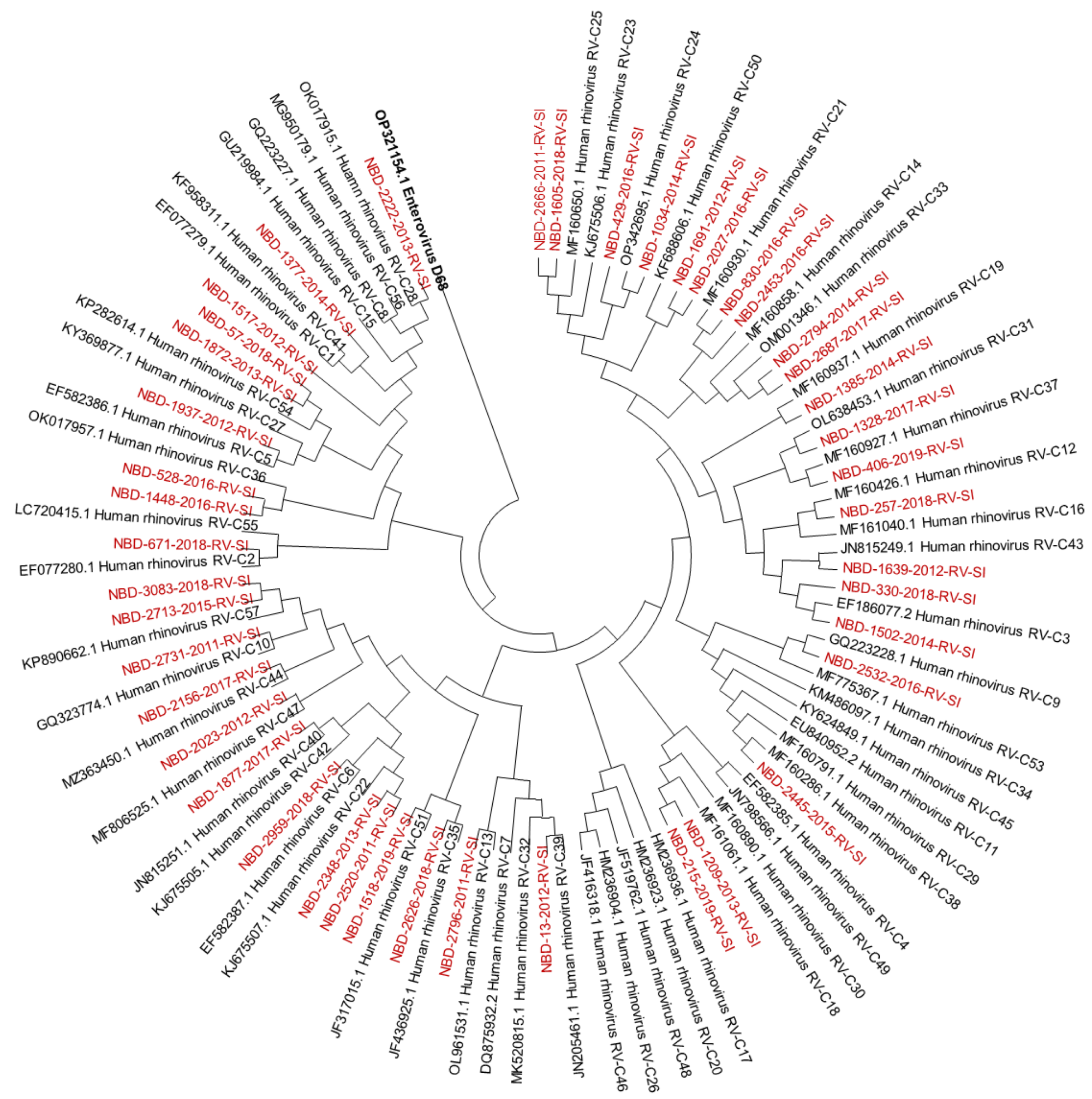
3.1. Molecular Identification and Genotyping of Rhinoviruses
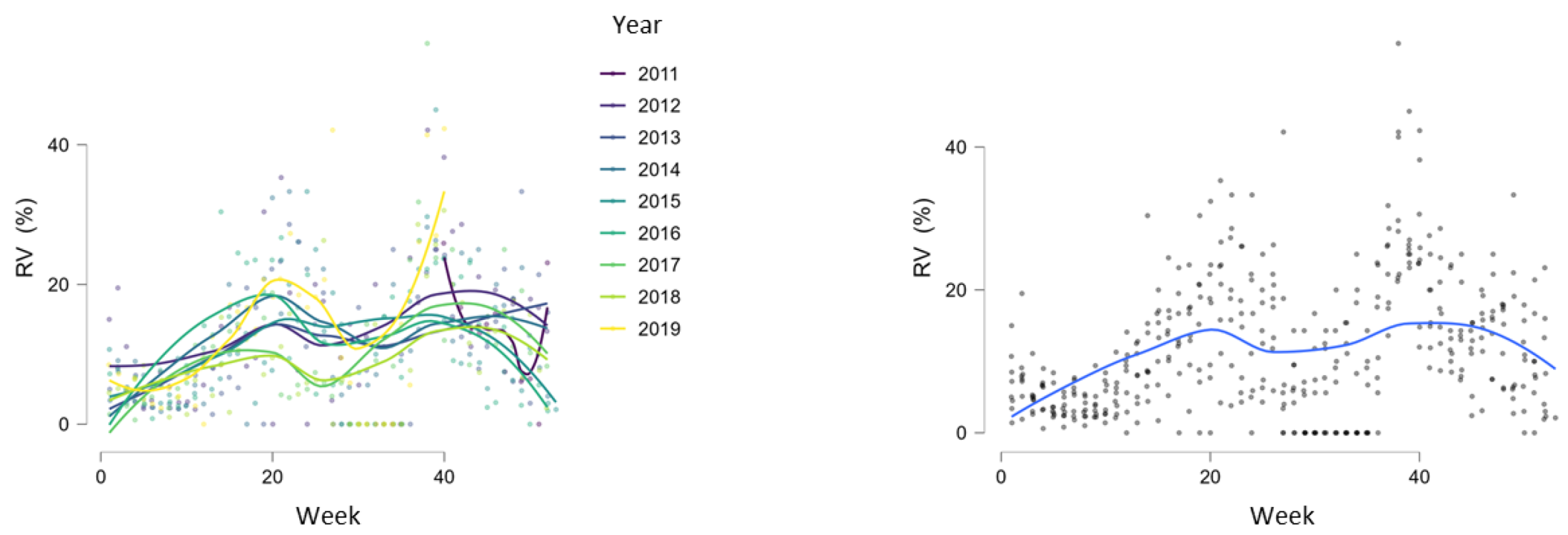
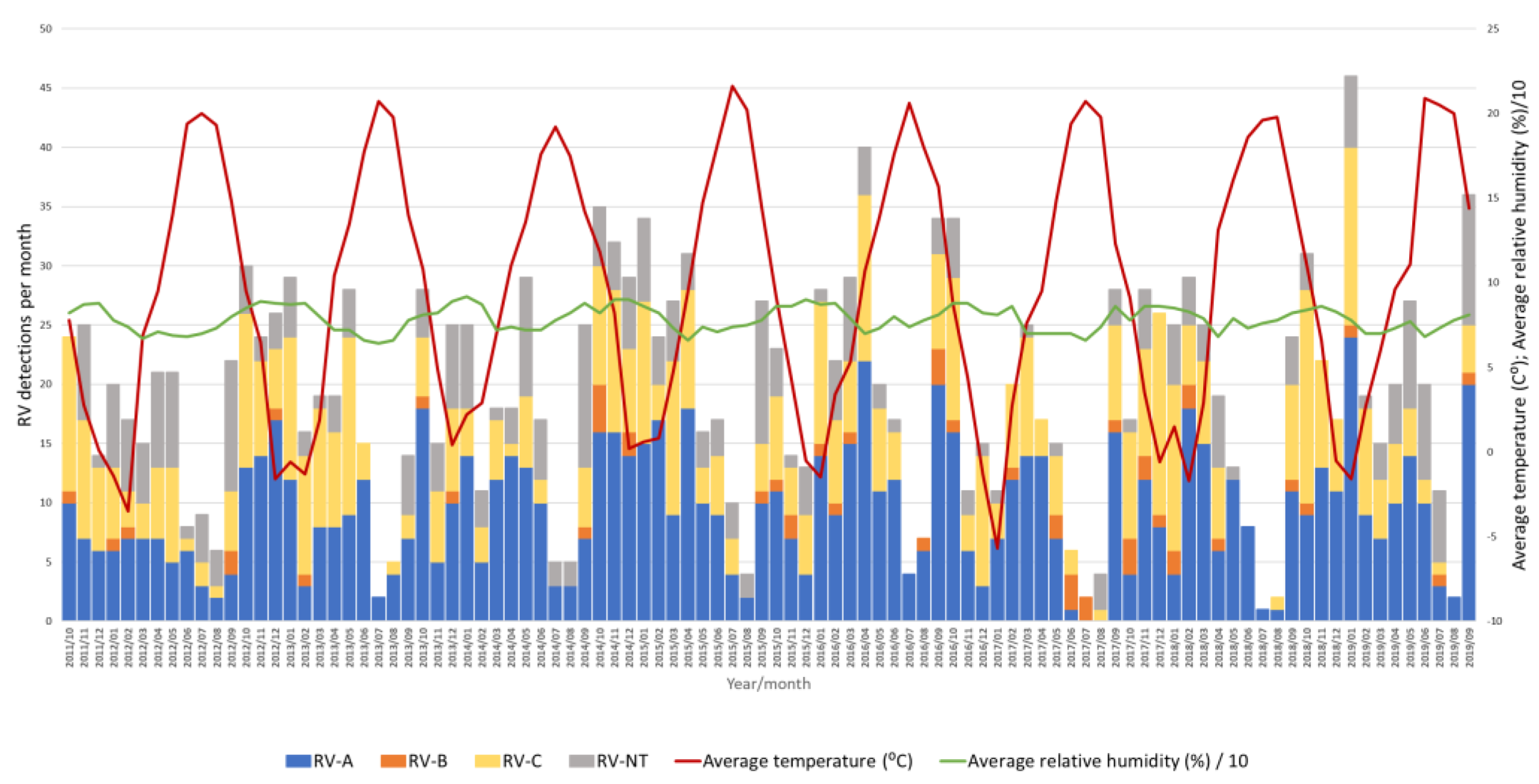
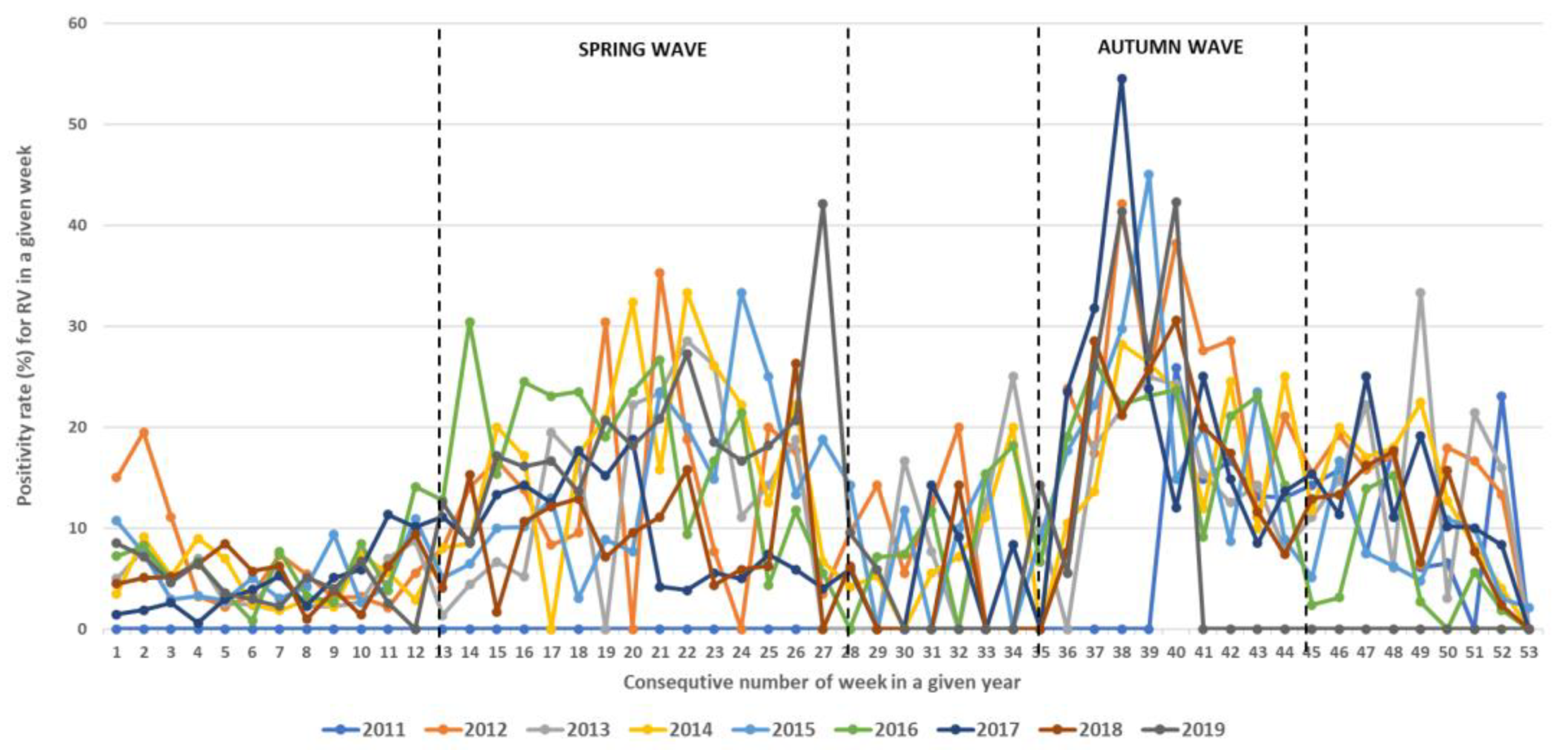
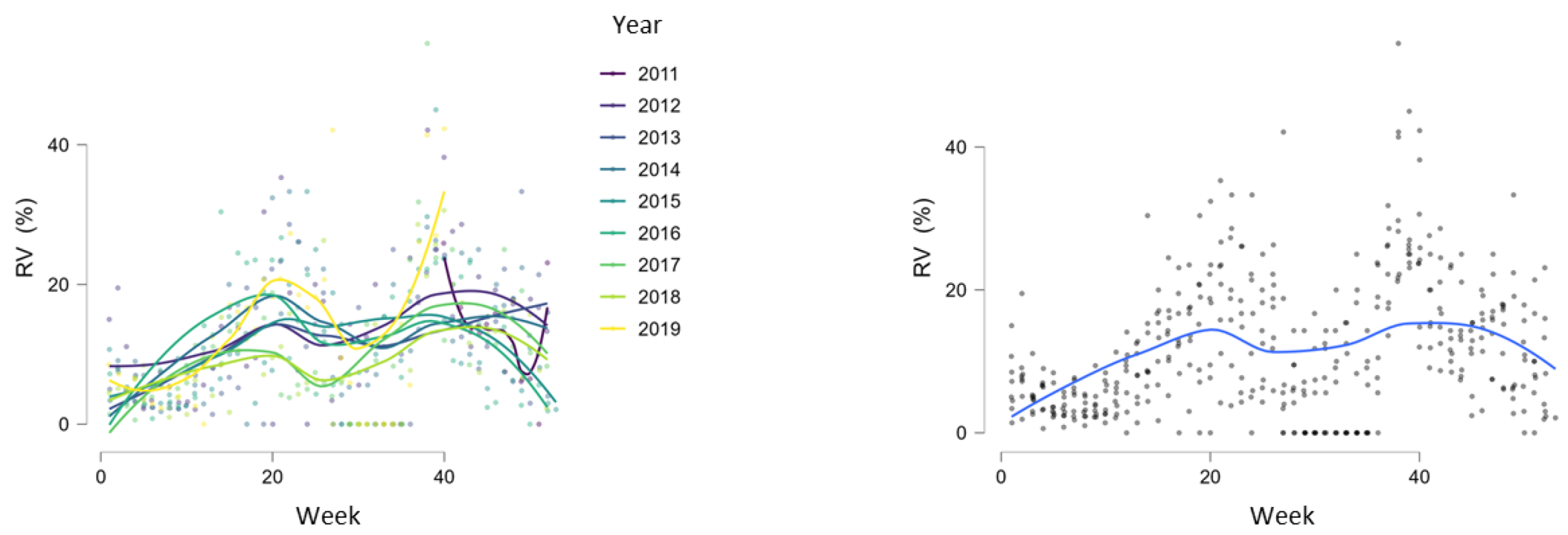
3.2. Seasonality of Rhinoviruses
3.3. Impact of Meteorological Factors on Rhinovirus Circulation
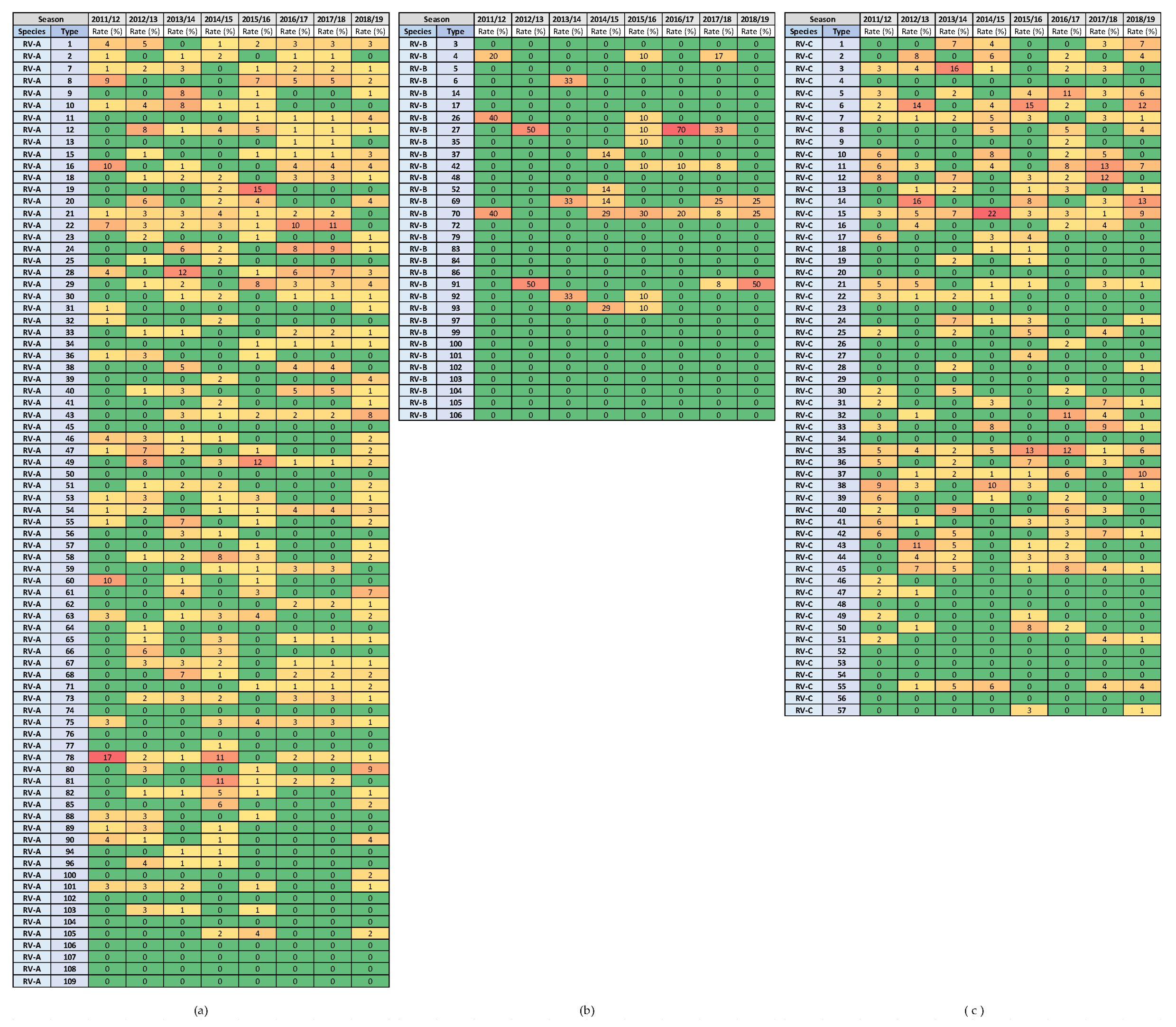
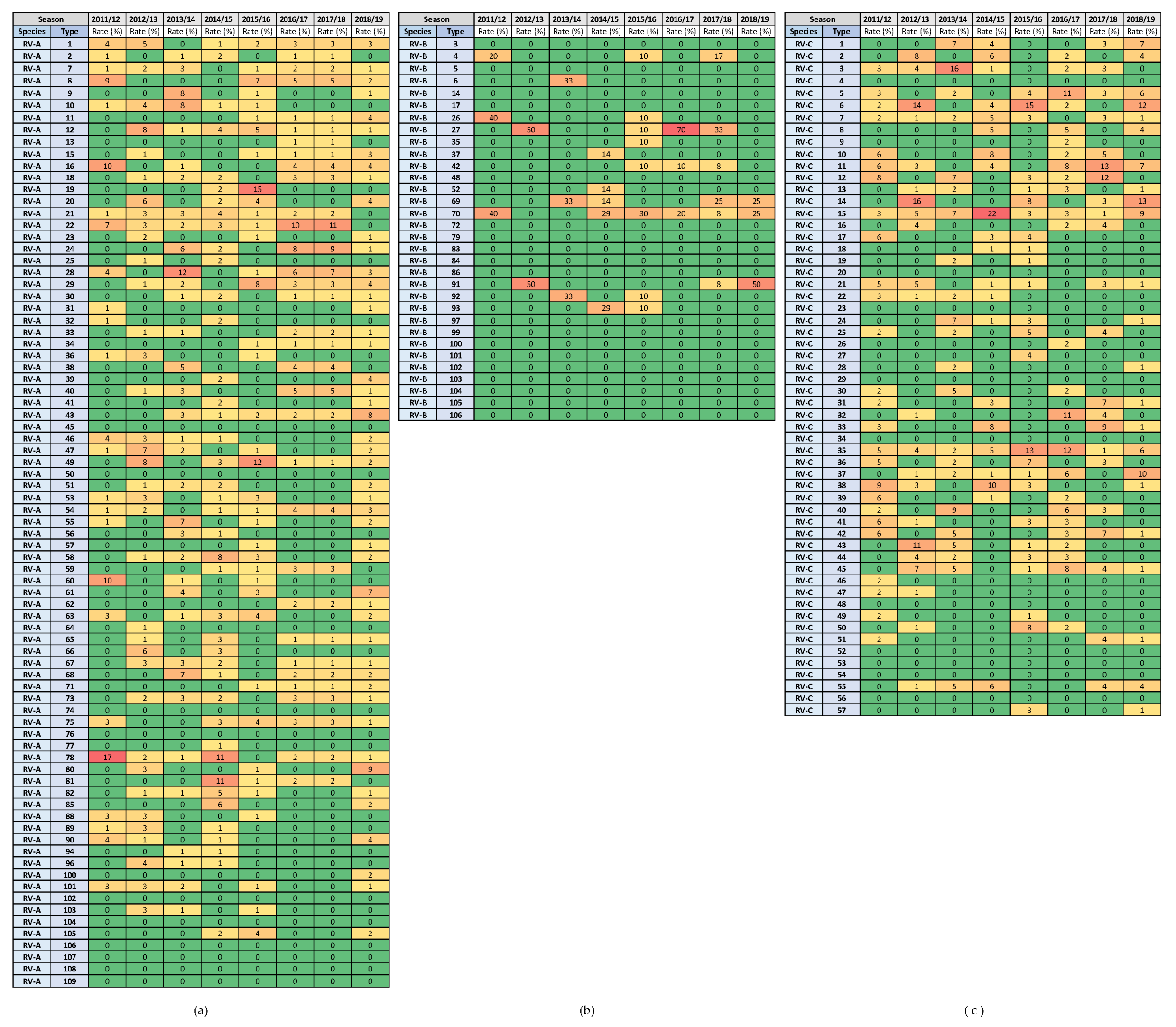
3.4. Variability in Rhinovirus Species and Types between and within Seasons
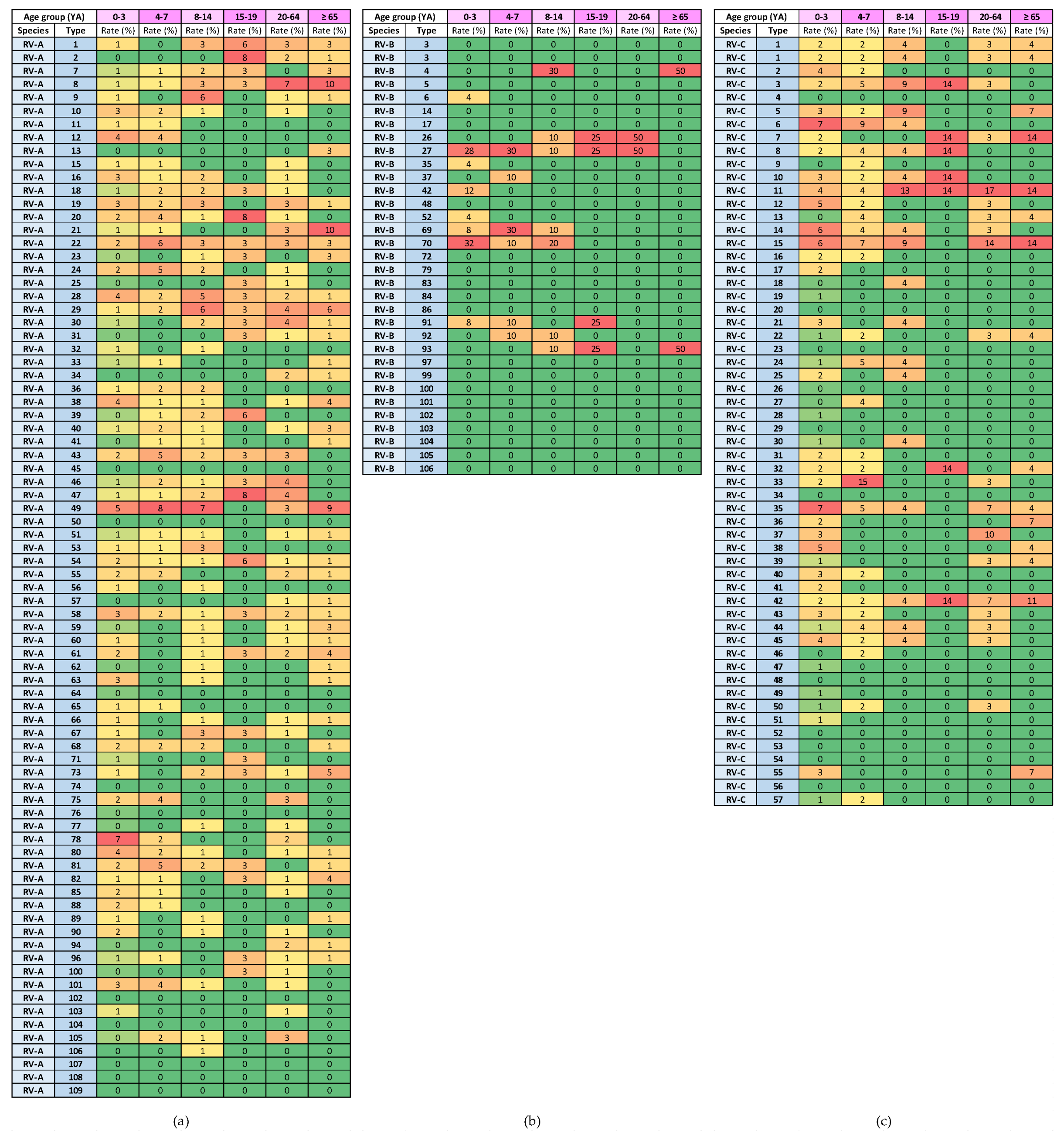
3.5. Variability in Rhinovirus Species and Types by Age Group
3.6. Variability in Rhinovirus Species and Types in Patients with MAARIs and SARIs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahn, J.G.; Kim, D.S.; Kim, K.H. Clinical characteristics and cytokine profiles of children with acute lower respiratory tract infections caused by human rhinovirus. PLoS ONE 2018, 13, e0198624. [Google Scholar] [CrossRef]
- Lee, W.M.; Lemanske, R.F., Jr.; Evans, M.D.; Vang, F.; Pappas, T.; Gangnon, R.; Jackson, D.J.; Gern, J.E. Human rhinovirus species and season of infection determine illness severity. Am. J. Respir. Crit. Care Med. 2012, 186, 886–891. [Google Scholar] [CrossRef]
- Georgieva, I.; Stoyanova, A.; Angelova, S.; Korsun, N.; Stoitsova, S.; Nikolaeva-Glomb, L. Rhinovirus Genotypes Circulating in Bulgaria, 2018–2021. Viruses 2023, 15, 1608. [Google Scholar] [CrossRef] [PubMed]
- Esneau, C.; Duff, A.C.; Bartlett, N.W. Understanding Rhinovirus Circulation and Impact on Illness. Viruses 2022, 14, 141. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Lamson, D.M.; St George, K.; Walsh, T.J. Human rhinoviruses. Clin. Microbiol. Rev. 2013, 26, 135–162. [Google Scholar] [CrossRef]
- Golke, P.; Hönemann, M.; Bergs, S.; Liebert, U.G. Human Rhinoviruses in Adult Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance. Viruses 2021, 13, 2027. [Google Scholar] [CrossRef] [PubMed]
- ICTV International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/picornavirales/w/picornaviridae/681/genus-enterovirus (accessed on 18 September 2023).
- Simmonds, P.; Gorbalenya, A.E.; Harvala, H.; Hovi, T.; Knowles, N.J.; Lindberg, A.M.; Oberste, M.S.; Palmenberg, A.C.; Reuter, G.; Skern, T.; et al. Recommendations for the nomenclature of enteroviruses and rhinoviruses. Arch. Virol. 2020, 165, 793–797. [Google Scholar] [CrossRef]
- Royston, L.; Tapparel, C. Rhinoviruses and Respiratory Enteroviruses: Not as Simple as ABC. Viruses 2016, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, C.L.; Knowles, N.J.; Simmonds, P. Proposals for the classification of human rhinovirus species A, B and C into genotypically assigned types. J. Gen. Virol. 2013, 94 Pt 8, 1791–1806. [Google Scholar] [CrossRef]
- Wisdom, A.; Leitch, E.C.; Gaunt, E.; Harvala, H.; Simmonds, P. Screening respiratory samples for detection of human rhinoviruses (HRVs) and enteroviruses: Comprehensive VP4-VP2 typing reveals high incidence and genetic diversity of HRV species C. J. Clin. Microbiol. 2009, 47, 3958–3967. [Google Scholar] [CrossRef]
- Giardina, F.A.M.; Piralla, A.; Ferrari, G.; Zavaglio, F.; Cassaniti, I.; Baldanti, F. Molecular Epidemiology of Rhinovirus/Enterovirus and Their Role on Cause Severe and Prolonged Infection in Hospitalized Patients. Microorganisms 2022, 10, 755. [Google Scholar] [CrossRef]
- To, K.K.W.; Yip, C.C.Y.; Yuen, K.Y. Rhinovirus-From bench to bedside. J. Formos. Med. Assoc. 2017, 116, 496–504. [Google Scholar] [CrossRef]
- Messacar, K.; Robinson, C.C.; Bagdure, D.; Curtis, D.J.; Glodé, M.P.; Dominguez, S.R. Rhino/enteroviruses in hospitalized children: A comparison to influenza viruses. J. Clin. Virol. 2013, 56, 41–45. [Google Scholar] [CrossRef]
- Ratnamohan, V.M.; Zeng, F.; Donovan, L.; MacIntyre, C.R.; Kok, J.; Dwyer, D.E. Phylogenetic analysis of human rhinoviruses collected over four successive years in Sydney, Australia. Influenza Other Respir. Viruses 2016, 10, 493–503. [Google Scholar] [CrossRef]
- Garcia, J.; Espejo, V.; Nelson, M.; Sovero, M.; Villaran, M.V.; Gomez, J.; Barrantes, M.; Sanchez, F.; Comach, G.; Arango, A.E.; et al. Human rhinoviruses and enteroviruses in influenza-like illness in Latin America. Virol. J. 2013, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Huang, L.; Wang, M.; Wang, Y.; Ji, W.; Zhu, C.; Chen, Z. Clinical and epidemiological profiles including meteorological factors of low respiratory tract infection due to human rhinovirus in hospitalized children. Ital. J. Pediatr. 2017, 43, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shen, J.; Wu, B.; Liu, G.; Lu, R.; Tan, W. Genotypic Diversity and Epidemiology of Human Rhinovirus Among Children with Severe Acute Respiratory Tract Infection in Shanghai, 2013–2015. Front. Microbiol. 2018, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Socan, M.; Prosenc, K. Integration of COVID-19 into the Sentinel Influenza and Other Acute Respiratory Infections Network. Sci. J. Public Health 2022, 10, 207–213. [Google Scholar] [CrossRef]
- Slovenian Environmental Agency, National Meteorological Services. Available online: https://meteo.arso.gov.si/ (accessed on 18 September 2023).
- Chen, Y.; Cui, D.; Zheng, S.; Yang, S.; Tong, J.; Yang, D.; Fan, J.; Zhang, J.; Lou, B.; Li, X.; et al. Simultaneous detection of influenza A, influenza B, and respiratory syncytial viruses and subtyping of influenza A H3N2 virus and H1N1 (2009) virus by multiplex real-time PCR. J. Clin. Microbiol. 2011, 49, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- De Vos, N.; Vankeerberghen, A.; Vaeyens, F.; Van Vaerenbergh, K.; Boel, A.; De Beenhouwer, H. Simultaneous detection of human bocavirus and adenovirus by multiplex real-time PCR in a Belgian paediatric population. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2009, 28, 1305–1310. [Google Scholar] [CrossRef]
- Dierssen, U.; Rehren, F.; Henke-Gendo, C.; Harste, G.; Heim, A. Rapid routine detection of enterovirus RNA in cerebrospinal fluid by a one-step real-time RT-PCR assay. J. Clin. Virol. 2008, 42, 58–64. [Google Scholar] [CrossRef]
- Auburn, H.; Zuckerman, M.; Broughton, S.; Greenough, A.; Smith, M. Detection of nine respiratory RNA viruses using three multiplex RT-PCR assays incorporating a novel RNA internal control transcript. J. Virol. Methods 2011, 176, 9–13. [Google Scholar] [CrossRef]
- Klemenc, J.; Asad Ali, S.; Johnson, M.; Tollefson, S.J.; Talbot, H.K.; Hartert, T.V.; Edwards, K.M.; Williams, J.V. Real-time reverse transcriptase PCR assay for improved detection of human metapneumovirus. J. Clin. Virol. 2012, 54, 371–375. [Google Scholar] [CrossRef]
- Bezerra, P.G.; Britto, M.C.; Correia, J.B.; Duarte, M.D.C.; Fonceca, A.M.; Rose, K.; Hopkins, M.J.; Cuevas, L.E.; McNamara, P.S. Viral and atypical bacterial detection in acute respiratory infection in children under five years. PLoS ONE 2011, 6, e18928. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, B.; Swamy, M.A.; Reddy, P.V.; Kumar, N.; Tiwari, J.K. Evaluation of custom multiplex real-time RT-PCR in comparison to fast-track diagnostics respiratory 21 pathogens kit for detection of multiple respiratory viruses. Virol. J. 2016, 13, 91. [Google Scholar] [CrossRef]
- Rossen, J. Parainfluenza Virus Research Using a Multiplex Real-Time RT_PCR Method and ViiA 7 Real-Time PCR System. 2010. AB—Innovatove Research Brochure. Available online: https://tools.thermofisher.cn/content/sfs/brochures/cms_088565.pdf (accessed on 2 February 2024).
- Hall, T.A. A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98 NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- NCBI-National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/nuccore (accessed on 18 September 2023).
- Nix, W.A.; Oberste, M.S.; Pallansch, M.A. Sensitive, seminested PCR amplification of VP1 sequences for direct identification of all enterovirus serotypes from original clinical specimens. J. Clin. Microbiol. 2006, 44, 2698–2704. [Google Scholar] [CrossRef] [PubMed]
- Mubareka, S.; Louie, L.; Wong, H.; Granados, A.; Chong, S.; Luinstra, K.; Petrich, A.; Smieja, M.; Vearncombe, M.; Mahony, J.; et al. Co-circulation of multiple genotypes of human rhinovirus during a large outbreak of respiratory illness in a veterans’ long-term care home. J. Clin. Virol. 2013, 58, 455–460. [Google Scholar] [CrossRef]
- MEGA—Molecular Evolutionary Genteics Analysis. Available online: https://www.megasoftware.net/ (accessed on 18 September 2023).
- Hall, B.G. Phylogenetic Trees Made Easy, 4th ed.Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- JASP (Version 0.17.3). Available online: https://jasp-stats.org/ (accessed on 18 September 2023).
- Marcone, D.N.; Culasso, A.; Carballal, G.; Campos, R.; Echavarría, M. Genetic diversity and clinical impact of human rhinoviruses in hospitalized and outpatient children with acute respiratory infection, Argentina. J. Clin. Virol. 2014, 61, 558–564. [Google Scholar] [CrossRef]
- van der Linden, L.; Bruning, A.H.; Thomas, X.V.; Minnaar, R.P.; Rebers, S.P.; Schinkel, J.; de Jong, M.D.; Pajkrt, D.; Wolthers, K.C. A molecular epidemiological perspective of rhinovirus types circulating in Amsterdam from 2007 to 2012. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2016, 22, 1002.e9–1002.e14. [Google Scholar] [CrossRef]
- Morobe, J.M.; Nyiro, J.U.; Brand, S.; Kamau, E.; Gicheru, E.; Eyase, F.; Otieno, G.P.; Munywoki, P.K.; Agoti, C.N.; Nokes, D.J. Human rhinovirus spatial-temporal epidemiology in rural coastal Kenya, 2015–2016, observed through outpatient surveillance. Welcome Open Res. 2019, 3, 128. [Google Scholar] [CrossRef]
- Richter, J.; Nikolaou, E.; Panayiotou, C.; Tryfonos, C.; Koliou, M.; Christodoulou, C. Molecular epidemiology of rhinoviruses in Cyprus over three consecutive seasons. Epidemiol. Infect. 2015, 143, 1876–1883. [Google Scholar] [CrossRef]
- Lau, S.K.; Yip, C.C.; Lin, A.W.; Lee, R.A.; So, L.Y.; Lau, Y.L.; Chan, K.H.; Woo, P.C.; Yuen, K.Y. Clinical and molecular epidemiology of human rhinovirus C in children and adults in Hong Kong reveals a possible distinct human rhinovirus C subgroup. J. Infect. Dis. 2009, 200, 1096–1103. [Google Scholar] [CrossRef]
- Naughtin, M.; Sareth, R.; Sentilhes, A.C.; Vong, S.; Joffret, M.L.; Cornillot, E.; Deubel, V.; Delpeyroux, F.; Frutos, R.; Buchy, P. Genetic diversity of human rhinoviruses in Cambodia during a three-year period reveals novel genetic types. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 35, 42–49. [Google Scholar] [CrossRef]
- Simmonds, P.; McIntyre, C.; Savolainen-Kopra, C.; Tapparel, C.; Mackay, I.M.; Hovi, T. Proposals for the classification of human rhinovirus species C into genotypically assigned types. J. Gen. Virol. 2010, 91 Pt 10, 2409–2419. [Google Scholar] [CrossRef]
- Megremis, S.; Demetriou, P.; Makrinioti, H.; Manoussaki, A.E.; Papadopoulos, N.G. The genomic signature of human rhinoviruses A, B and C. PLoS ONE 2012, 7, e44557. [Google Scholar] [CrossRef]
- Pierangeli, A.; Ciccozzi, M.; Chiavelli, S.; Concato, C.; Giovanetti, M.; Cella, E.; Spano, L.; Scagnolari, C.; Moretti, C.; Papoff, P.; et al. Molecular epidemiology and genetic diversity of human rhinovirus affecting hospitalized children in Rome. Med. Microbiol. Immunol. 2013, 202, 303–311. [Google Scholar] [CrossRef]
- Cuevas, M.T.; Molinero, M.; Pozo, F.; Calvo, C.; García-García, M.L.; Reyes, N.; Ledesma, J.; Casas, I. Spread of different rhinovirus B genotypes in hospitalized children in Spain. Influenza Other Respir. Viruses 2013, 7, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Viral Interference between Respiratory Viruses. Emerg. Infect. Dis. 2022, 28, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Zlateva, K.T.; van Rijn, A.L.; Simmonds, P.; Coenjaerts, F.E.J.; van Loon, A.M.; Verheij, T.J.M.; de Vries, J.J.C.; Little, P.; Butler, C.C.; van Zwet, E.W.; et al. Molecular epidemiology and clinical impact of rhinovirus infections in adults during three epidemic seasons in 11 European countries (2007–2010). Thorax 2020, 75, 882–890. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 AG (2 YA) | 1 (0–3) | 2 (4–7) | 3 (8–14) | 4 (15–19) | 5 (20–64) | 6 (≥65) | ||||||
| 3 n | 4 Rate (%) | n | Rate (%) | n | Rate (%) | n | Rate (%) | n | Rate (%) | n | Rate (%) | |
| Male | 686 | 59.6 | 115 | 57.8 | 76 | 51.7 | 23 | 39.7 | 54 | 39.1 | 65 | 52.4 |
| Female | 465 | 40.4 | 84 | 42.2 | 71 | 48.3 | 35 | 60.3 | 84 | 60.9 | 59 | 47.6 |
| SARI | 1079 | 93.7 | 187 | 94.0 | 116 | 78.9 | 29 | 50.0 | 91 | 65.9 | 121 | 97.6 |
| MAARI | 72 | 6.3 | 12 | 6.0 | 31 | 21.1 | 29 | 50.0 | 47 | 34.1 | 3 | 2.4 |
| M (%) | ||||
|---|---|---|---|---|
| Seasonal Waves | RV (All Species) | RV-A | RV-B | RV-C |
| Spring waves | 15.8 | 21.9 | 1.6 | 14.5 |
| Autumn waves | 15.4 | 19.1 | 7.8 | 14.3 |
| p-value | 0.706 | 0.226 | 0.002 | 0.609 |
| M (%) | ||||
|---|---|---|---|---|
| 1 AG (2 YA) | RV (All Species) | RV-A | RV-B | RV-C |
| 1 (0–3) | 28.6 | 32.5 | 6.7 | 35.5 |
| 2 (4–7) | 7.9 | 9.1 | 3.1 | 8.9 |
| 3 (8–14) | 7.8 | 11.4 | 3.9 | 4.9 |
| 4 (15–19) | 3.4 | 5.5 | 1.6 | 1.6 |
| 5 (20–64) | 6.8 | 11.1 | 0.8 | 5.9 |
| 6 (≥65) | 6.2 | 9.6 | 0.8 | 4.6 |
| Severity of Infection | M (%) | |||
|---|---|---|---|---|
| RV (All Species) | RV-A | RV-B | RV-C | |
| SARI | 35.4 | 42.1 | 11.7 | 39.5 |
| MAARI | 9.1 | 12.5 | 2.7 | 7.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berginc, N.; Sočan, M.; Prosenc Trilar, K.; Petrovec, M. Seasonality and Genotype Diversity of Human Rhinoviruses during an Eight-Year Period in Slovenia. Microorganisms 2024, 12, 341. https://doi.org/10.3390/microorganisms12020341
Berginc N, Sočan M, Prosenc Trilar K, Petrovec M. Seasonality and Genotype Diversity of Human Rhinoviruses during an Eight-Year Period in Slovenia. Microorganisms. 2024; 12(2):341. https://doi.org/10.3390/microorganisms12020341
Chicago/Turabian StyleBerginc, Nataša, Maja Sočan, Katarina Prosenc Trilar, and Miroslav Petrovec. 2024. "Seasonality and Genotype Diversity of Human Rhinoviruses during an Eight-Year Period in Slovenia" Microorganisms 12, no. 2: 341. https://doi.org/10.3390/microorganisms12020341
APA StyleBerginc, N., Sočan, M., Prosenc Trilar, K., & Petrovec, M. (2024). Seasonality and Genotype Diversity of Human Rhinoviruses during an Eight-Year Period in Slovenia. Microorganisms, 12(2), 341. https://doi.org/10.3390/microorganisms12020341