
Soil Microbiome in Conditions of Oil Pollution of Subarctic Ecosystems

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sample Collection
2.3. Soil Chemical Analyses
2.4. Soil Metabarcoding Analysis
2.5. Statistical Analyses
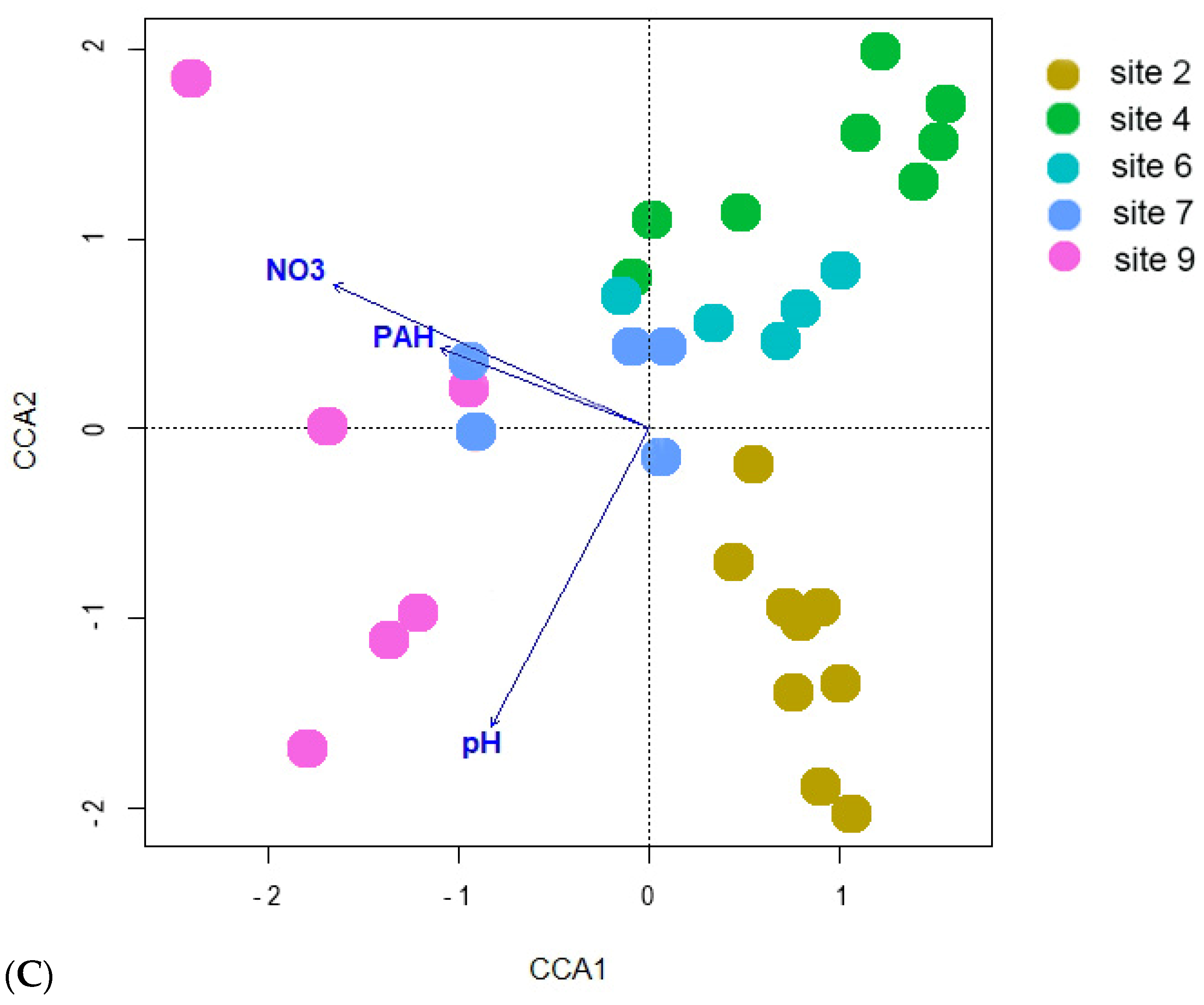
3. Results
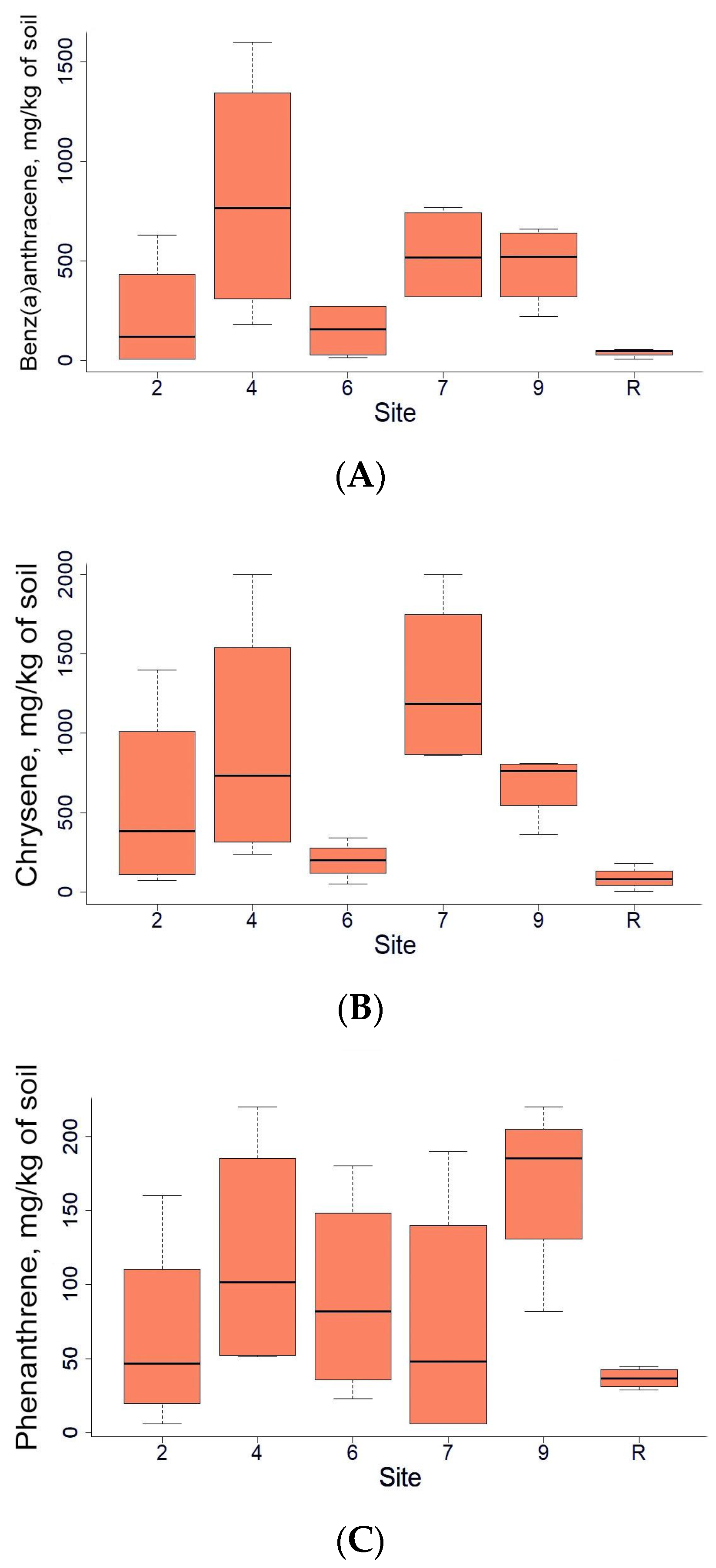
3.1. Chemical Properties of the Soil
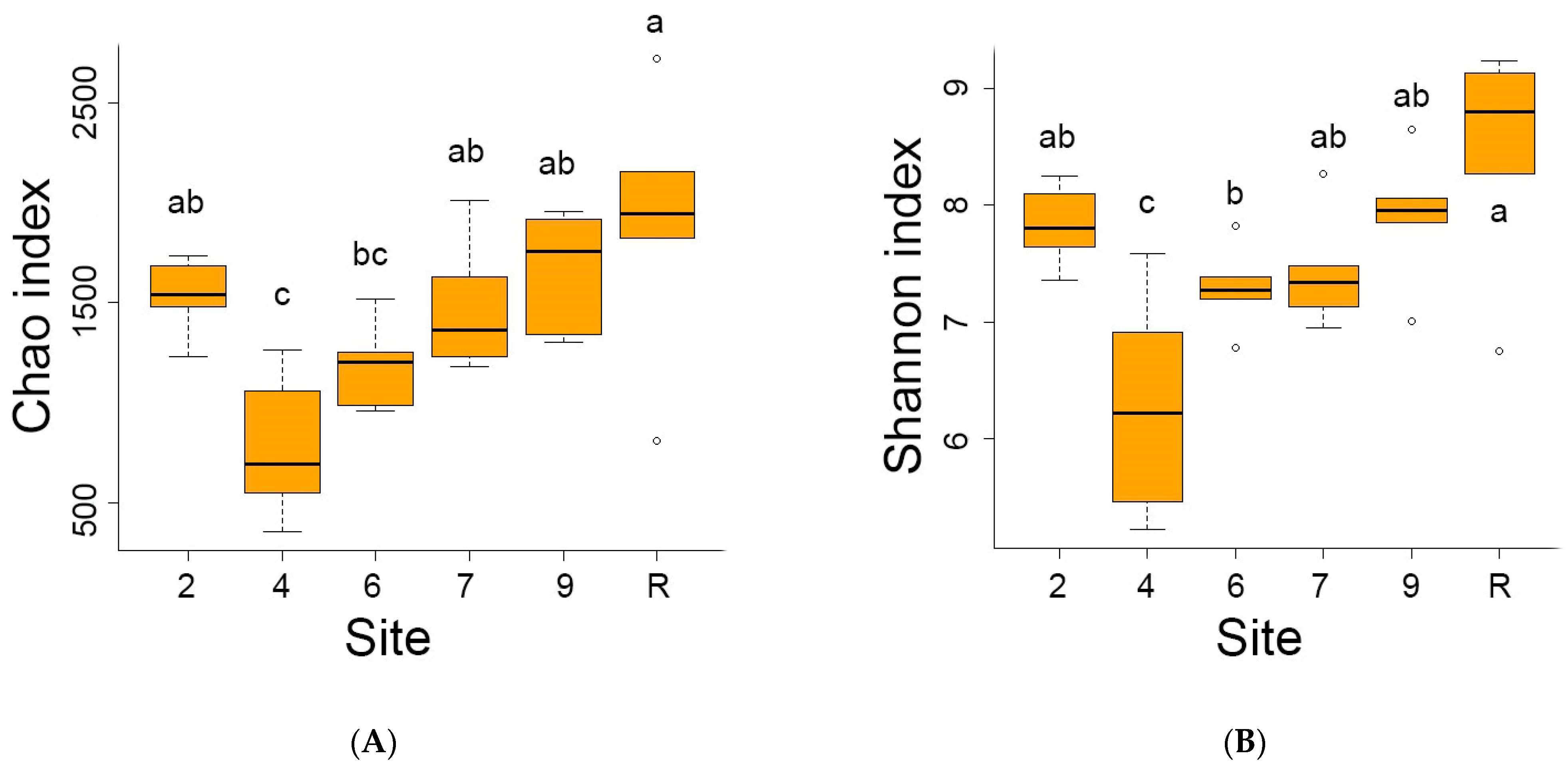
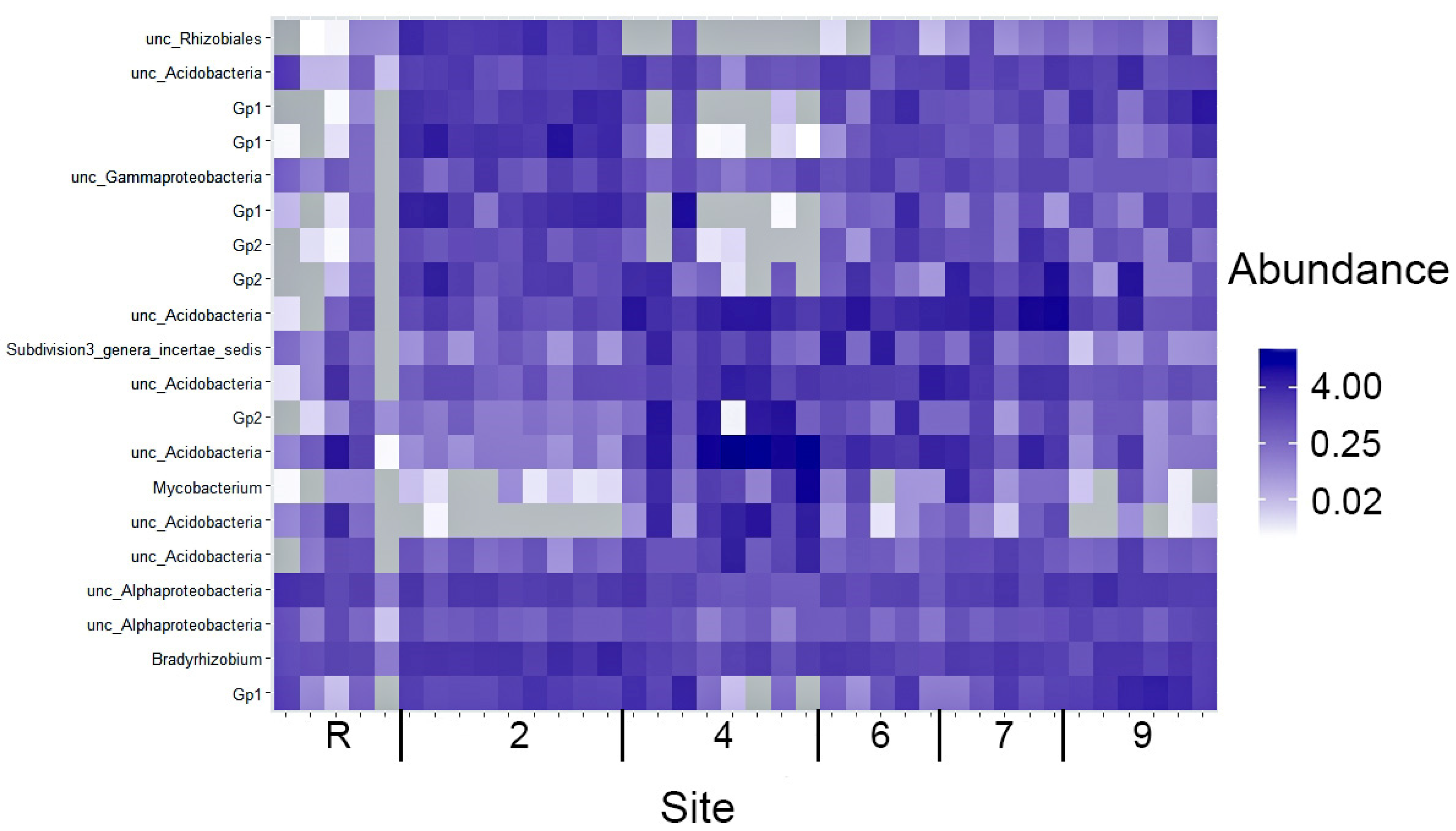
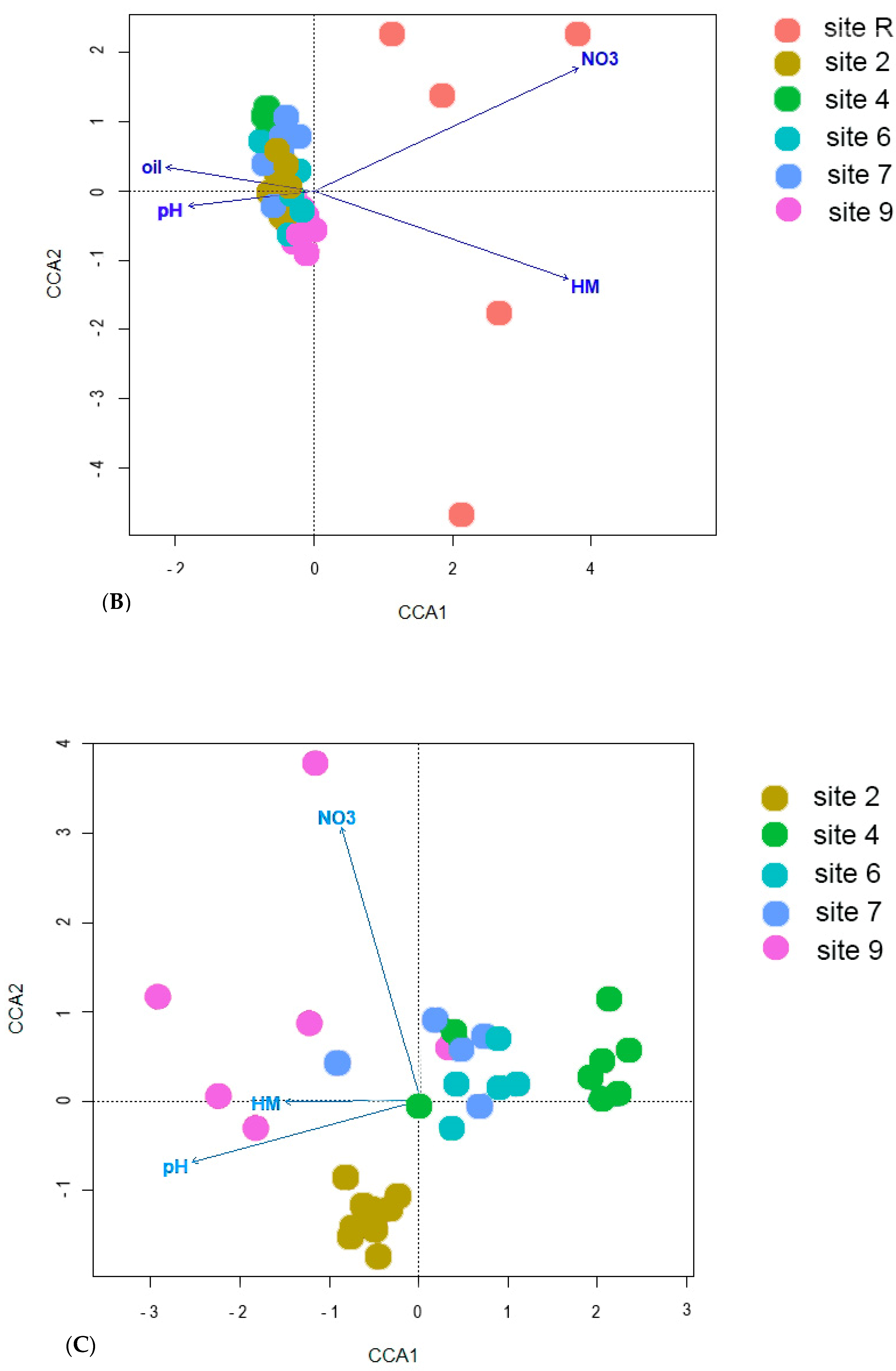
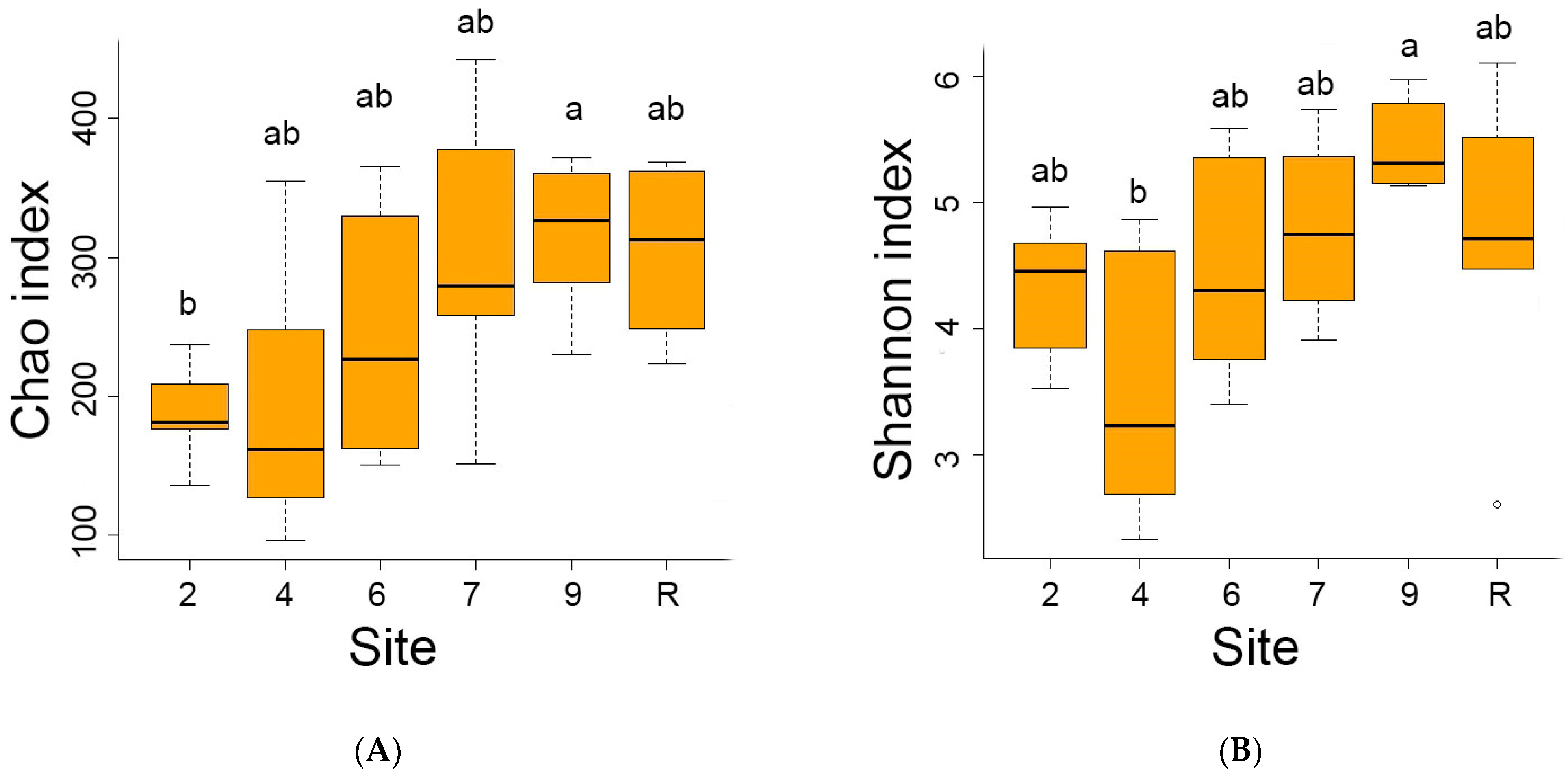
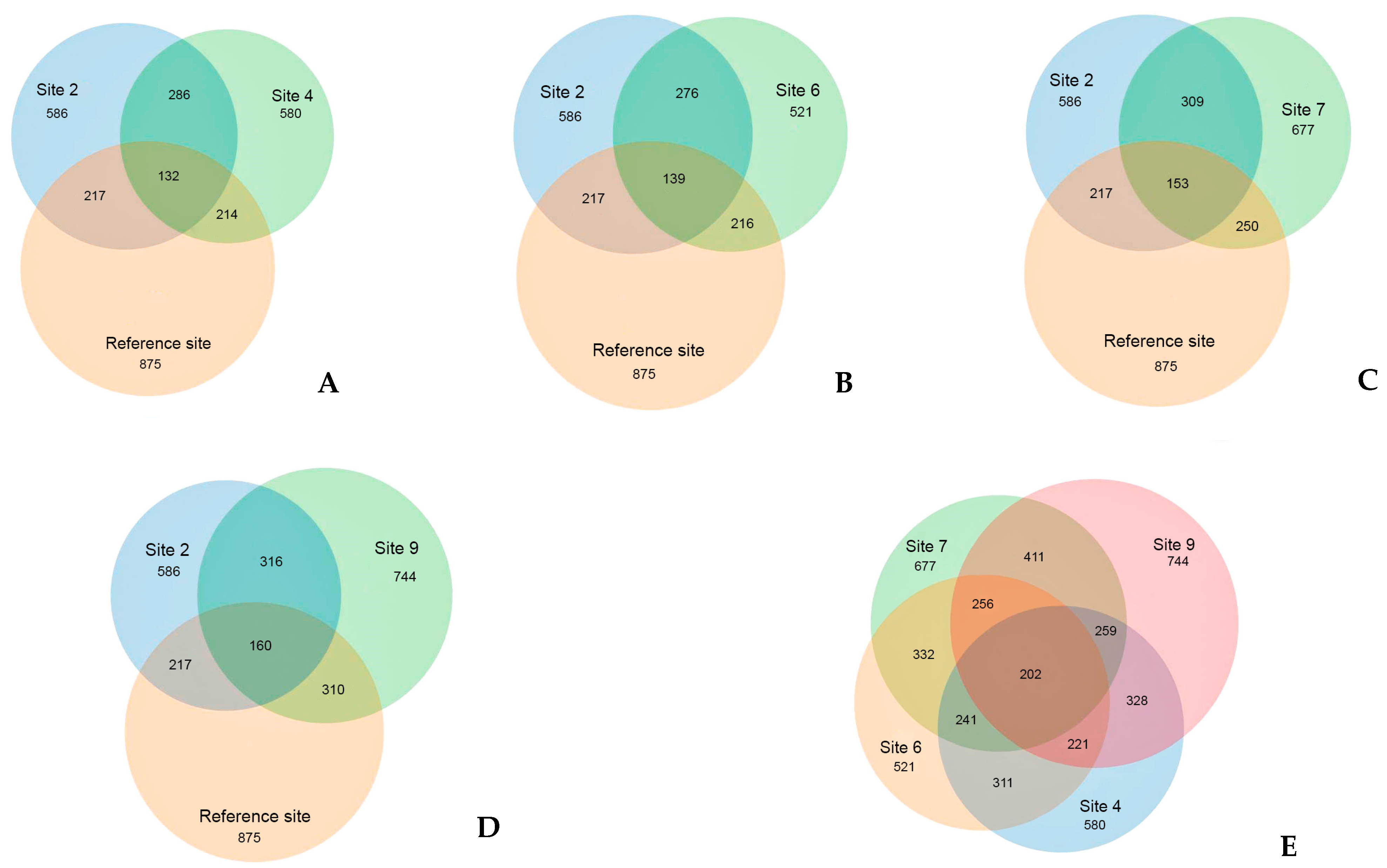
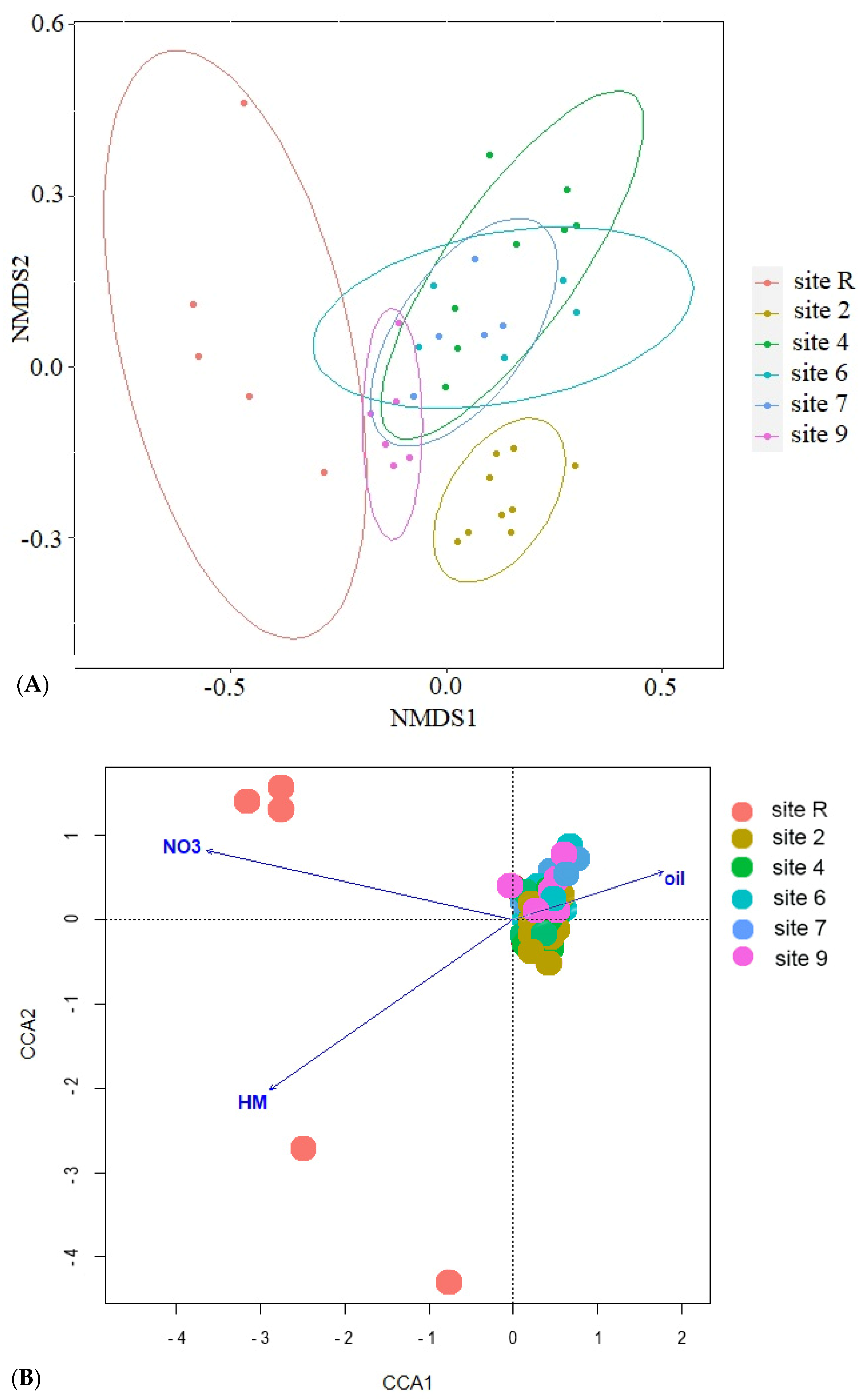
3.2. Bacteria Diversity
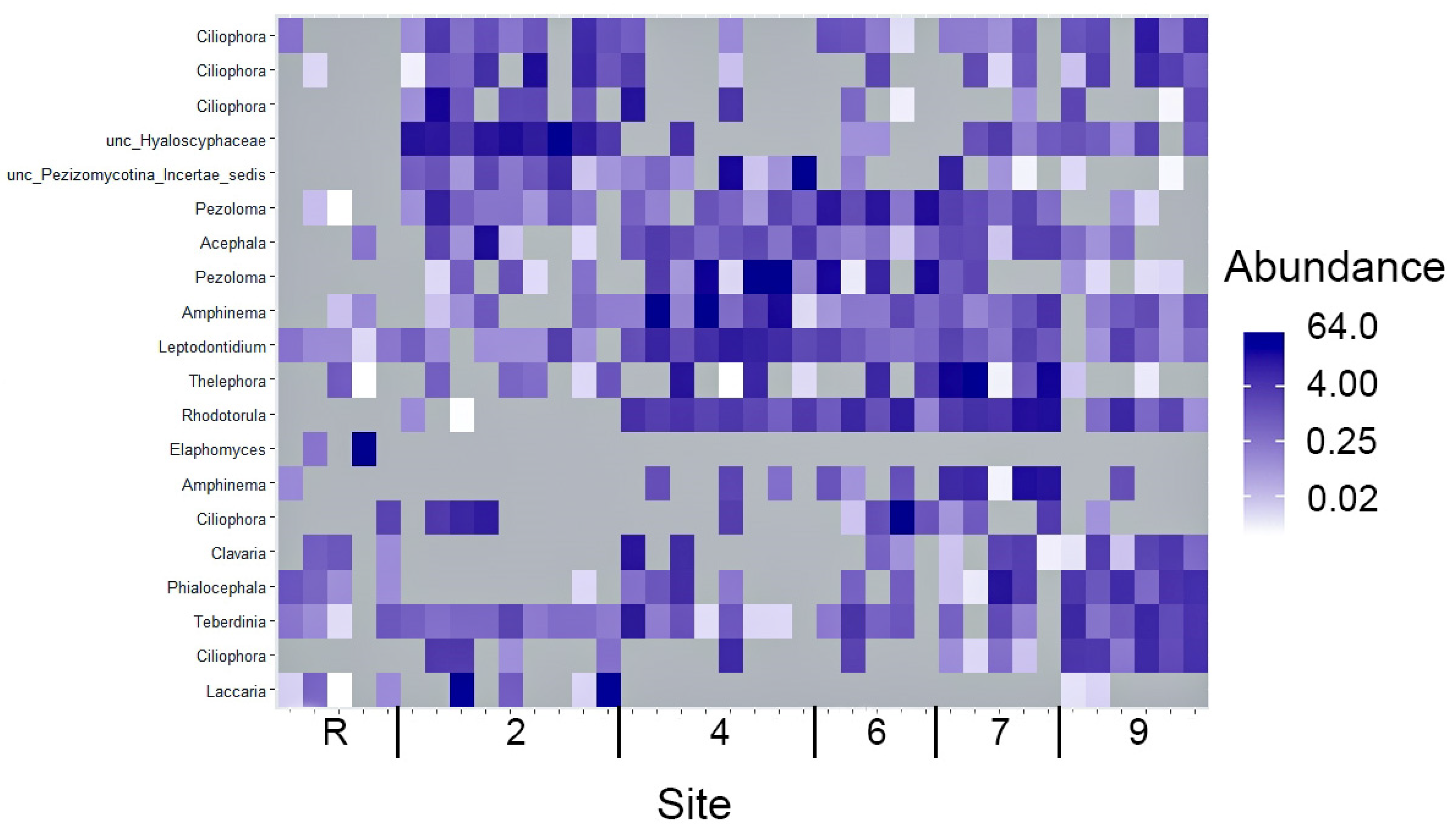
3.3. Fungi Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.R.S.; Al-Baldawi, I.A.; Almansoory, A.F.; Purwanti, I.F.; Al-Sbani, N.H.; Sharuddin, S.S.N. Plant-Assisted Remediation of Hydrocarbons in Water and Soil: Application, Mechanisms, Challenges and Opportunities. Chemosphere 2020, 247, 125932. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Gnansounou, E.; Pandey, A. Comprehensive Review on Toxicity of Persistent Organic Pollutants from Petroleum Refinery Waste and Their Degradation by Microorganisms. Chemosphere 2017, 188, 280–291. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, J.J.; Maranho, L.T. Rhizospheric Microorganisms as a Solution for the Recovery of Soils Contaminated by Petroleum: A Review. J. Environ. Manag. 2018, 210, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Aislabie, J.; Saul, D.J.; Foght, J.M. Bioremediation of Hydrocarbon-Contaminated Polar Soils. Extremophiles 2006, 10, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Sinkko, H.; Penttinen, P.; Lindström, K. Characterization of Successional Changes in Bacterial Community Composition during Bioremediation of Used Motor Oil-Contaminated Soil in a Boreal Climate. Sci. Total Environ. 2016, 542, 817–825. [Google Scholar] [CrossRef]
- Davoodi, S.M.; Miri, S.; Taheran, M.; Brar, S.K.; Galvez-Cloutier, R.; Martel, R. Bioremediation of Unconventional Oil Contaminated Ecosystems under Natural and Assisted Conditions: A Review. Environ. Sci. Technol. 2020, 54, 2054–2067. [Google Scholar] [CrossRef]
- Shahi, A.; Ince, B.; Aydin, S.; Ince, O. Assessment of the Horizontal Transfer of Functional Genes as a Suitable Approach for Evaluation of the Bioremediation Potential of Petroleum-Contaminated Sites: A Mini-Review. Appl. Microbiol. Biotechnol. 2017, 101, 4341–4348. [Google Scholar] [CrossRef]
- Maganov, R.U.; Markarova, M.Y.; Muliak, V.V.; Zagvozdkin, V.K.; Zaikin, I.A. Nature Conservation Measures on the Refinery Companies. Prt 1: Rehabilitation of Petroleum Polluted Soils in Usinsk District, Komi Republic; Publishing house of the Komi Scientific Center of the Ural Branch of the Russian Academy of Sciences: Syktyvkar, Russia, 2006. [Google Scholar]
- Melekhina, E.N.; Markarova, M.Y.; Shchemelinina, T.N.; Anchugova, E.M.; Kanev, V.A. Secondary Successions of Biota in Oil-Polluted Peat Soil upon Different Biological Remediation Methods. Eurasian Soil Sci. 2015, 48, 643–653. [Google Scholar] [CrossRef]
- Wang, S.; Wang, D.; Yu, Z.; Dong, X.; Liu, S.; Cui, H.; Sun, B. Advances in Research on Petroleum Biodegradability in Soil. Environ. Sci. Process. Impacts 2021, 23, 9–27. [Google Scholar] [CrossRef]
- Soleimani, M.; Afyuni, M.; Hajabbasi, M.A.; Nourbakhsh, F.; Sabzalian, M.R.; Christensen, J.H. Phytoremediation of an Aged Petroleum Contaminated Soil Using Endophyte Infected and Non-Infected Grasses. Chemosphere 2010, 81, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Echartea, E.; Strejcek, M.; Mukherjee, S.; Uhlik, O.; Yrjälä, K. Bacterial Succession in Oil-Contaminated Soil under Phytoremediation with Poplars. Chemosphere 2020, 243, 125242. [Google Scholar] [CrossRef] [PubMed]
- Shahi, A.; Aydin, S.; Ince, B.; Ince, O. Evaluation of Microbial Population and Functional Genes during the Bioremediation of Petroleum-Contaminated Soil as an Effective Monitoring Approach. Ecotoxicol. Environ. Saf. 2016, 125, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Margesin, R.; Schinner, F. Efficiency of Indigenous and Inoculated Cold-Adapted Soil Microorganisms for Biodegradation of Diesel Oil in Alpine Soils. Appl. Environ. Microbiol. 1997, 63, 2660–2664. [Google Scholar] [CrossRef] [PubMed]
- Melekhina, E.N. Recovery Successions of Soil Microfauna in Oil-Polluted Ecosystems of the European Subarctic. Biol. Bull. 2020, 47, 97–105. [Google Scholar] [CrossRef]
- Melekhina, E.N.; Belykh, E.S.; Markarova, M.Y.; Taskaeva, A.A.; Rasova, E.E.; Baturina, O.A.; Kabilov, M.R.; Velegzhaninov, I.O. Soil Microbiota and Microarthropod Communities in Oil Contaminated Sites in the European Subarctic. Sci. Rep. 2021, 11, 19620. [Google Scholar] [CrossRef]
- Aksenov, A.S.; Shirokova, L.S.; Kisil, O.Y.; Kolesova, S.N.; Lim, A.G.; Kuzmina, D.; Pouillé, S.; Alexis, M.A.; Castrec-Rouelle, M.; Loiko, S.V.; et al. Bacterial Number and Genetic Diversity in a Permafrost Peatland (Western Siberia): Testing a Link with Organic Matter Quality and Elementary Composition of a Peat Soil Profile. Diversity 2021, 13, 328. [Google Scholar] [CrossRef]
- Bonomo, M.G.; Calabrone, L.; Scrano, L.; Bufo, S.A.; Di Tomaso, K.; Buongarzone, E.; Salzano, G. Metagenomic Monitoring of Soil Bacterial Community after the Construction of a Crude Oil Flowline. Environ. Monit. Assess. 2022, 194, 48. [Google Scholar] [CrossRef]
- Taskaev, A.I. (Ed.) Atlas of the Komi Republic on Climate and Hydrology; Drofa, DiK: Moskva, Russia, 1997; 116p. (In Russian) [Google Scholar]
- Dobrovol’skii, G.V.; Taskaev, A.I.; Zaboeva, I.V. (Eds.) Atlas of Soils of the Republic of Komi; Komi Republican Printing House: Syktyvkar, Russia, 2010. [Google Scholar]
- Novakovskiy, A.B.; Kanev, V.A.; Markarova, M.Y. Long-Term Dynamics of Plant Communities after Biological Remediation of Oil-Contaminated Soils in Far North. Sci. Rep. 2021, 11, 4888. [Google Scholar] [CrossRef]
- Markarova, M.Y. Experience of using the biological product “Universal” for the reclamation of oil-contaminated lands. Vestn. Inst. Biol. Komi Sci. Cent. Ural. Branch Russ. Acad. Sci. 2004, 10, 21–23. [Google Scholar]
- Murygina, V.P.; Voishvillo, N.E.; Kalyuzhny, S.V. Biological Product “Roder” for Cleaning Soils, Fresh and Mineralized Waters from Oil and Oil Products. Patent RU 2174496, 10 October 2001. [Google Scholar]
- GOST 26423-85 2011; Soils. Methods for Determination of Specific Electric Conductivity, pH and Solid Residue of Water Extract. Interstate Standards: Moscow, Russia, 1986.
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An Improved Dual-Indexing Approach for Multiplexed 16S rRNA Gene Sequencing on the Illumina MiSeq Platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Naumova, N.B.; Savenkov, O.A.; Alikina, T.Y.; Kabilov, M.R. Rhizosphere Bacteriobiome of the Husk Tomato Grown in the Open Field in West Siberia. Agriculture (Pol’nohospodárstvo) 2019, 65, 147–154. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UNOISE2: Improved Error-Correction for Illumina 16S and ITS Amplicon Sequencing. Bioinformatics 2016. [Google Scholar] [CrossRef]
- Edgar, R.C. SINTAX: A Simple Non-Bayesian Taxonomy Classifier for 16S and ITS Sequences. Bioinformatics 2016. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE USEARCH/UTAX release for eukaryotes. Version 04.02.2020. UNITE Community. 2020. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R Package for Rarefaction and Extrapolation of Species Diversity (H Ill Numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. Community Ecology Package. Vegan Package 2013, 10, 719. [Google Scholar]
- About the Procedure of Assessment of the Degree of Loss from Contamination of Soil with Chemical Compounds; A letter of Ministry of protection of environment and nature resources of Russian Federation from 27.12.1993 year. №04-25-61-5678; Ministry of Environmental Protection and Natural Resources of the Russian Federation: Moscow, Russia, 1993.
- Beznosikov, V.A.; Lodygin, E.D. Ecological-Geochemical Assessment of Hydrocarbons in Soils of Northeastern European Russia. Eurasian Soil Sci. 2010, 43, 550–555. [Google Scholar] [CrossRef]
- Patel, A.B.; Shaikh, S.; Jain, K.R.; Desai, C.; Madamwar, D. Polycyclic Aromatic Hydrocarbons: Sources, Toxicity, and Remediation Approaches. Front. Microbiol. 2020, 11, 562813. [Google Scholar] [CrossRef]
- Patowary, K.; Patowary, R.; Kalita, M.C.; Deka, S. Development of an Efficient Bacterial Consortium for the Potential Remediation of Hydrocarbons from Contaminated Sites. Front. Microbiol. 2016, 7, 1092. [Google Scholar] [CrossRef] [PubMed]
- Patowary, R.; Devi, A.; Mukherjee, A.K. Advanced Bioremediation by an Amalgamation of Nanotechnology and Modern Artificial Intelligence for Efficient Restoration of Crude Petroleum Oil-Contaminated Sites: A Prospective Study. Environ. Sci. Pollut. Res. 2023, 30, 74459–74484. [Google Scholar] [CrossRef] [PubMed]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of Bacterial and Fungal Communities to High Petroleum Pollution in Different Soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Brassington, K.J.; Prpich, G.; Paton, G.I.; Semple, K.T.; Pollard, S.J.T.; Coulon, F. Insights into the Biodegradation of Weathered Hydrocarbons in Contaminated Soils by Bioaugmentation and Nutrient Stimulation. Chemosphere 2016, 161, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Murygina, V.P.; Markarova, M.Y.; Kalyuzhnyi, S.V. Application of Biopreparation “Rhoder” for Remediation of Oil Polluted Polar Marshy Wetlands in Komi Republic. Environ. Int. 2005, 31, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, D.W.; Kwon, S.L.; Heo, Y.M.; Jang, S.; Kwon, B.-O.; Khim, J.S.; Kim, G.-H.; Kim, J.-J. Importance of Functional Diversity in Assessing the Recovery of the Microbial Community after the Hebei Spirit Oil Spill in Korea. Environ. Int. 2019, 128, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Al-Otibi, F.; Al-Zahrani, R.M.; Marraiki, N. The Crude Oil Biodegradation Activity of Candida Strains Isolated from Oil-Reservoirs Soils in Saudi Arabia. Sci. Rep. 2022, 12, 10708. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-L.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y.; Sekiguchi, Y. Syntrophorhabdus aromaticivorans Gen. Nov., Sp. Nov., the First Cultured Anaerobe Capable of Degrading Phenol to Acetate in Obligate Syntrophic Associations with a Hydrogenotrophic Methanogen. Appl. Environ. Microbiol. 2008, 74, 2051–2058. [Google Scholar] [CrossRef]
- Liesack, W.; Bak, F.; Kreft, J.-U.; Stackebrandt, E. Holophaga foetida Gen. Nov., Sp. Nov., a New, Homoacetogenic Bacterium Degrading Methoxylated Aromatic Compounds. Arch. Microbiol. 1994, 162, 85–90. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil Bacterial Community Composition Altered by Increased Nutrient Availability in Arctic Tundra Soils. Front. Microbiol. 2014, 5, 516. [Google Scholar] [CrossRef]
- Peng, M.; Zi, X.; Wang, Q. Bacterial Community Diversity of Oil-Contaminated Soils Assessed by High Throughput Sequencing of 16S rRNA Genes. Int. J. Environ. Res. Public Health 2015, 12, 12002–12015. [Google Scholar] [CrossRef] [PubMed]
- Makhalanyane, T.P.; Van Goethem, M.W.; Cowan, D.A. Microbial Diversity and Functional Capacity in Polar Soils. Curr. Opin. Biotechnol. 2016, 38, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Hu, X.; Li, X.; Zhang, Y.; Jiang, L.; Li, J.; Guan, Z.; Cai, Y.; Liao, X. The Rhizospheric Microbial Community Structure and Diversity of Deciduous and Evergreen Forests in Taihu Lake Area, China. PLoS ONE 2017, 12, e0174411. [Google Scholar] [CrossRef] [PubMed]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef]
- Weiß, M.; Waller, F.; Zuccaro, A.; Selosse, M. Sebacinales—One Thousand and One Interactions with Land Plants. New Phytol. 2016, 211, 20–40. [Google Scholar] [CrossRef]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 2015, 86, 158–164. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Bio-Recultivation Method | Design | Fertilizers | Seeded Plants |
|---|---|---|---|---|
| Site 2 | Control. Only mechanical oil removal. No biopreparations | - | No fertilizing | No seed plants |
| Site 4 | Biopreparation “Universal” 1 | yeasts Rhodotorula glutinis and bacteria Rhodococcus egvi, Rhodococcus erythropolis, Pseudomonas fuorescens | Mineral fertilizers 3 | Phleum pratense, Agrostis gigantea, Avena sativa |
| Site 6 | Biopreparation “Universal” 1, lignin sorbents, BAG | yeasts Rhodotorula glutinis and bacteria Rhodococcus egvi, Rhodococcus erythropolis, Pseudomonas fuorescens | Compost and mineral fertilizers | Deschampsia cespitosa |
| Site 7 | Phytoremediation (without biopreparation) | - | Mineral fertilizers, dolomitic meal | Avena sativa, Phleum pratense |
| Site 9 | Biopreparation “Roder” 2 | bacteria Rhodococcus ruber and Rhodococcus erythropolis | Mineral fertilizers and lime | Phalaris arundinacea, Phleum pratense, Avena sativa |
| Site R | A reference site. Not subjected to oil contamination and other anthropogenic influences | - | - | - |
| Site | 2 | 4 | 6 | 7 | 9 | R 1 |
|---|---|---|---|---|---|---|
| pH | 5.1 ± 0.1 * | 4.7 ± 0.2 | 4.7 ± 0.2 | 4.9 ± 0.1 | 5.2 ± 0.3 * | 4.8 ± 0.3 |
| N (NH4) | 60.2 ± 18.5 * | 82.9 ± 19.5 * | 125.3 ± 12.9 * | 71.1 ± 9.2 * | 80.4 ± 22.3 * | 808.6 ± 62.6 |
| N (NO3) | 8.1 ± 1.7 * | 9.6 ± 1.0 * | 14.0 ± 3.2 * | 19.8 ± 4.4 * | 20.4 ± 17.8 * | 145.3 ± 103.4 |
| Cu | 8.0 ± 6.0 | 4.4 ± 1.6 | 4.6 ± 0.2 | 5.2 ± 0.8 | 5.8 ± 1.1 | 6.5 ± 1.8 |
| Pb | 3.7 ± 0.5 * | 2.7 ± 0.3 * | 4.2 ± 1.5 * | 3.8 ± 1.2 * | 4.1 ± 1.7 * | 11.4 ± 2.0 |
| Cd | 0.3 ± 0.03 | 0.3 ± 0.08 | 0.3 ± 0.02 | 0.2 ± 0.05 * | 0.4 ± 0.06 | 0.4 ± 0.09 |
| Zn | 7.5 ± 1.8 * | 6.4 ± 2.9 * | 5.0 ± 1.3 * | 16.6 ± 4.3 | 10.6 ± 3.3 * | 23.8 ± 6.8 |
| TPH × 103/Decrease 2 | 62.4 ± 29.2 */3.3 | 71.2 ± 93.8 */4.1 | 9.8 ± 3.1 */33.0 | 72.3 ± 28.6 */3.1 | 42.3 ± 46.4 */7.2 | 0.9 ± 0.7/3.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melekhina, E.N.; Belykh, E.S.; Kanev, V.A.; Taskaeva, A.A.; Tarabukin, D.V.; Zinovyeva, A.N.; Velegzhaninov, I.O.; Rasova, E.E.; Baturina, O.A.; Kabilov, M.R.; et al. Soil Microbiome in Conditions of Oil Pollution of Subarctic Ecosystems. Microorganisms 2024, 12, 80. https://doi.org/10.3390/microorganisms12010080
Melekhina EN, Belykh ES, Kanev VA, Taskaeva AA, Tarabukin DV, Zinovyeva AN, Velegzhaninov IO, Rasova EE, Baturina OA, Kabilov MR, et al. Soil Microbiome in Conditions of Oil Pollution of Subarctic Ecosystems. Microorganisms. 2024; 12(1):80. https://doi.org/10.3390/microorganisms12010080
Chicago/Turabian StyleMelekhina, Elena N., Elena S. Belykh, Vladimir A. Kanev, Anastasia A. Taskaeva, Dmitry V. Tarabukin, Aurika N. Zinovyeva, Ilya O. Velegzhaninov, Elena E. Rasova, Olga A. Baturina, Marsel R. Kabilov, and et al. 2024. "Soil Microbiome in Conditions of Oil Pollution of Subarctic Ecosystems" Microorganisms 12, no. 1: 80. https://doi.org/10.3390/microorganisms12010080
APA StyleMelekhina, E. N., Belykh, E. S., Kanev, V. A., Taskaeva, A. A., Tarabukin, D. V., Zinovyeva, A. N., Velegzhaninov, I. O., Rasova, E. E., Baturina, O. A., Kabilov, M. R., & Markarova, M. Y. (2024). Soil Microbiome in Conditions of Oil Pollution of Subarctic Ecosystems. Microorganisms, 12(1), 80. https://doi.org/10.3390/microorganisms12010080