

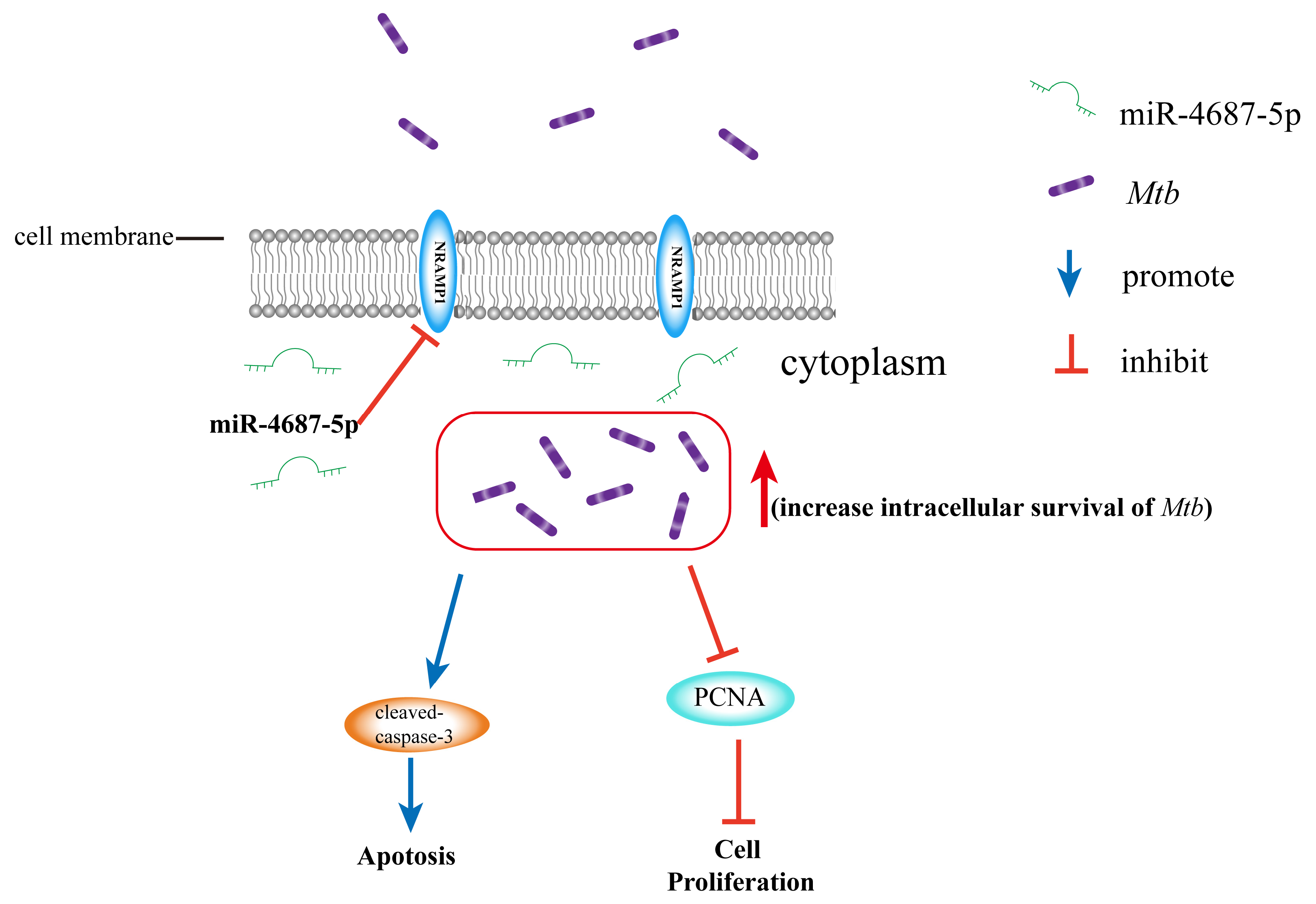
miR-4687-5p Affects Intracellular Survival of Mycobacterium tuberculosis through Its Regulation of NRAMP1 Expression in A549 Cells
 , and
, and 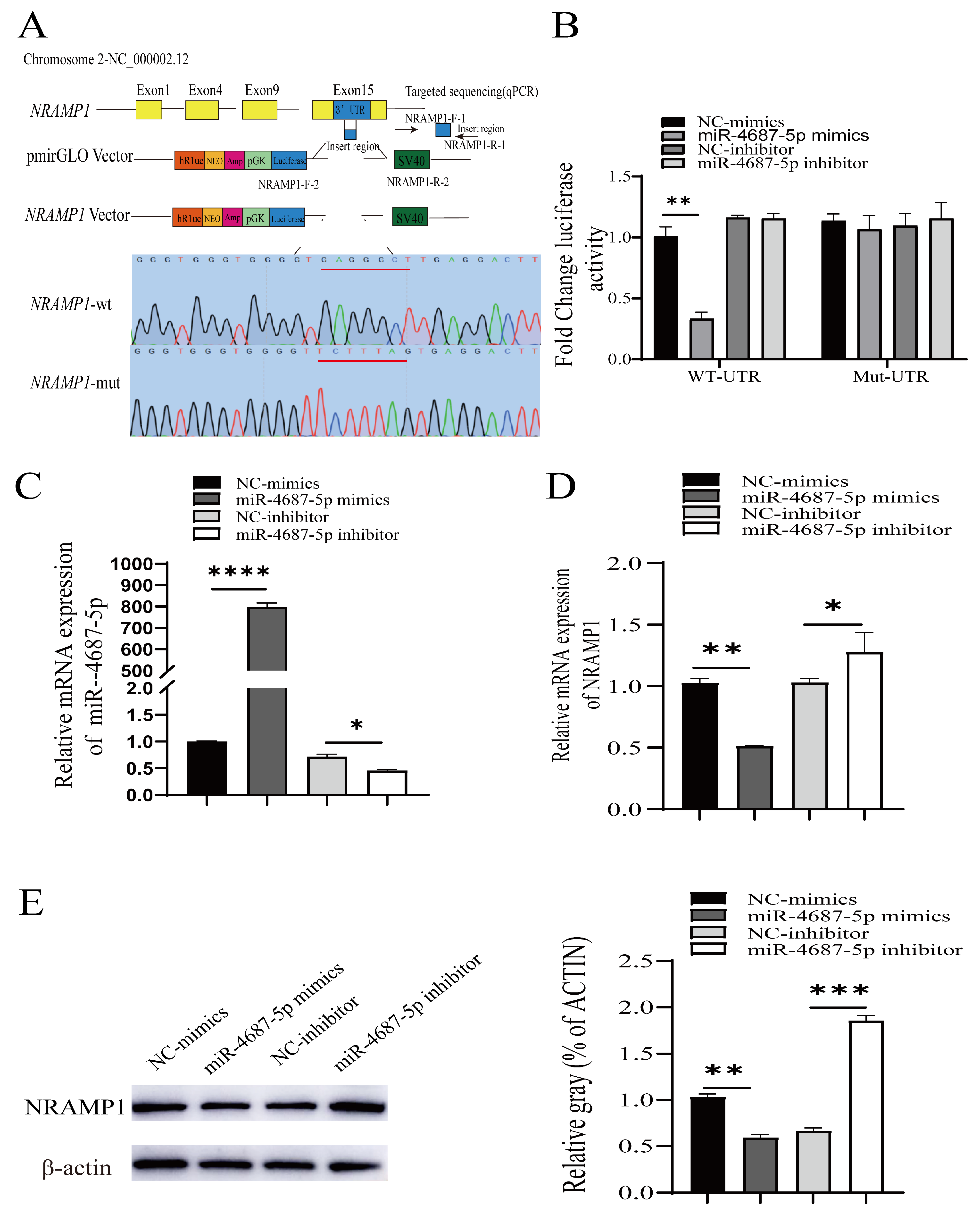{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell and Bacteria Culture
2.2. Cell Transfection
2.3. Luciferase Reporter Assay
2.4. Cell Proliferation Assay
2.5. Cell Apoptosis Test
2.6. Mtb Infection Assay and CFU
2.7. Western Blotting
2.8. qRT-PCR
2.9. Immunofluorescence
2.10. Statistical Analysis
3. Results
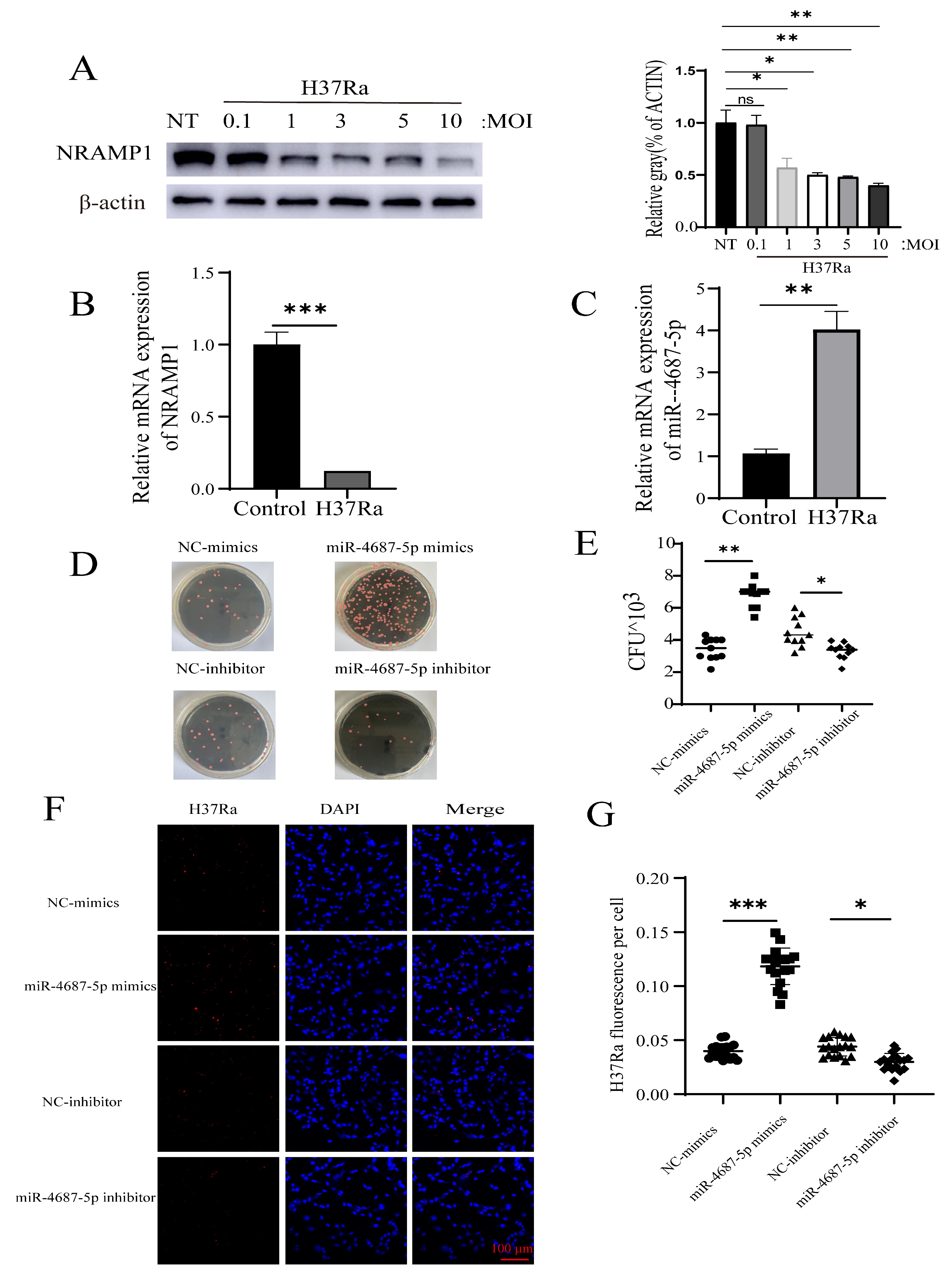
3.1. Identification of NRAMP1 as a Direct Target of miR-4687-5p
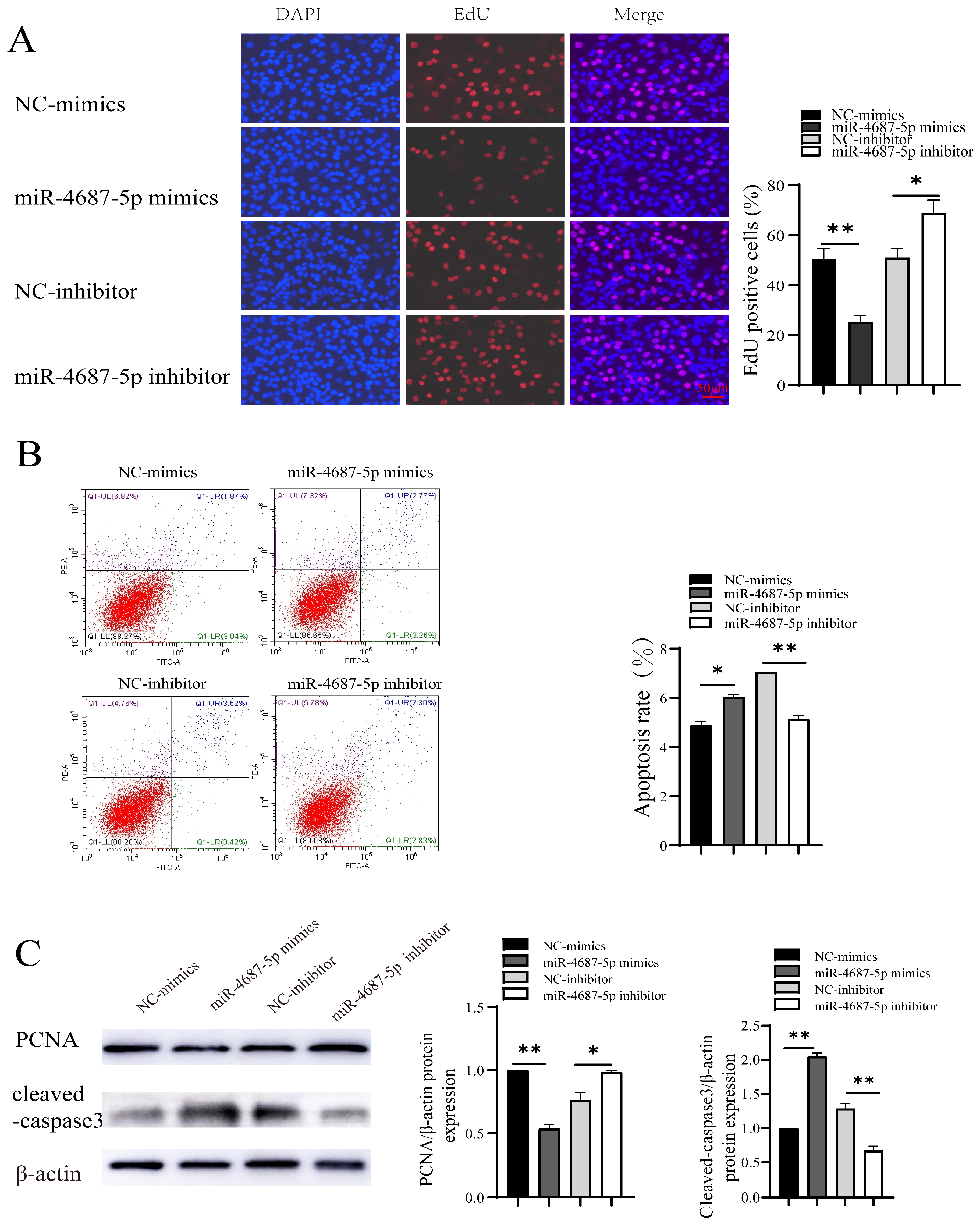
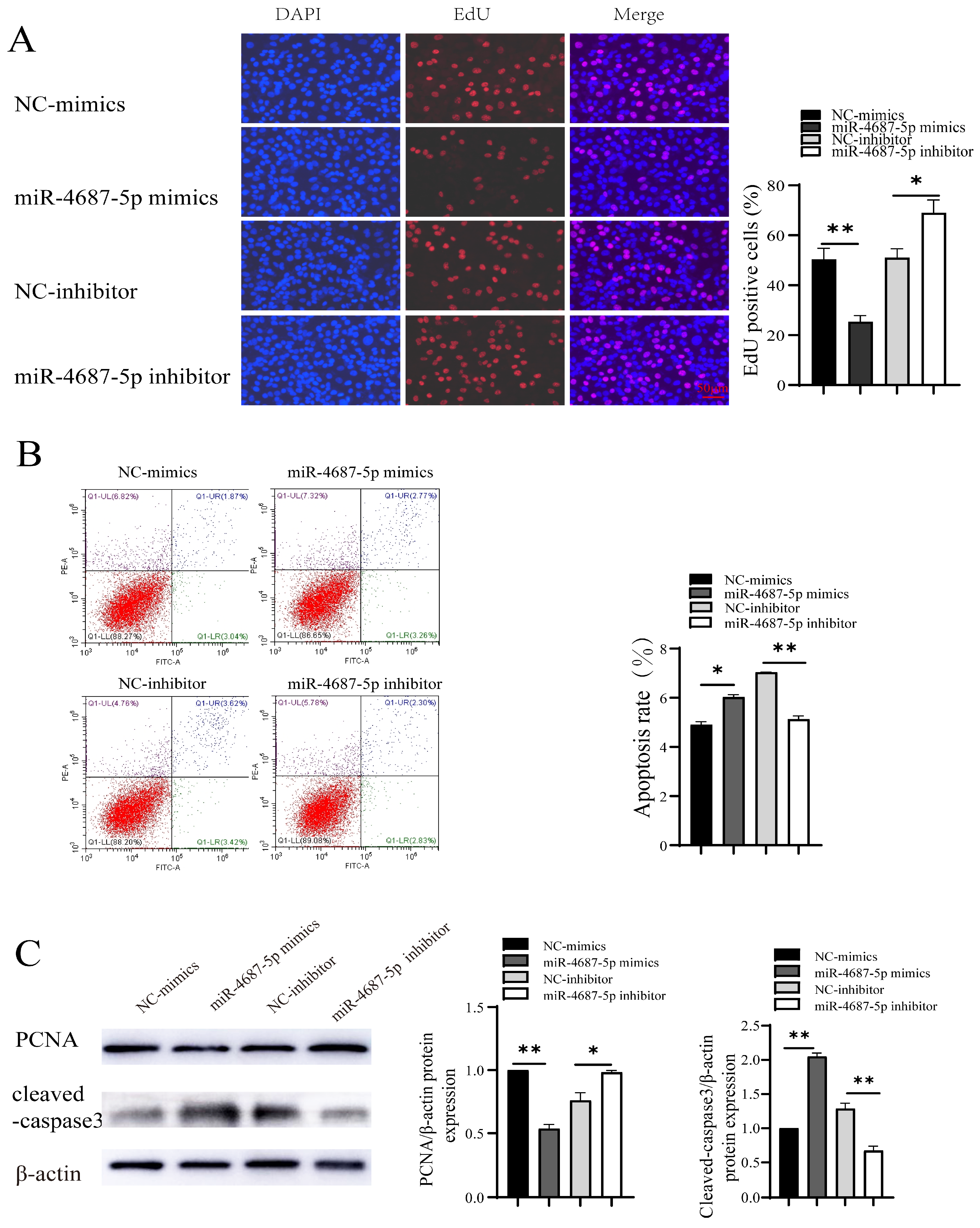
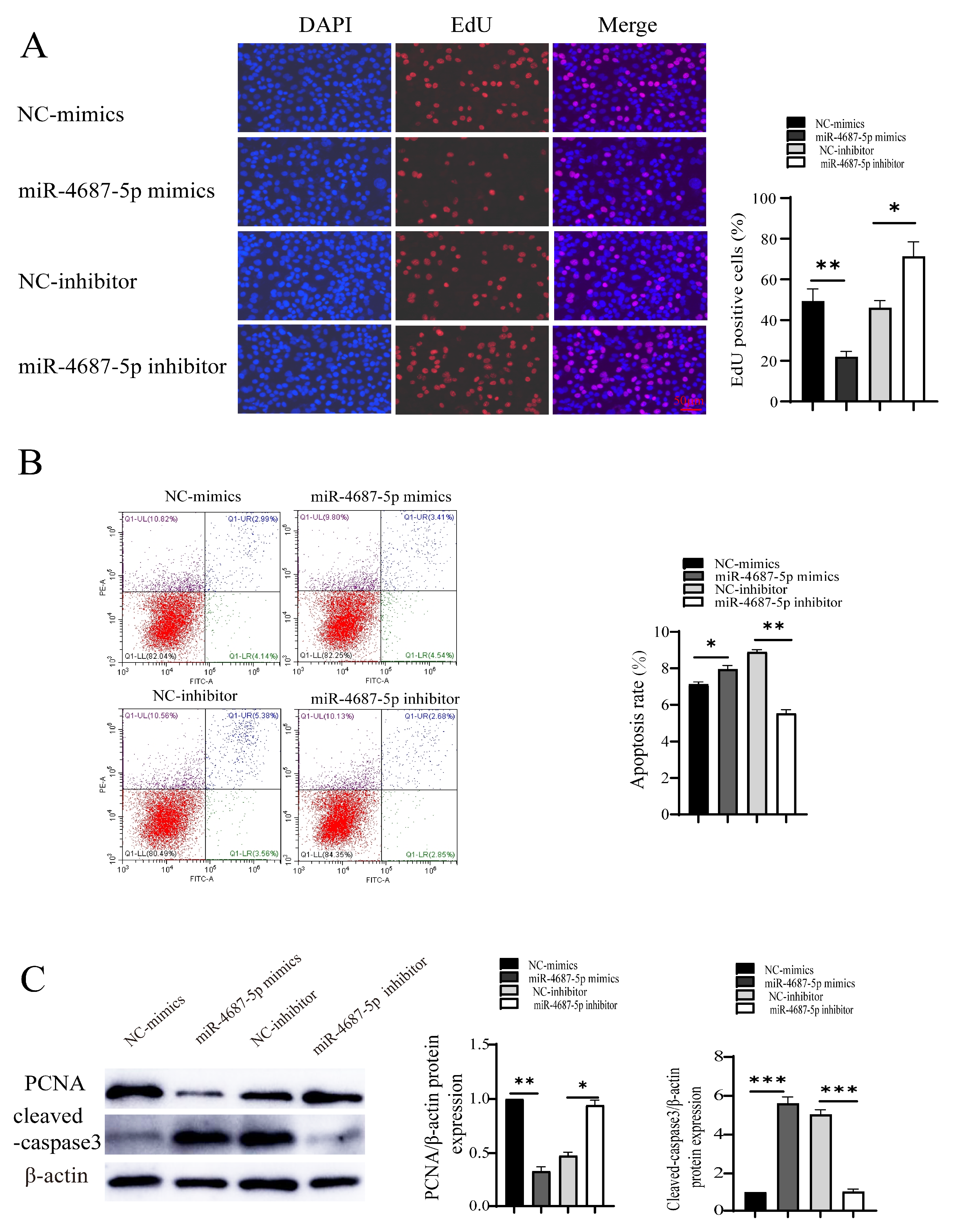
3.2. The Role of miR-4687-5p/NRAMP1 on Cell Proliferation and Apoptosis
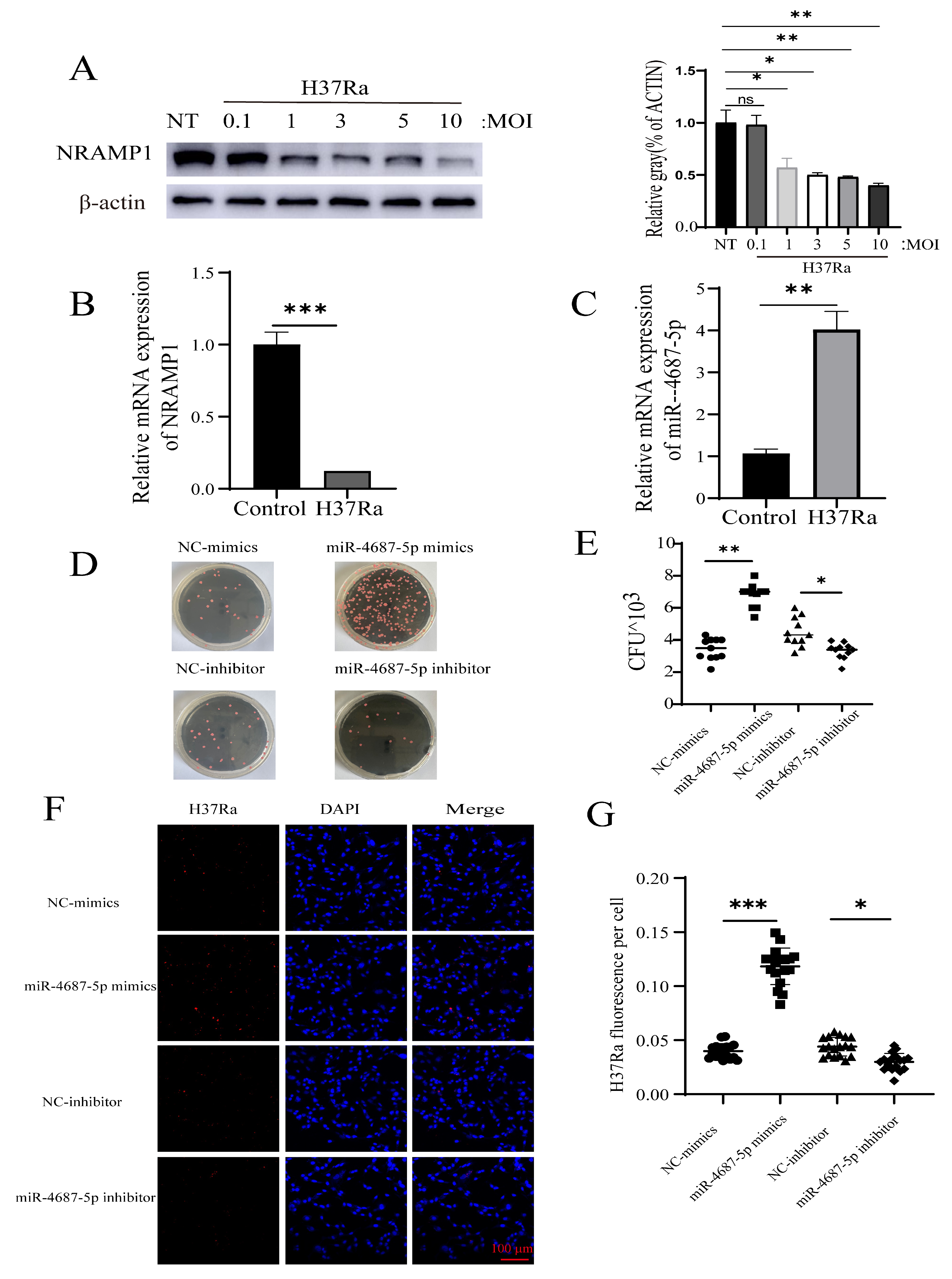
3.3. miR-4687-5p Affects Mtb Susceptibility via Targeting NRAMP1
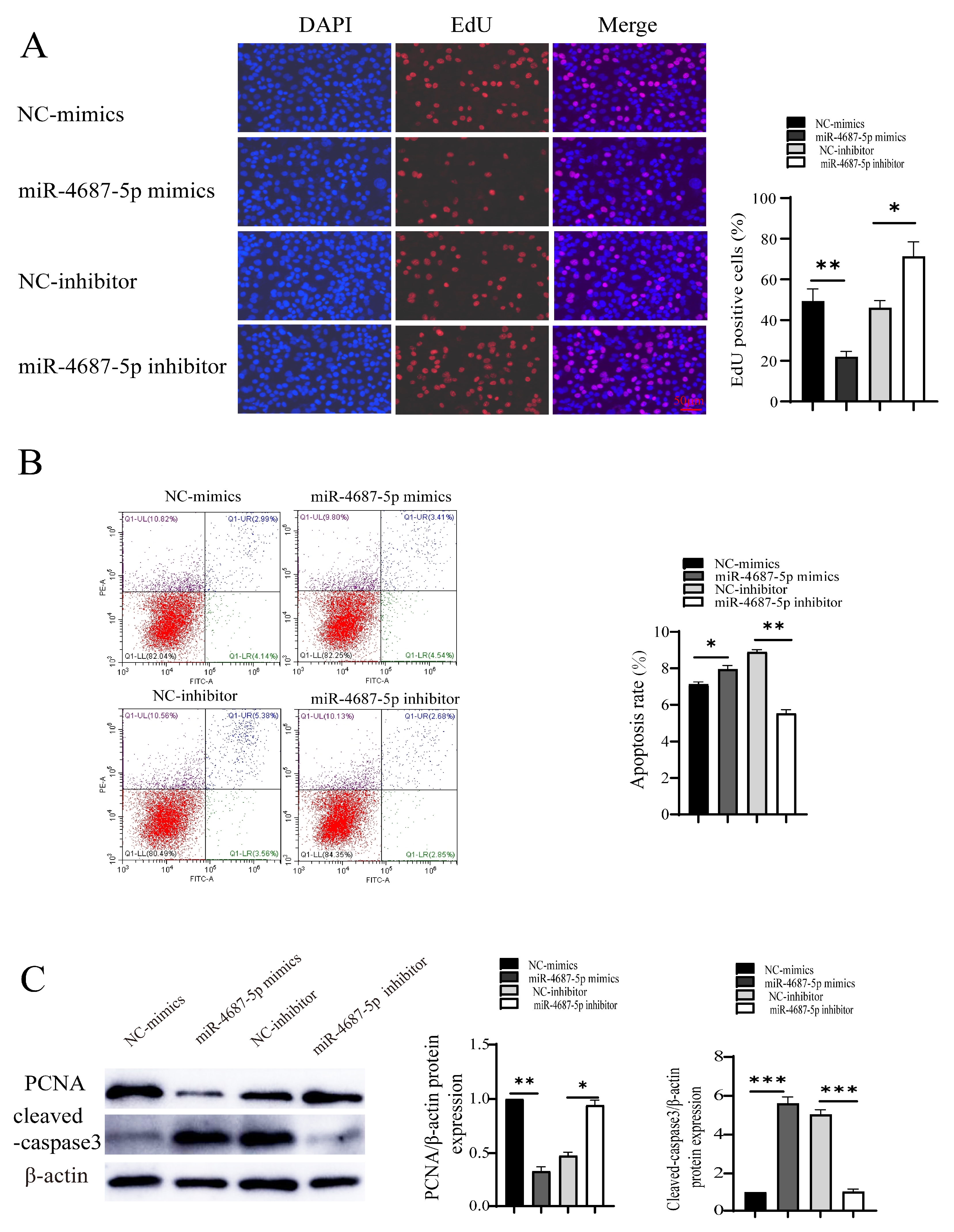
3.4. The Role of miR-4687-5p/NRAMP1 on Cell Proliferation and Apoptosis Post Mtb Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADC | acid albumin dextrose catalase |
| ANOVA | analysis of variance |
| BSA | bovine serum albumin |
| CFU | colony forming units |
| DR-TB | drug-resistant TB |
| EdU | 5-ethynyl-2′-deoxyuridine |
| FBS | fetal bovine serum |
| HRP | horse radish peroxidase |
| Mtb | Mycobacterium tuberculosis |
| MOI | multiplicity of infection |
| NC | negative control |
| NT | non-treated |
| NRAMP1 | natural resistance-associated macrophage protein 1 |
| PCNA | proliferating cell nuclear antigen |
| PFA | paraformaldehyde |
| PVDF | polyvinylidene fluoride |
| qRT-PCR | quantitative reverse transcription-PCR |
| RIPA | radio immunoprecipitation assay |
| SEM | standard error of mean |
| SLC11 | solute carrier family 11 |
| SNP | the single nucleotide polymorphism |
| TB | tuberculosis |
| UTR | untranslated region |
References
- Bagcchi, S. WHO’s Global Tuberculosis Report 2022. Lancet Microbe 2023, 4, e20. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Baltimore, D. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, D.; Boldin, M.P.; O’Connell, R.M.; Rao, D.S.; Taganov, K.D. MicroRNAs: New regulators of immune cell development and function. Nat. Immunol. 2008, 9, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Julie Li, Y.S.; Huang, H.D.; Shyy, J.Y.; Chien, S. microRNA: A master regulator of cellular processes for bioengineering systems. Annu. Rev. Biomed. Eng. 2010, 12, 1–27. [Google Scholar] [CrossRef]
- Ivey, K.N.; Srivastava, D. microRNAs as Developmental Regulators. Cold Spring Harb. Perspect. Biol. 2015, 7, a008144. [Google Scholar] [CrossRef]
- Li, H.; Yuan, J.; Duan, S.; Pang, Y. Resistance and tolerance of Mycobacterium tuberculosis to antimicrobial agents-How M. tuberculosis can escape antibiotics. WIREs Mech. Dis. 2022, 14, e1573. [Google Scholar] [CrossRef]
- Chandan, K.; Gupta, M.; Sarwat, M. Role of Host and Pathogen-Derived MicroRNAs in Immune Regulation During Infectious and Inflammatory Diseases. Front. Immunol. 2019, 10, 3081. [Google Scholar] [CrossRef]
- Jumat, M.I.; Sarmiento, M.E.; Acosta, A.; Chin, K.L. Role of non-coding RNAs in tuberculosis and their potential for clinical applications. J. Appl. Microbiol. 2023, 134, lxad104. [Google Scholar] [CrossRef]
- Yang, T.; Ge, B. miRNAs in immune responses to Mycobacterium tuberculosis infection. Cancer Lett. 2018, 431, 22–30. [Google Scholar] [CrossRef]
- Cellier, M.; Govoni, G.; Vidal, S.; Kwan, T.; Groulx, N.; Liu, J.; Sanchez, F.; Skamene, E.; Schurr, E.; Gros, P. Human natural resistance-associated macrophage protein: cDNA cloning, chromosomal mapping, genomic organization, and tissue-specific expression. J. Exp. Med. 1994, 180, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Gros, P.; Skamene, E. Natural resistance to infection with intracellular parasites: Molecular genetics identifies Nramp1 as the Bcg/Ity/Lsh locus. J. Leukoc. Biol. 1995, 58, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Canonne-Hergaux, F.; Calafat, J.; Richer, E.; Cellier, M.; Grinstein, S.; Borregaard, N.; Gros, P. Expression and subcellular localization of NRAMP1 in human neutrophil granules. Blood 2002, 100, 268–275. [Google Scholar] [CrossRef]
- Wyllie, S.; Seu, P.; Goss, J.A. The natural resistance-associated macrophage protein 1 Slc11a1 (formerly Nramp1) and iron metabolism in macrophages. Microbes Infect. 2002, 4, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, P.P. Host genetics and tuberculosis: Theory of genetic polymorphism and tuberculosis. Lung India Off. Organ Indian Chest Soc. 2019, 36, 244–252. [Google Scholar]
- Shahzad, F.; Bashir, N.; Ali, A.; Nadeem, A.; Ammar, A.; Kashif, M.; Javaid, K.; Jahan, S.; Tahir, R.; Rizwan, M.; et al. SLC11A1 genetic variation and low expression may cause immune response impairment in TB patients. Genes Immun. 2022, 23, 85–92. [Google Scholar] [CrossRef]
- Panio, A.; Cava, C.; D’Antona, S.; Bertoli, G.; Porro, D. Diagnostic Circulating miRNAs in Sporadic Amyotrophic Lateral Sclerosis. Front. Med. 2022, 9, 861960. [Google Scholar] [CrossRef]
- Floyd, K.; Glaziou, P.; Zumla, A.; Raviglione, M. The global tuberculosis epidemic and progress in care, prevention, and research: An overview in year 3 of the End TB era. Lancet Respir. Med. 2018, 6, 299–314. [Google Scholar] [CrossRef]
- Thye, T.; Owusu-Dabo, E.; Vannberg, F.O.; van Crevel, R.; Curtis, J.; Sahiratmadja, E.; Balabanova, Y.; Ehmen, C.; Muntau, B.; Ruge, G.; et al. Common variants at 11p13 are associated with susceptibility to tuberculosis. Nat. Genet. 2012, 44, 257–259. [Google Scholar] [CrossRef]
- Sobota, R.S.; Stein, C.M.; Kodaman, N.; Scheinfeldt, L.B.; Maro, I.; Wieland-Alter, W.; Igo, R.P., Jr.; Magohe, A.; Malone, L.L.; Chervenak, K.; et al. A Locus at 5q33.3 Confers Resistance to Tuberculosis in Highly Susceptible Individuals. Am. J. Hum. Genet. 2016, 98, 514–524. [Google Scholar] [CrossRef]
- Bermudez, L.E.; Goodman, J. Mycobacterium tuberculosis invades and replicates within type II alveolar cells. Infect. Immun. 1996, 64, 1400–1406. [Google Scholar] [CrossRef]
- Lerner, T.R.; de Souza Carvalho-Wodarz, C.; Repnik, U.; Russell, M.R.; Borel, S.; Diedrich, C.R.; Rohde, M.; Wainwright, H.; Collinson, L.M.; Wilkinson, R.J.; et al. Lymphatic endothelial cells are a replicative niche for Mycobacterium tuberculosis. J. Clin. Investig. 2016, 126, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Song, O.R.; Queval, C.J.; Iantomasi, R.; Delorme, V.; Marion, S.; Veyron-Churlet, R.; Werkmeister, E.; Popoff, M.; Ricard, I.; Jouny, S.; et al. ArfGAP1 restricts Mycobacterium tuberculosis entry by controlling the actin cytoskeleton. EMBO Rep. 2018, 19, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.H.; Shin, D.M.; Kang, G.; Kim, K.H.; Park, J.B.; Hur, G.M.; Lee, H.M.; Lim, Y.J.; Park, J.K.; Jo, E.K.; et al. Endoplasmic reticulum stress response is involved in Mycobacterium tuberculosis protein ESAT-6-mediated apoptosis. FEBS Lett. 2010, 584, 2445–2454. [Google Scholar] [CrossRef] [PubMed]
- Lutay, N.; Håkansson, G.; Alaridah, N.; Hallgren, O.; Westergren-Thorsson, G.; Godaly, G. Mycobacteria bypass mucosal NF-kB signalling to induce an epithelial anti-inflammatory IL-22 and IL-10 response. PLoS ONE 2014, 9, e86466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar]
- Angria, N.; Massi, M.N.; Bukhari, A.; Djaharuddin, I.; Jumadi, O.; Ahmad, A.; Miskad, U.A.; Ladju, R.B.; Santoso, A.; Halik, H. Expression of miRNA-29a-3p and IFN-γ as biomarkers in active and latent pulmonary tuberculosis. Ann. Med. Surg. 2022, 83, 104786. [Google Scholar] [CrossRef]
- Ebert, P.J.; Jiang, S.; Xie, J.; Li, Q.J.; Davis, M.M. An endogenous positively selecting peptide enhances mature T cell responses and becomes an autoantigen in the absence of microRNA miR-181a. Nat. Immunol. 2009, 10, 1162–1169. [Google Scholar] [CrossRef]
- Iwai, N.; Naraba, H. Polymorphisms in human pre-miRNAs. Biochem. Biophys. Res. Commun. 2005, 331, 1439–1444. [Google Scholar] [CrossRef]
- Amila, A.; Acosta, A.; Sarmiento, M.E.; Suraiya, S.; Zafarina, Z.; Panneerchelvam, S.; Norazmi, M.N. Sequence comparison of six human microRNAs genes between tuberculosis patients and healthy individuals. Int. J. Mycobacteriology 2015, 4, 341–346. [Google Scholar] [CrossRef]
- Chen, W.W.; Wan, B.; Zhang, R.; Cao, W.; Liang, L.; Zhao, Y.L.; Chen, J.; Yue, J. Genetic Polymorphisms of miR-149 Associated with Susceptibility to Both Pulmonary and Extrapulmonary Tuberculosis. Genet. Test. Mol. Biomark. 2019, 23, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Furci, L.; Schena, E.; Miotto, P.; Cirillo, D.M. Alteration of human macrophages microRNA expression profile upon infection with Mycobacterium tuberculosis. Int. J. Mycobacteriology 2013, 2, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, Z.; Sun, H.; Xin, H. Evaluation of Exosomal miRNA in Blood as a Potential Diagnostic Biomarker for Human Non-Small Cell Lung Cancer. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e924721. [Google Scholar] [CrossRef]
- Yuan, L.; Ke, Z.; Guo, Y.; Xi, X.; Luo, Z. NRAMP1 D543N and INT4 polymorphisms in susceptibility to pulmonary tuberculosis: A meta-analysis. Infect. Genet. Evol. 2017, 54, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Deng, H.; Zheng, Y.; Mansjö, M.; Zheng, X.; Hu, Y.; Xu, B. An association study of NRAMP1, VDR, MBL and their interaction with the susceptibility to tuberculosis in a Chinese population. Int. J. Infect. Dis. 2015, 38, 129–135. [Google Scholar] [CrossRef]
- Canonne-Hergaux, F.; Gruenheid, S.; Govoni, G.; Gros, P. The Nramp1 protein and its role in resistance to infection and macrophage function. Proc. Assoc. Am. Physicians 1999, 111, 283–289. [Google Scholar] [CrossRef]
- Green, D.R.; Llambi, F. Cell Death Signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Aguilo, J.I.; Alonso, H.; Uranga, S.; Marinova, D.; Arbués, A.; de Martino, A.; Anel, A.; Monzon, M.; Badiola, J.; Pardo, J.; et al. ESX-1-induced apoptosis is involved in cell-to-cell spread of Mycobacterium tuberculosis. Cell. Microbiol. 2013, 15, 1994–2005. [Google Scholar] [CrossRef]
- Lerner, T.R.; Borel, S.; Greenwood, D.J.; Repnik, U.; Russell, M.R.; Herbst, S.; Jones, M.L.; Collinson, L.M.; Griffiths, G.; Gutierrez, M.G. Mycobacterium tuberculosis replicates within necrotic human macrophages. J. Cell Biol. 2017, 216, 583–594. [Google Scholar] [CrossRef]
- Behar, S.M.; Martin, C.J.; Booty, M.G.; Nishimura, T.; Zhao, X.; Gan, H.X.; Divangahi, M.; Remold, H.G. Apoptosis is an innate defense function of macrophages against Mycobacterium tuberculosis. Mucosal Immunol. 2011, 4, 279–287. [Google Scholar] [CrossRef]
- Keane, J.; Balcewicz-Sablinska, M.K.; Remold, H.G.; Chupp, G.L.; Meek, B.B.; Fenton, M.J.; Kornfeld, H. Infection by Mycobacterium tuberculosis promotes human alveolar macrophage apoptosis. Infect. Immun. 1997, 65, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Seimon, T.A.; Kim, M.J.; Blumenthal, A.; Koo, J.; Ehrt, S.; Wainwright, H.; Bekker, L.G.; Kaplan, G.; Nathan, C.; Tabas, I.; et al. Induction of ER stress in macrophages of tuberculosis granulomas. PLoS ONE 2010, 5, e12772. [Google Scholar] [CrossRef] [PubMed]
- Aporta, A.; Arbues, A.; Aguilo, J.I.; Monzon, M.; Badiola, J.J.; de Martino, A.; Ferrer, N.; Marinova, D.; Anel, A.; Martin, C.; et al. Attenuated Mycobacterium tuberculosis SO2 vaccine candidate is unable to induce cell death. PLoS ONE 2012, 7, e45213. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, C.; Chen, G.; Liu, Y.; Wen, D.; Cui, J.; Dong, L.; Yang, Z.; Meng, H.; Gao, Y.; Feng, J.; et al. miR-4687-5p Affects Intracellular Survival of Mycobacterium tuberculosis through Its Regulation of NRAMP1 Expression in A549 Cells. Microorganisms 2024, 12, 227. https://doi.org/10.3390/microorganisms12010227
Meng C, Chen G, Liu Y, Wen D, Cui J, Dong L, Yang Z, Meng H, Gao Y, Feng J, et al. miR-4687-5p Affects Intracellular Survival of Mycobacterium tuberculosis through Its Regulation of NRAMP1 Expression in A549 Cells. Microorganisms. 2024; 12(1):227. https://doi.org/10.3390/microorganisms12010227
Chicago/Turabian StyleMeng, Chaoqun, Guangxin Chen, Yue Liu, Da Wen, Jia Cui, Li Dong, Zhiqiang Yang, Hangting Meng, Yuanting Gao, Jiao Feng, and et al. 2024. "miR-4687-5p Affects Intracellular Survival of Mycobacterium tuberculosis through Its Regulation of NRAMP1 Expression in A549 Cells" Microorganisms 12, no. 1: 227. https://doi.org/10.3390/microorganisms12010227
APA StyleMeng, C., Chen, G., Liu, Y., Wen, D., Cui, J., Dong, L., Yang, Z., Meng, H., Gao, Y., Feng, J., Cui, X., & Wu, C. (2024). miR-4687-5p Affects Intracellular Survival of Mycobacterium tuberculosis through Its Regulation of NRAMP1 Expression in A549 Cells. Microorganisms, 12(1), 227. https://doi.org/10.3390/microorganisms12010227