
A Comprehensive Study of the Effects by Sequence Truncation within Inverted Terminal Repeats (ITRs) on the Productivity, Genome Packaging, and Potency of AAV Vectors
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
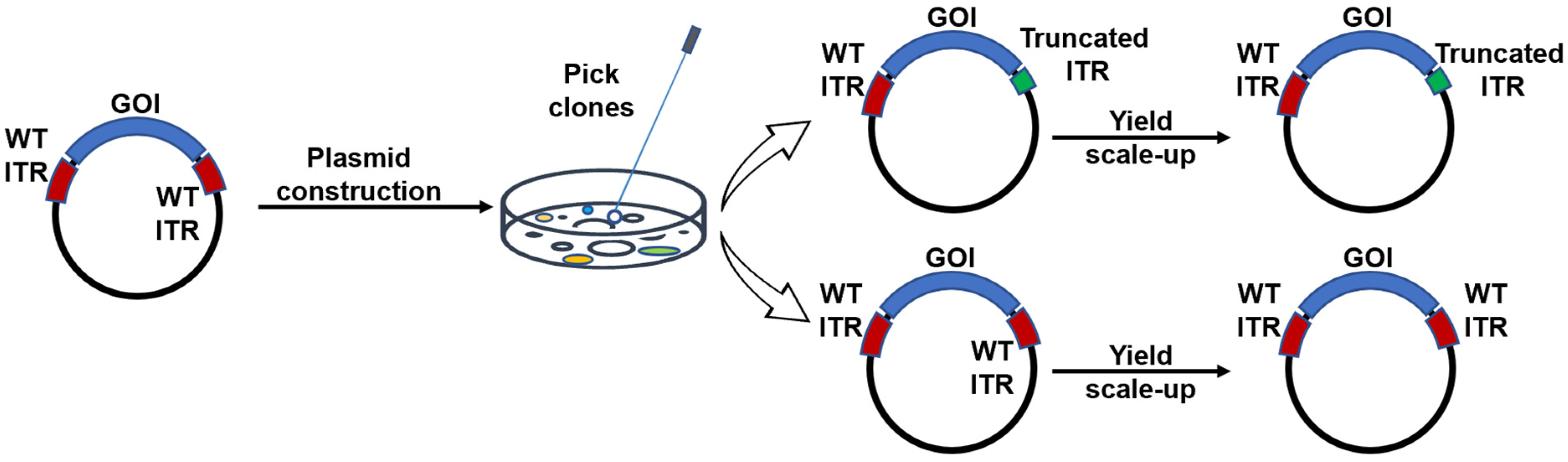
2.1. Plasmid Design and Construction
2.2. AAV Production
2.3. AAV Titration by Droplet Digital PCR (ddPCR)
2.4. AAV Purification by Solid-Phase Extraction (SPE)
2.5. Empty/Full Capsid Measurement by High-Performance Liquid Chromatography (HPLC)
2.6. AAV DNA Checking by Nanopore Sequencing
2.7. In Vitro Potency
2.8. AAV Genome Replication for Dual Deletion Design with ddPCR
3. Results
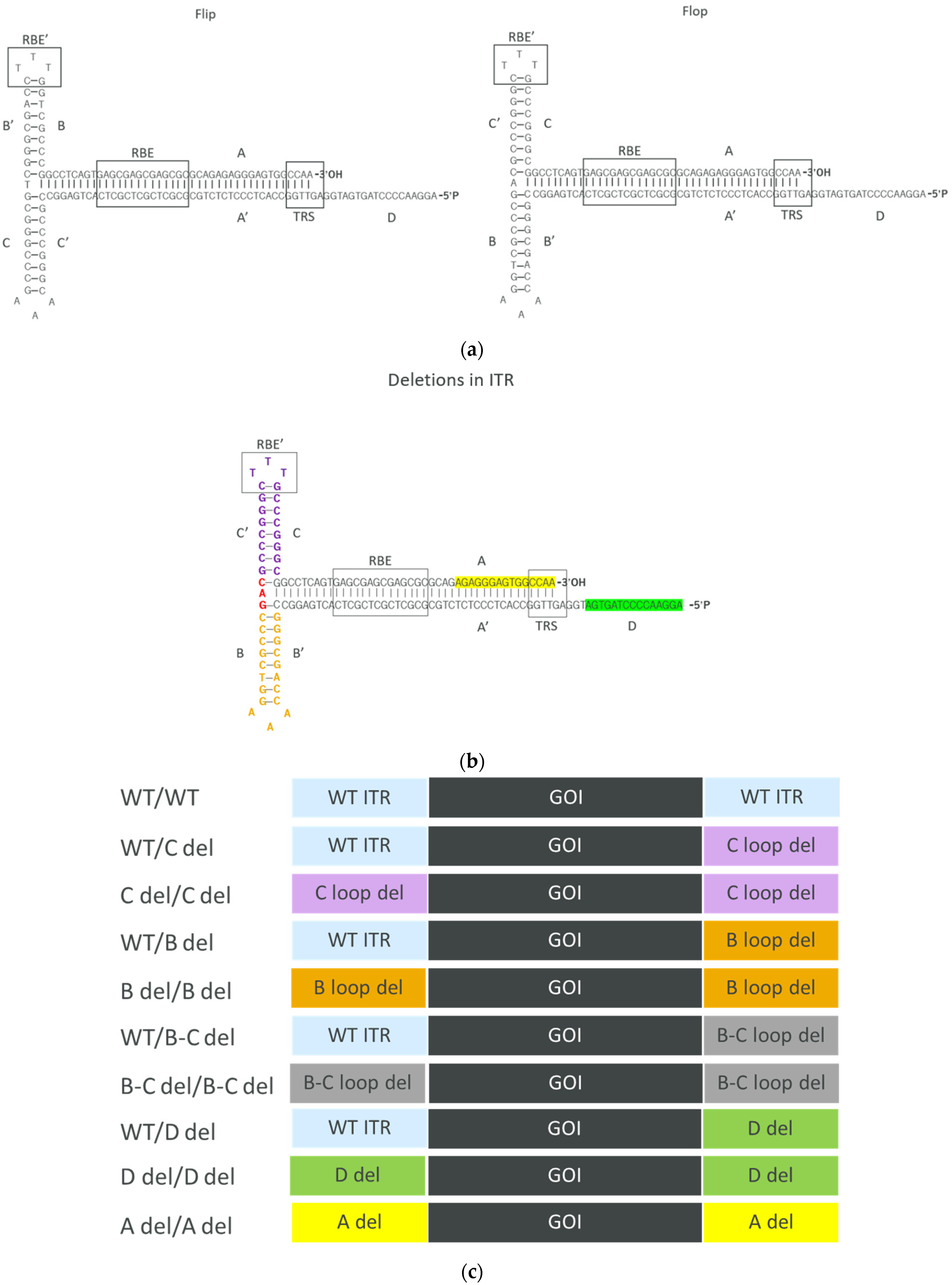
3.1. Design of ITRs with Different Deletions
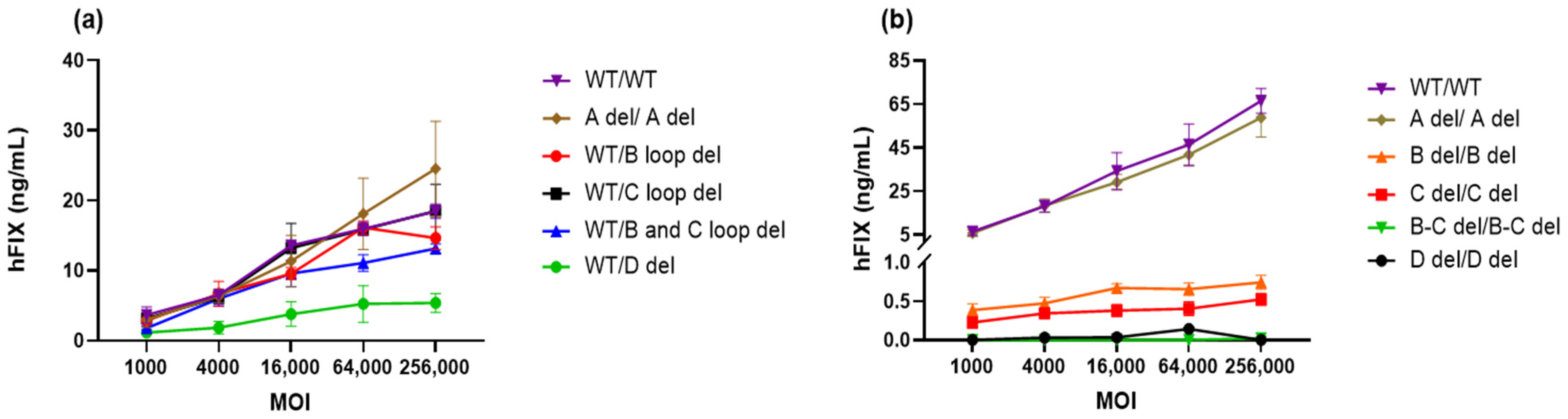
3.2. Productivity of Vectors with ITRs Possessing Different Deletions
3.3. Reduced Packaging Efficiency Leads to Diminished Yield
3.4. Diminished Replication Is a Critical Factor Contributing to the Low Yield in Vectors with Deletions in Both ITRs
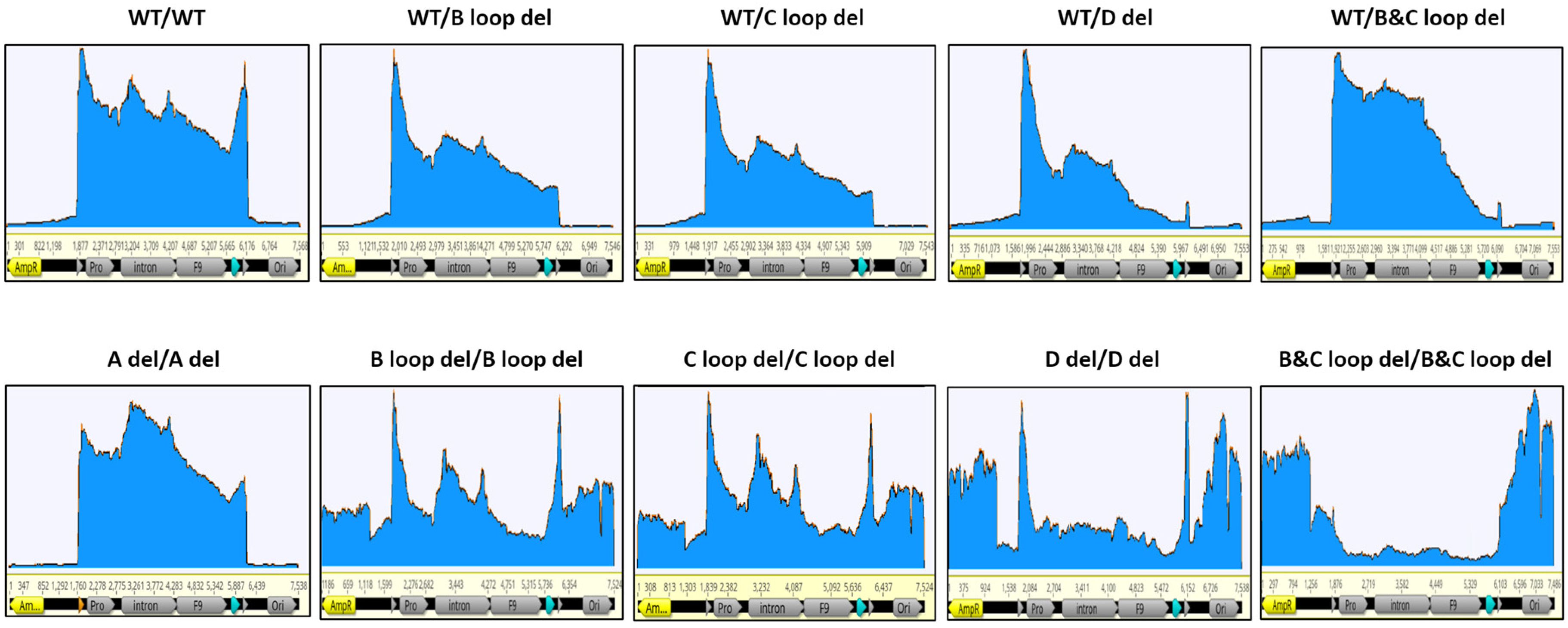
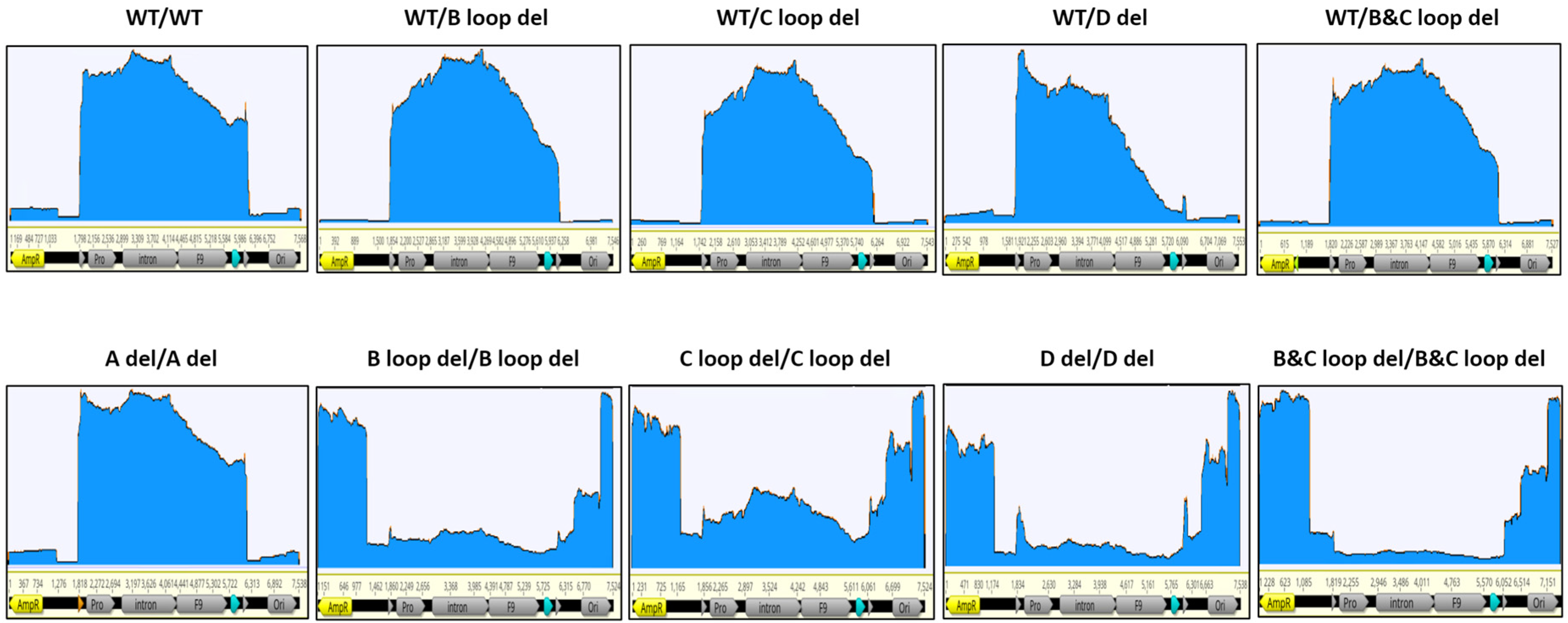
3.5. Assessment of the Packaged DNA Integrity Using Nanopore Sequencing
3.6. Assessment for the Biological Potency of the Vectors with Modified ITRs In Vitro
4. Discussion
4.1. ITR Function in rAAV and Influence of Potential Truncations
4.2. NGS for Checking Plasmid and Vector Homogeneity
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef]
- Flotte, T.R.; Berns, K.I. Adeno-associated viral vectors for gene therapy. Lab. Tech. Biochem. Mol. Biol. 2005, 31, 255–272. [Google Scholar] [CrossRef]
- Young, S.M.; Samulski, R.J. Adeno-associated virus (AAV) site-specific recombination does not require a Rep-dependent origin of replication within the AAV terminal repeat. Proc. Natl. Acad. Sci. USA 2001, 98, 13525–13530. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, M.D.; Linden, R.M. Adeno-Associated Virus Biology. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 807. [Google Scholar] [CrossRef]
- Wilmott, P.; Lisowski, L.; Alexander, I.E.; Logan, G.J. A User’s Guide to the Inverted Terminal Repeats of Adeno-Associated Virus. Hum. Gene Ther. Methods 2019, 30, 206–213. [Google Scholar] [CrossRef]
- Savy, A.; Dickx, Y.; Nauwynck, L.; Bonnin, D.; Merten, O.W.; Galibert, L. Impact of Inverted Terminal Repeat Integrity on rAAV8 Production Using the Baculovirus/Sf9 Cells System. Hum. Gene Ther. Methods 2017, 28, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Shitik, E.M.; Shalik, I.K.; Yudkin, D.V. AAV-based vector improvements unrelated to capsid protein modification. Front. Med. 2023, 10, 1106085. [Google Scholar] [CrossRef] [PubMed]
- Feiner, R.C.; Teschner, J.; Teschner, K.E.; Radukic, M.T.; Baumann, T.; Hagen, S.; Hannappel, Y.; Biere, N.; Anselmetti, D.; Arndt, K.M.; et al. rAAV Engineering for Capsid-Protein Enzyme Insertions and Mosaicism Reveals Resilience to Mutational, Structural and Thermal Perturbations. Int. J. Mol. Sci. 2019, 20, 5702. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Tian, W.; Liu, C.; Lian, Z.; Dong, X.; Wu, X. Deletion of the B-B’ and C-C’ regions of inverted terminal repeats reduces rAAV productivity but increases transgene expression. Sci. Rep. 2017, 7, 5432. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Qing, K.; Ponnazhagan, S.; Srivastava, A. Adeno-associated virus type 2 DNA replication in vivo: Mutation analyses of the D sequence in viral inverted terminal repeats. J. Virol. 1997, 71, 3077–3082. [Google Scholar] [CrossRef]
- Ryan, J.H.; Zolotukhin, S.; Muzyczka, N. Sequence requirements for binding of Rep68 to the adeno-associated virus terminal repeats. J. Virol. 1996, 70, 1542–1553. [Google Scholar] [CrossRef]
- Snyder, R.O.; Im, D.S.; Ni, T.; Xiao, X.; Samulski, R.J.; Muzyczka, N. Features of the adeno-associated virus origin involved in substrate recognition by the viral Rep protein. J. Virol. 1993, 67, 6096–6104. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Yue, Y.; Boftsi, M.; Wasala, L.P.; Tran, N.T.; Zhang, K.; Pintel, D.J.; Tai, P.W. Rational engineering of a functional CpG-free ITR for AAV gene therapy. Gene Ther. 2022, 29, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zak, R.; Zhang, Y.; Engelhardt, J.F. Inverted Terminal Repeat Sequences Are Important for Intermolecular Recombination and Circularization of Adeno-Associated Virus Genomes. J. Virol. 2005, 79, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Yin, Z.; Li, J.; Zhang, D.; Aslanidi, G.; Srivastava, A. Strategies to generate high-titer, high-potency recombinant AAV3 serotype vectors. Mol. Ther. Methods Clin. Dev. 2016, 3, 29. [Google Scholar] [CrossRef]
- Tran, N.T.; Lecomte, E.; Saleun, S.; Namkung, S.; Robin, C.; Weber, K. Human and Insect Cell-Produced Recombinant Adeno-Associated Viruses Show Differences in Genome Heterogeneity. Hum. Gene Ther. 2022, 33, 371–388. [Google Scholar] [CrossRef]
- Mroske, C.; Rivera, H.; Ul-Hasan, T.; Chatterjee, S.; Wong, K.K., Jr. Naturally Occurring Non-Functional Mutations of the AAV ITR. Mol. Ther. 2011, 19 (Suppl. 1), S51. [Google Scholar] [CrossRef]
- Samulski, R.J.; Srivastava, A.; Berns, K.I.; Muzyczka, N. Rescue of adeno-associated virus from recombinant plasmids: Gene correction within the terminal repeats of AAV. Cell 1983, 33, 135–143. [Google Scholar] [CrossRef]
- Samulski, R.J.; Berns, K.I.; Tan, M.; Muzyczka, N. Cloning of adeno-associated virus into pBR322: Rescue of intact virus from the recombinant plasmid in human cells. Proc. Natl. Acad. Sci. USA 1982, 79, 2077–2081. [Google Scholar] [CrossRef]
- Grimm, D.; Lee, J.S.; Wang, L.; Desai, T.; Akache, B.; Storm, T.A.; Kay, M.A. In Vitro and In Vivo Gene Therapy Vector Evolution via Multispecies Interbreeding and Retargeting of Adeno-Associated Viruses. J. Virol. 2008, 82, 5887–5911. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Lusby, E.W.; Berns, K.I. Nucleotide sequence and organization of the adeno-associated virus 2 genome. J. Virol. 1983, 45, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Linden, R.M.; Ward, P.; Giraud, C.; Winocour, E.; Berns, K.I. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA 1996, 93, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, J.; Xiao, X. Absence of T-Shaped Structure and Deletions of B and C Hairpins Have Minimal Effects on Essential Functions of AAV Inverted Terminal Repeats. Mol. Ther. 2004, 9 (Suppl. 1), S6. [Google Scholar] [CrossRef]
- Earley, L.F.; Conatser, L.M.; Lue, V.M.; Dobbins, A.L.; Li, C.; Hirsch, M.L.; Samulski, R.J. Adeno-Associated Virus Serotype-Specific Inverted Terminal Repeat Sequence Role in Vector Transgene Expression. Hum. Gene Ther. 2020, 31, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.F. Product-related impurities in clinical-grade recombinant AAV vectors: Characterization and risk assessment. Biomedicines 2014, 2, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Chadeuf, G.; Ciron, C.; Moullier, P.; Salvetti, A. Evidence for encapsidation of prokaryotic sequences during recombinant adeno-associated virus production and their in vivo persistence after vector delivery. Mol. Ther. 2005, 12, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Schnödt, M.; Schmeer, M.; Kracher, B.; Krüsemann, C.; Espinosa, L.E.; Grünert, A. DNA Minicircle Technology Improves Purity of Adeno-associated Viral Vector Preparations. Mol. Ther. Nucleic Acids 2016, 5, e355. [Google Scholar] [CrossRef]
- Lecomte, E.; Tournaire, B.; Cogné, B.; Dupont, J.B.; Lindenbaum, P.; Martin-Fontaine, M.; Broucque, F.; Robin, C.; Hebben, M.; Merten, O.-M.; et al. Advanced characterization of DNA molecules in rAAV vector preparations by single-stranded virus next-generation sequencing. Mol. Ther. Nucleic Acids 2015, 4, e260. [Google Scholar] [CrossRef]
- Guerin, K.; Rego, M.; Bourges, D.; Ersing, I.; Haery, L.; Harten DeMaio, K.; Sanders, E.; Tasissa, M.; Kostman, M.; Tillgren, M.; et al. A Novel Next-Generation Sequencing and Analysis Platform to Assess the Identity of Recombinant Adeno-Associated Viral Preparations from Viral DNA Extracts. Hum. Gene Ther. 2020, 31, 664–678. [Google Scholar] [CrossRef]
- Namkung, S.; Tran, N.T.; Manokaran, S.; He, R.; Su, Q.; Xie, J.; Gao, G.; Tai, P.W.L. Direct ITR-to-ITR Nanopore Sequencing of AAV Vector Genomes. Hum. Gene Ther. 2022, 33, 1187–1196. [Google Scholar] [CrossRef]
- Tai, P.W.L.; Xie, J.; Fong, K.; Seetin, M.; Heiner, C.; Su, Q.; Weiand, M.; Wilmot, D.; Zapp, M.L.; Gao, G. Adeno-associated Virus Genome Population Sequencing Achieves Full Vector Genome Resolution and Reveals Human-Vector Chimeras. Mol. Ther. Methods Clin. Dev. 2018, 9, 130–141. [Google Scholar] [CrossRef]
- Radukic, M.T.; Brandt, D.; Haak, M.; Muller, K.M.; Kalinowski, J. Nanopore sequencing of native adeno-Associated virus (AAV) single-stranded DNA using a transposase-based rapid protocol. NAR Genom. Bioinform. 2020, 2, lqaa074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chrzanowski, M.; Frabutt, D.A.; Lam, A.K.; Mulcrone, P.L.; Li, L.; Konkle, B.A.; Miao, C.H.; Xiao, W.H. Cryptic resolution sites in the vector plasmid lead to the heterogeneities in the rAAV vectors. J. Med. Virol. 2023, 95, e28433. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Hu, S.; Lee, W.; Walsh, N.; Iozza, K.; Huang, N.; Preston, G.; Drouin, L.M.; Jia, N.; Deng, J.; et al. A Comprehensive Study of the Effects by Sequence Truncation within Inverted Terminal Repeats (ITRs) on the Productivity, Genome Packaging, and Potency of AAV Vectors. Microorganisms 2024, 12, 310. https://doi.org/10.3390/microorganisms12020310
Chen Y, Hu S, Lee W, Walsh N, Iozza K, Huang N, Preston G, Drouin LM, Jia N, Deng J, et al. A Comprehensive Study of the Effects by Sequence Truncation within Inverted Terminal Repeats (ITRs) on the Productivity, Genome Packaging, and Potency of AAV Vectors. Microorganisms. 2024; 12(2):310. https://doi.org/10.3390/microorganisms12020310
Chicago/Turabian StyleChen, Yinxing, Shiliang Hu, William Lee, Noel Walsh, Kayla Iozza, Neil Huang, Gregory Preston, Lauren M. Drouin, Nannan Jia, June Deng, and et al. 2024. "A Comprehensive Study of the Effects by Sequence Truncation within Inverted Terminal Repeats (ITRs) on the Productivity, Genome Packaging, and Potency of AAV Vectors" Microorganisms 12, no. 2: 310. https://doi.org/10.3390/microorganisms12020310
APA StyleChen, Y., Hu, S., Lee, W., Walsh, N., Iozza, K., Huang, N., Preston, G., Drouin, L. M., Jia, N., Deng, J., Hebben, M., & Liao, J. (2024). A Comprehensive Study of the Effects by Sequence Truncation within Inverted Terminal Repeats (ITRs) on the Productivity, Genome Packaging, and Potency of AAV Vectors. Microorganisms, 12(2), 310. https://doi.org/10.3390/microorganisms12020310