
Chlamydomonas reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism **



{kind=link}
{kind=link}
Abstract
1. Chlamydomonas reinhardtii as a Model in Mo Homeostasis
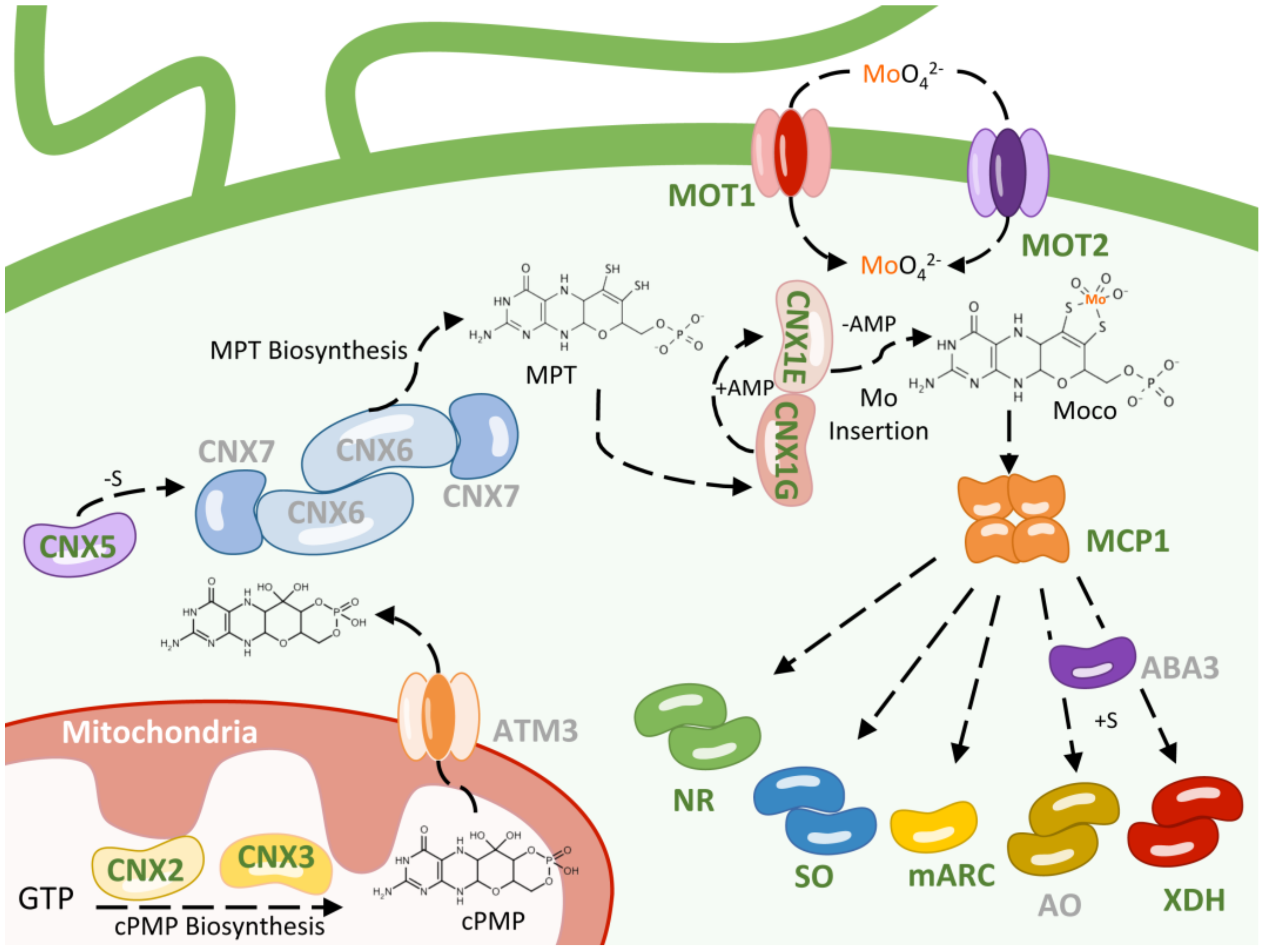
2. Molybdenum Uptake
3. Moco Biosynthesis
3.1. cPMP Biosynthesis
3.2. MPT Biosynthesis
3.3. Mo Insertion
4. Moco Storage
5. Chlamydomonas Moco Enzymes
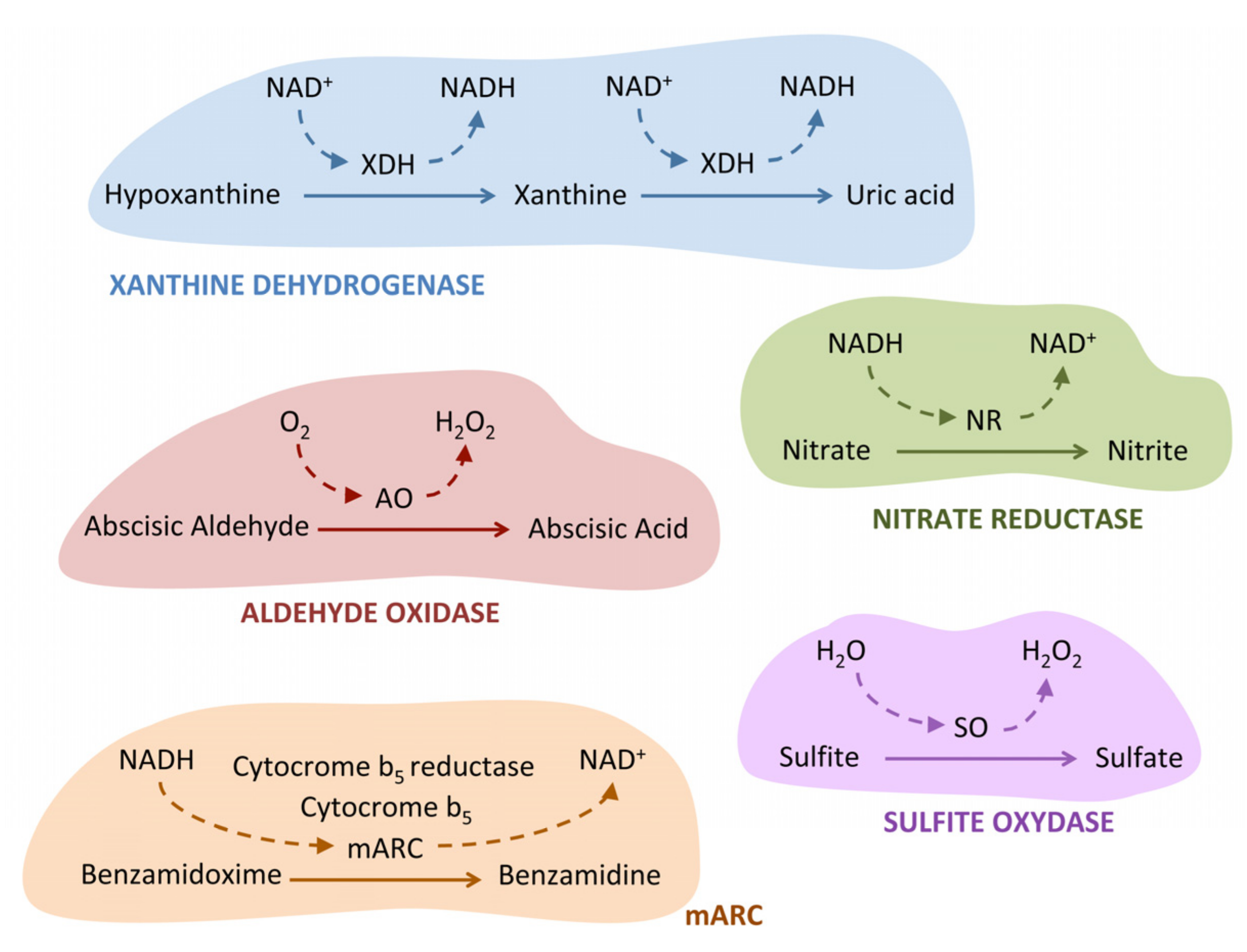
5.1. Xanthine Dehydrogenase
5.2. Aldehyde Oxidase
5.3. Sulfite Oxidase
5.4. Nitrate Reductase
5.5. mARC
6. Concluding Remarks and Future Outlook
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quintas-Nunes, F.; Brandão, P.R.; Barreto Crespo, M.T.; Glick, B.R.; Nascimento, F.X. Plant Growth Promotion, Phytohormone Production and Genomics of the Rhizosphere-Associated Microalga, Micractinium rhizosphaerae sp. nov. Plants 2023, 12, 651. [Google Scholar] [CrossRef] [PubMed]
- Maire, J.; Buerger, P.; Chan, W.Y.; Deore, P.; Dungan, A.M.; Nitschke, M.R.; van Oppen, M.J.H. Effects of Ocean Warming on the Underexplored Members of the Coral Microbiome. Integr. Comp. Biol. 2022, 62, 1700–1709. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.; Grube, M.; Schiefelbein, U.; Zühlke, D.; Bernhardt, J.; Riedel, K. The Lichens’ Microbiota, Still a Mystery? Front. Microbiol. 2021, 12, 623839. [Google Scholar] [CrossRef] [PubMed]
- Cvetkovska, M.; Vakulenko, G.; Smith, D.R.; Zhang, X.; Hüner, N.P.A. Temperature Stress in Psychrophilic Green Microalgae: Minireview. Physiol. Plant. 2022, 174, e13811. [Google Scholar] [CrossRef]
- Falkowski, P.G. The Role of Phytoplankton Photosynthesis in Global Biogeochemical Cycles. Photosynth. Res. 1994, 39, 235–258. [Google Scholar] [CrossRef]
- Tarafdar, A.; Sowmya, G.; Yogeshwari, K.; Rattu, G.; Negi, T.; Awasthi, M.K.; Hoang, A.T.; Sirohi, R. Environmental Pollution Mitigation through Utilization of Carbon Dioxide by Microalgae. Environ. Pollut. 2023, 328, 121623. [Google Scholar] [CrossRef]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived Intensification in Harmful Algal Blooms Is a Wave of Cumulative Threat to the Aquatic Ecosystems. Biology 2022, 11, 852. [Google Scholar] [CrossRef]
- Gerotto, C.; Norici, A.; Giordano, M. Toward Enhanced Fixation of CO2 in Aquatic Biomass: Focus on Microalgae. Front. Energy Res. 2020, 8, 213. [Google Scholar] [CrossRef]
- Parmar, P.; Kumar, R.; Neha, Y.; Srivatsan, V. Microalgae as next Generation Plant Growth Additives: Functions, Applications, Challenges and Circular Bioeconomy Based Solutions. Front. Plant Sci. 2023, 14, 1073546. [Google Scholar] [CrossRef]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.H. Microalgal-Bacterial Consortia: From Interspecies Interactions to Biotechnological Applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- Harris, E.H. Introduction into Chlamydomonas and Its Laboratory Use. Chlamydomonas Sourcebook; Oxford Academic Press: Oxford, UK, 2009. [Google Scholar]
- Salomé, P.A.; Merchant, S.S. A Series of Fortunate Events: Introducing Chlamydomonas as a Reference Organism. Plant Cell 2019, 31, 1682–1707. [Google Scholar] [CrossRef]
- Llamas, A.; Tejada-Jiménez, M.; Fernández, E.; Galván, A. Molybdenum Metabolism in the Alga Chlamydomonas Stands at the Crossroad of Those in Arabidopsis and Humans. Metallomics 2011, 3, 578–590. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef]
- Fauser, F.; Vilarrasa-Blasi, J.; Onishi, M.; Ramundo, S.; Patena, W.; Millican, M.; Osaki, J.; Philp, C.; Nemeth, M.; Salomamp, P.A.; et al. Systematic Characterization of Gene Function in the Photosynthetic Alga Chlamydomonas reinhardtii. Nat. Genet. 2022, 54, 705–714. [Google Scholar] [CrossRef]
- Goodenough, U. The Chlamydomonas Sourcebook. Volume 1: Introduction to Chlamydomonas and Its Laboratory Use; Elsevier Academic Press: Cambridge, MA, USA, 2023. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Sanz-Luque, E.; Tejada-Jiménez, M.; Llamas, A.; Plouviez, M.; Guieysse, B.; Fernández, E.; Galván, A. Chlamydomonas reinhardtii, an Algal Model in the Nitrogen Cycle. Plants 2020, 9, 903. [Google Scholar] [CrossRef]
- Saroussi, S.; Sanz-Luque, E.; Kim, R.G.; Grossman, A.R. Nutrient Scavenging and Energy Management: Acclimation Responses in Nitrogen and Sulfur Deprived Chlamydomonas. Curr. Opin. Plant Biol. 2017, 39, 114–122. [Google Scholar] [CrossRef]
- Irihimovitch, V.; Yehudai-Resheff, S. Phosphate and Sulfur Limitation Responses in the Chloroplast of Chlamydomonas reinhardtii. FEMS Microbiol. Lett. 2008, 283, 1–8. [Google Scholar] [CrossRef]
- Calatrava, V.; Hom, E.F.Y.; Llamas, A.; Fernández, E.; Galvan, A. Nitrogen Scavenging from Amino Acids and Peptides in the Model Alga Chlamydomonas reinhardtii. The Role of Extracellular L-Amino Oxidase. Algal Res. 2019, 38, 101395. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Kong, F.; Wang, P.; Lee, Y.; Kang, B.-H. The Disassembly of Lipid Droplets in Chlamydomonas. New Phytol. 2021, 231, 1359–1364. [Google Scholar] [CrossRef]
- Rengel, R.; Giraldez, I.; Díaz, M.J.; García, T.; Vigara, J.; Leon, R. Simultaneous Production of Carotenoids and Chemical Building Blocks Precursors from Chlorophyta Microalgae. Bioresour. Technol. 2022, 351, 127035. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, R.; Joun, J.; Choi, H.I.; Gaur, V.K.; Sim, S.J. Algal Glycobiotechnology: Omics Approaches for Strain Improvement. Microb. Cell Fact. 2021, 20, 163. [Google Scholar] [CrossRef] [PubMed]
- Calderon, R.H.; de Vitry, C.; Wollman, F.A.; Niyogi, K.K. Rubredoxin 1 Promotes the Proper Folding of D1 and Is Not Required for Heme B559 Assembly in Chlamydomonas Photosystem II. J. Biol. Chem. 2023, 299, 102968. [Google Scholar] [CrossRef] [PubMed]
- Przybyla-Toscano, J.; Couturier, J.; Remacle, C.; Rouhier, N. Occurrence, Evolution and Specificities of Iron-Sulfur Proteins and Maturation Factors in Chloroplasts from Algae. Int. J. Mol. Sci. 2021, 22, 3175. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Baur, T.; Kranner, I. β-Cyclocitral Does Not Contribute to Singlet Oxygen-Signalling in Algae, but May Down-Regulate Chlorophyll Synthesis. Plants 2022, 11, 2155. [Google Scholar] [CrossRef]
- Yang, M.; Xie, X.; Kong, F.T.; Xie, K.P.; Yu, S.H.; Ma, J.Y.; Xue, S.; Gong, Z. Differences in Glycerolipid Response of Chlamydomonas reinhardtii Starchless Mutant to High Light and Nitrogen Deprivation Stress under Three Carbon Supply Regimes. Front. Plant Sci. 2022, 13, 860966. [Google Scholar] [CrossRef]
- Kreis, E.; Niemeyer, J.; Merz, M.; Scheuring, D.; Schroda, M. CLPB3 Is Required for the Removal of Chloroplast Protein Aggregates and Thermotolerance in Chlamydomonas. J. Exp. Bot. 2023, erad109. [Google Scholar] [CrossRef]
- Zou, Y.; Bozhkov, P.V. Chlamydomonas Proteases: Classification, Phylogeny, and Molecular Mechanisms. J. Exp. Bot. 2021, 72, 7680–7693. [Google Scholar] [CrossRef]
- Marchetti, G.M.; Füsser, F.; Singh, R.K.; Brummel, M.; Koch, O.; Kümmel, D.; Hippler, M. Structural Analysis Revealed a Novel Conformation of the NTRC Reductase Domain from Chlamydomonas reinhardtii. J. Struct. Biol. 2022, 214, 107829. [Google Scholar] [CrossRef]
- Marshall, W.F. The Flagellar Length Control System: Exploring the Physical Biology of Organelle Size. Phys. Biol. 2023, 20, 021001. [Google Scholar] [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From Molecular Manipulation of Domesticated Chlamydomonas reinhardtii to Survival in Nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Masi, A.; Leonelli, F.; Scognamiglio, V.; Gasperuzzo, G.; Antonacci, A.; Terzidis, M.A. Chlamydomonas reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health. Molecules 2023, 28, 1185. [Google Scholar] [CrossRef]
- Dehghani, J.; Movafeghi, A.; Mathieu-Rivet, E.; Mati-Baouche, N.; Calbo, S.; Lerouge, P.; Bardor, M. Microalgae as an Efficient Vehicle for the Production and Targeted Delivery of Therapeutic Glycoproteins against SARS-CoV-2 Variants. Mar. Drugs 2022, 20, 657. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The Potential of Algal Biotechnology to Produce Antiviral Compounds and Biopharmaceuticals. Molecules 2020, 25, 4049. [Google Scholar] [CrossRef]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the Microalgae Chlamydomonas on Gastrointestinal Health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Calatrava, V.; Tejada-Jimenez, M.; Sanz-Luque, E.; Fernandez, E.; Galvan, A.; Llamas, A. Chlamydomonas reinhardtii, a Reference Organism to Study Algal–Microbial Interactions: Why Can’t They Be Friends? Plants 2023, 12, 788. [Google Scholar] [CrossRef]
- Fakhimi, N.; Gonzalez-Ballester, D.; Fernández, E.; Galván, A.; Dubini, A. Algae-Bacteria Consortia as a Strategy to Enhance H2 Production. Cells 2020, 9, 1353. [Google Scholar] [CrossRef]
- Torres, M.; Gonzalez-Ballester, D.; Gomez-Osuna, A.; Galván, A.; Fernandez, E.; Dubini, A. Chlamydomonas-Methylobacterium oryzae Cooperation Leads to Increased Biomass, Nitrogen Removal, and Hydrogen Production. Bioresour. Technol. 2022, 352, 127088. [Google Scholar] [CrossRef]
- Kelterborn, S.; Boehning, F.; Sizova, I.; Baidukova, O.; Evers, H.; Hegemann, P. Gene Editing in Green Alga Chlamydomonas reinhardtii via CRISPR-Cas9 Ribonucleoproteins. Plant Synth. Biol. Methods Mol. Biol. 2022, 2379, 45–65. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Llamas, A.; Sanz-Luque, E.; Galván, A.; Fernández, E. A High-Affinity Molybdate Transporter in Eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 20126–20130. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Galván, A.; Fernández, E. Algae and Humans Share a Molybdate Transporter. Proc. Natl. Acad. Sci. USA 2011, 108, 6420–6425. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Llamas, A.; Tejada-Jiménez, M.; Schrader, N.; Kuper, J.; Ataya, F.S.; Galván, A.; Mendel, R.R.; Fernández, E.; Schwarz, G. Function and Structure of the Molybdenum Cofactor Carrier Protein from Chlamydomonas reinhardtii. J. Biol. Chem. 2006, 281, 30186–30194. [Google Scholar] [CrossRef] [PubMed]
- Ataya, F.S.; Witte, C.P.; Galván, A.; Igeño, M.I.; Fernández, E. Mcp1 Encodes the Molybdenum Cofactor Carrier Protein in Chlamydomonas reinhardtii and Participates in Protection, Binding, and Storage Functions of the Cofactor. J. Biol. Chem. 2003, 278, 10885–10890. [Google Scholar] [CrossRef] [PubMed]
- Llamas, A.; Tejada-Jimenez, M.; González-Ballester, D.; Higuera, J.J.; Schwarz, G.; Galván, A.; Fernández, E. Chlamydomonas Reinhardtii CNX1E Reconstitutes Molybdenum Cofactor Biosynthesis in Escherichia coli Mutants. Eukaryot. Cell 2007, 6, 1063–1067. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galván, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Ocaña-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galván, A.; Fernández, E. A Dual System Formed by the ARC and NR Molybdoenzymes Mediates Nitrite-Dependent NO Production in Chlamydomonas. Plant. Cell Environ. 2016, 39, 2097–2107. [Google Scholar] [CrossRef]
- Emsley, J. Nature’s Building Blocks; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Lešková, A.; Javot, H.; Giehl, R.F.H. Metal Crossroads in Plants: Modulation of Nutrient Acquisition and Root Development by Essential Trace Metals. J. Exp. Bot. 2022, 73, 1751–1765. [Google Scholar] [CrossRef]
- Llamas, A.; Kalakoutskii, K.L.; Fernández, E. Molybdenum Cofactor Amounts in Chlamydomonas reinhardtii Depend on the Nit5 Gene Function Related to Molybdate Transport. Plant Cell Environ. 2000, 23, 1247–1255. [Google Scholar] [CrossRef]
- Mendel, R.R.; Haensh, R. Molybdoenzymes and Molybdenum Cofactor in Plants. J. Exp. Bot. 2002, 53, 1689–1698. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Characterization of Chlamydomonas 102 and 104 Mutants Reveals Intermolecular Complementation in the Molybdenum Cofactor Protein CNX1E. Protist 2013, 164, 116–128. [Google Scholar] [CrossRef]
- Schwarz, G.; Schulze, J.; Bittner, F.; Eilers, T.; Kuper, J.; Bollmann, G.; Nerlich, A.; Brinkmann, H.; Mendel, R.R. The Molybdenum Cofactor Biosynthetic Protein Cnx1 Complements Molybdate-Repairable Mutants, Transfers Molybdenum to the Metal Binding Pterin, and Is Associated with the Cytoskeleton. Plant Cell 2000, 12, 2455–2471. [Google Scholar] [CrossRef]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis thaliana High-Affinity Molybdate Transporter Required for Efficient Uptake of Molybdate from Soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar] [CrossRef]
- Ide, Y.; Kusano, M.; Oikawa, A.; Fukushima, A.; Tomatsu, H.; Saito, K.; Hirai, M.Y.; Fujiwara, T. Effects of Molybdenum Deficiency and Defects in Molybdate Transporter MOT1 on Transcript Accumulation and Nitrogen/Sulphur Metabolism in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 1483–1497. [Google Scholar] [CrossRef]
- Gao, J.-S.; Wu, F.-F.; Shen, Z.-L.; Meng, Y.; Cai, Y.-P.; Lin, Y. A Putative Molybdate Transporter LjMOT1 Is Required for Molybdenum Transport in Lotus japonicus. Physiol. Plant. 2016, 158, 331–340. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Gil-Díez, P.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. Medicago truncatula Molybdate Transporter Type 1 (MtMOT1.3) Is a Plasma Membrane Molybdenum Transporter Required for Nitrogenase Activity in Root Nodules under Molybdenum Deficiency. New Phytol. 2017, 216, 1223–1235. [Google Scholar] [CrossRef]
- Gil-Díez, P.; Tejada-Jiménez, M.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. MtMOT1.2 Is Responsible for Molybdate Supply to Medicago truncatula Nodules. Plant. Cell Environ. 2019, 42, 310–320. [Google Scholar] [CrossRef]
- Forsberg, S.K.G.; Andreatta, M.E.; Huang, X.Y.; Danku, J.; Salt, D.E.; Carlborg, Ö. The Multi-Allelic Genetic Architecture of a Variance-Heterogeneity Locus for Molybdenum Concentration in Leaves Acts as a Source of Unexplained Additive Genetic Variance. PLoS Genet. 2015, 11, 1005648. [Google Scholar] [CrossRef]
- Li, W.; Fingrut, D.R.; Maxwell, D.P. Characterization of a Mutant of Chlamydomonas reinhardtii Deficient in the Molybdenum Cofactor. Physiol. Plant. 2009, 136, 336–350. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Calatrava, V.; Galván, A.; Fernández, E.; Llamas, A. From the Eukaryotic Molybdenum Cofactor Biosynthesis to the Moonlighting Enzyme MARC. Molecules 2018, 23, 3287. [Google Scholar] [CrossRef]
- Schwarz, G.; Mendel, R.R. Molybdenum Cofactor Biosynthesis and Molybdenum Enzymes. Annu. Rev. Plant Biol. 2006, 57, 623–647. [Google Scholar] [CrossRef]
- Kruse, I.; Maclean, A.E.; Hill, L.; Balk, J. Genetic Dissection of Cyclic Pyranopterin Monophosphate Biosynthesis in Plant Mitochondria. Biochem. J. 2018, 475, 495–509. [Google Scholar] [CrossRef] [PubMed]
- González-Ballester, D.; de Montaigu, A.; Higuera, J.J.; Galván, A.; Fernández, E. Functional Genomics of the Regulation of the Nitrate Assimilation Pathway in Chlamydomonas. Plant Physiol. 2005, 137, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. The Chlamydomonas reinhardtii Molybdenum Cofactor Enzyme CrARC Has a Zn-Dependent Activity and Protein Partners Similar to Those of Its Human Homologue. Eukaryot. Cell 2011, 10, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Teschner, J.; Lachmann, N.; Schulze, J.; Geisler, M.; Selbach, K.; Santamaria-Araujo, J.; Balk, J.; Mendel, R.R.; Bittner, F. A Novel Role for Arabidopsis Mitochondrial ABC Transporter ATM3 in Molybdenum Cofactor Biosynthesis. Plant Cell 2010, 22, 468–480. [Google Scholar] [CrossRef]
- Kaufholdt, D.; Baillie, C.K.; Meinen, R.; Mendel, R.R.; Hänsch, R. The Molybdenum Cofactor Biosynthesis Network: In Vivo Protein-Protein Interactions of an Actin Associated Multi-Protein Complex. Front. Plant Sci. 2017, 8, 1946. [Google Scholar] [CrossRef]
- Nakai, Y.; Harada, A.; Hashiguchi, Y.; Nakai, M.; Hayashi, H. Arabidopsis Molybdopterin Biosynthesis Protein Cnx5 Collaborates with the Ubiquitin-like Protein Urm11 in the Thio-Modification of TRNA. J. Biol. Chem. 2012, 287, 30874–30884. [Google Scholar] [CrossRef]
- Veldman, A.; Santamaria-Araujo, J.A.; Sollazzo, S.; Pitt, J.; Gianello, R.; Yaplito-Lee, J.; Wong, F.; Ramsden, C.A.; Reiss, J.; Cook, I.; et al. Successful Treatment of Molybdenum Cofactor Deficiency Type A with CPMP. Pediatrics 2010, 125, e1249-54. [Google Scholar] [CrossRef]
- Farrell, S.; Karp, J.; Hager, R.; Wang, Y.; Adeniyi, O.; Wang, J.; Li, L.; Ma, L.; Peretz, J.; Summan, M.; et al. Regulatory News: Nulibry (Fosdenopterin) Approved to Reduce the Risk of Mortality in Patients with Molybdenum Cofactor Deficiency Type A: FDA Approval Summary. J. Inherit. Metab. Dis. 2021, 44, 1085–1087. [Google Scholar] [CrossRef]
- Kuper, J.; Llamas, A.; Hecht, H.-J.; Mendel, R.R.; Schwarz, G. Structure of the Molybdopterin-Bound Cnx1G Domain Links Molybdenum and Copper Metabolism. Nature 2004, 430, 803–806. [Google Scholar] [CrossRef]
- Kruse, T. Function of Molybdenum Insertases. Molecules 2022, 27, 5372. [Google Scholar] [CrossRef]
- Llamas, A.; Mendel, R.R.; Schwarz, G. Synthesis of Adenylated Molybdopterin: An Essential Step for Molybdenum Insertion. J. Biol. Chem. 2004, 279, 55241–55246. [Google Scholar] [CrossRef]
- Llamas, A.; Otte, T.; Multhaup, G.; Mendel, R.R.; Schwarz, G. The Mechanism of Nucleotide-Assisted Molybdenum Insertion into Molybdopterin. J. Biol. Chem. 2006, 281, 18343–18350. [Google Scholar] [CrossRef]
- Nichols, J.D.; Rajagopalan, K.V. In Vitro Molybdenum Ligation to Molybdopterin Using Purified Components. J. Biol. Chem. 2005, 280, 7817–7822. [Google Scholar] [CrossRef]
- Selles, B.; Moseler, A.; Caubrière, D.; Sun, S.K.; Ziesel, M.; Dhalleine, T.; Hériché, M.; Wirtz, M.; Rouhier, N.; Couturier, J. The Cytosolic Arabidopsis thaliana Cysteine Desulfurase ABA3 Delivers Sulfur to the Sulfurtransferase STR18. J. Biol. Chem. 2022, 298, 101749. [Google Scholar] [CrossRef]
- Mendel, R.R. The History of the Molybdenum Cofactor-A Personal View. Molecules 2022, 27, 4934. [Google Scholar] [CrossRef]
- Aguilar, M.; Cárdenas, J.; Fernández, E. Quantitation of Molybdopterin Oxidation Product in Wild-Type and Molybdenum Cofactor Deficient Mutants of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 1992, 1160, 269–274. [Google Scholar] [CrossRef]
- Witte, C.-P.; Igeño, M.I.; Mendel, R.; Schwarz, G.; Fernández, E. The Chlamydomonas reinhardtii MoCo Carrier Protein Is Multimeric and Stabilizes Molybdopterin Cofactor in a Molybdate Charged Form. FEBS Lett. 1998, 431, 205–209. [Google Scholar] [CrossRef]
- Aguilar, M.; Kalakoutskii, K.; Cárdenas, J.; Fernández, E. Direct Transfer of Molybdopterin Cofactor to Aponitrate Reductase from a Carrier Protein in Chlamydomonas reinhardtii. FEBS Lett 1992, 307, 162–163. [Google Scholar] [CrossRef]
- Krausze, J.; Hercher, T.W.; Archna, A.; Kruse, T. The Structure of the Moco Carrier Protein from Rippkaea orientalis. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2020, 76, 453–463. [Google Scholar] [CrossRef]
- Hercher, T.W.; Krausze, J.; Yang, J.; Kirk, M.L.; Kruse, T. Identification and Characterization of the Volvox carteri Moco Carrier Protein. Biosci. Rep. 2020, 40, BSR20202351. [Google Scholar] [CrossRef]
- Kruse, T.; Gehl, C.; Geisler, M.; Lehrke, M.; Ringel, P.; Hallier, S.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Molybdenum Cofactor-Binding Proteins from Arabidopsis thaliana. J. Biol. Chem. 2010, 285, 6623–6635. [Google Scholar] [CrossRef] [PubMed]
- Kruse, T. Moco Carrier and Binding Proteins. Molecules 2022, 27, 6571. [Google Scholar] [CrossRef] [PubMed]
- Lawson, D.M.; Williams, C.E.; White, D.J.; Choay, A.P.; Mitchenall, L.A.; Pau, R.N. Protein Ligands for Molybdate. Specificity and Charge Stabilisation at the Anion-Binding Sites of Periplasmic and Intracellular Molybdate-Binding Proteins of Azotobacter vinelandii. J. Chem. Soc. Dalt. Trans. 1997, 21, 3981–3984. [Google Scholar] [CrossRef]
- Fenske, D.; Gnida, M.; Schneider, K.; Meyer-Klaucke, W.; Schemberg, J.; Henschel, V.; Meyer, A.-K.; Knöchel, A.; Müller, A. A New Type of Metalloprotein: The Mo Storage Protein from Azotobacter vinelandii Contains a Polynuclear Molybdenum-Oxide Cluster. ChemBioChem 2005, 6, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Steinke, D.R.; Majak, W.; Sorensen, T.S.; Parvez, M. Chelation of Molybdenum in Medicago Sativa (Alfalfa) Grown on Reclaimed Mine Tailings. J. Agric. Food Chem. 2008, 56, 5437–5442. [Google Scholar] [CrossRef]
- Hille, R. Xanthine Oxidase-A Personal History. Molecules 2023, 28, 1921. [Google Scholar] [CrossRef]
- Pineda, M.; Cardenas, J. Transport and Assimilation of Purines in Chlamydomonas reinhardtii. Sci. Mar. 1996, 60, 195–201. [Google Scholar]
- Schmollinger, S.; Mühlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing Mechanisms in Chlamydomonas Affect the Transcriptome, the Proteome, and Photosynthetic Metabolism. Plant Cell 2014, 26, 1410–1435. [Google Scholar] [CrossRef]
- Perez-Vicente, R.; Pineda, M.; Cardenas, J. Isolation and Characterization of Xanthine Dehydrogenase from Chlamydomonas reinhardtii. Physiol. Plant. 1988, 72, 101–107. [Google Scholar] [CrossRef]
- Seo, M.; Koiwai, H.; Akaba, S.; Komano, T.; Oritani, T.; Kamiya, Y.; Koshiba, T. Abscisic Aldehyde Oxidase in Leaves of Arabidopsis thaliana. Plant J. 2000, 23, 481–488. [Google Scholar] [CrossRef]
- Rodríguez-Trelles, F.; Tarrío, R.; Ayala, F.J. Convergent Neofunctionalization by Positive Darwinian Selection after Ancient Recurrent Duplications of the Xanthine Dehydrogenase Gene. Proc. Natl. Acad. Sci. USA 2003, 100, 13413–13417. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Iluz, D.; Dubinsky, Z.; Miller, G. Exogenous Abscisic Acid Confers Salinity Tolerance in Chlamydomonas reinhardtii During Its Life Cycle. J. Phycol. 2021, 57, 1323–1334. [Google Scholar] [CrossRef]
- Al-Hijab, L.; Gregg, A.; Davies, R.; Macdonald, H.; Ladomery, M.; Wilson, I. Abscisic Acid Induced a Negative Geotropic Response in Dark-Incubated Chlamydomonas reinhardtii. Sci. Rep. 2019, 9, 12063. [Google Scholar] [CrossRef]
- Feng, C.; Tollin, G.; Enemark, J.H. Sulfite Oxidizing Enzymes. Biochim. Biophys. Acta-Proteins Proteom. 2007, 1774, 527–539. [Google Scholar] [CrossRef]
- Kirk, M.L.; Hille, R. Spectroscopic Studies of Mononuclear Molybdenum Enzyme Centers. Molecules 2022, 27, 4802. [Google Scholar] [CrossRef]
- Randewig, D.; Hamisch, D.; Herschbach, C.; Eiblmeier, M.; Gehl, C.; Jurgeleit, J.; Skerra, J.; Mendel, R.R.; Rennenberg, H.; Hänsch, R. Sulfite Oxidase Controls Sulfur Metabolism under SO2 Exposure in Arabidopsis thaliana. Plant. Cell Environ. 2012, 35, 100–115. [Google Scholar] [CrossRef]
- Mellis, A.T.; Roeper, J.; Misko, A.L.; Kohl, J.; Schwarz, G. Sulfite Alters the Mitochondrial Network in Molybdenum Cofactor Deficiency. Front. Genet. 2021, 11, 594828. [Google Scholar] [CrossRef]
- Nowak, K.; Luniak, N.; Witt, C.; Wüstefeld, Y.; Wachter, A.; Mendel, R.R.; Hänsch, R. Peroxisomal Localization of Sulfite Oxidase Separates It from Chloroplast-Based Sulfur Assimilation. Plant Cell Physiol. 2004, 45, 1889–1894. [Google Scholar] [CrossRef]
- Eilers, T.; Schwarz, G.; Brinkmann, H.; Witt, C.; Richter, T.; Nieder, J.; Koch, B.; Hille, R.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Arabidopsis thaliana Sulfite Oxidase: A New Player in Plant Sulfur Metabolism. J. Biol. Chem. 2001, 276, 46989–46994. [Google Scholar] [CrossRef]
- Gerin, S.; Mathy, G.; Blomme, A.; Franck, F.; Sluse, F.E. Plasticity of the Mitoproteome to Nitrogen Sources (Nitrate and Ammonium) in Chlamydomonas reinhardtii: The Logic of Aox1 Gene Localization. Biochim. Biophys. Acta-Bioenerg. 2010, 1797, 994–1003. [Google Scholar] [CrossRef]
- Coelho, C.; Romão, M.J. Structural and Mechanistic Insights on Nitrate Reductases. Protein Sci. 2015, 24, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Schnell, R.; Ranum, L.P.; Hussey, S.C.; Silflow, C.D.; Lefebvre, P.A. Isolation and Characterization of the Nitrate Reductase Structural Gene of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1989, 86, 6449–6453. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty Years from Transport to Signaling Networks. Plant Cell 2020, 32, 2094–2119. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.V.; Harper, J.E. The Conversion of Nitrite to Nitrogen Oxide(s) by the Constitutive NAD(P)H-Nitrate Reductase Enzyme from Soybean. Plant Physiol. 1988, 88, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Yun, B.W. Nitric Oxide Acts as a Key Signaling Molecule in Plant Development under Stressful Conditions. Int. J. Mol. Sci. 2023, 24, 4782. [Google Scholar] [CrossRef]
- Gupta, K.J.; Kaladhar, V.C.; Fitzpatrick, T.B.; Fernie, A.R.; Møller, I.M.; Loake, G.J. Nitric Oxide Regulation of Plant Metabolism. Mol. Plant 2022, 15, 228–242. [Google Scholar] [CrossRef]
- Foresi, N.; Correa-Aragunde, N.; Parisi, G.; Caló, G.; Salerno, G.; Lamattina, L. Characterization of a Nitric Oxide Synthase from the Plant Kingdom: NO Generation from the Green Alga Ostreococcus tauri Is Light Irradiance and Growth Phase Dependent. Plant Cell 2010, 22, 3816–3830. [Google Scholar] [CrossRef]
- Santolini, J.; André, F.; Jeandroz, S.; Wendehenne, D. Nitric Oxide Synthase in Plants: Where Do We Stand? Nitric Oxide 2017, 63, 30–38. [Google Scholar] [CrossRef]
- Havemeyer, A.; Bittner, F.; Wollers, S.; Mendel, R.; Kunze, T.; Clement, B. Identification of the Missing Component in the Mitochondrial Benzamidoxime Prodrug-Converting System as a Novel Molybdenum Enzyme. J. Biol. Chem. 2006, 281, 34796–34802. [Google Scholar] [CrossRef]
- Hsieh, S.I.; Castruita, M.; Malasarn, D.; Urzica, E.; Erde, J.; Page, M.D.; Yamasaki, H.; Casero, D.; Pellegrini, M.; Merchant, S.S.; et al. The Proteome of Copper, Iron, Zinc, and Manganese Micronutrient Deficiency in Chlamydomonas reinhardtii. Mol. Cell. Proteom. 2013, 12, 65–86. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Study of Different Variants of Mo Enzyme CrARC and the Interaction with Its Partners CrCytb5-R and CrCytb5-1. Int. J. Mol. Sci. 2017, 18, 670. [Google Scholar] [CrossRef]
- Ott, G.; Plitzko, B.; Krischkowski, C.; Reichmann, D.; Bittner, F.; Mendel, R.R.; Kunze, T.; Clement, B.; Havemeyer, A. Reduction of Sulfamethoxazole Hydroxylamine (SMX-HA) by the Mitochondrial Amidoxime Reducing Component (MARC). Chem. Res. Toxicol. 2014, 27, 1687–1695. [Google Scholar] [CrossRef]
- Kotthaus, J.; Wahl, B.; Havemeyer, A.; Schade, D.; Garbe-Schönberg, D.; Mendel, R.; Bittner, F.; Clement, B. Reduction of N(ω)-Hydroxy-L-Arginine by the Mitochondrial Amidoxime Reducing Component (MARC). Biochem. J. 2011, 433, 383–391. [Google Scholar] [CrossRef]
- Sparacino-Watkins, C.E.; Tejero, J.; Sun, B.; Gauthier, M.C.; Thomas, J.; Ragireddy, V.; Merchant, B.A.; Wang, J.; Azarov, I.; Basu, P.; et al. Nitrite Reductase and Nitric-Oxide Synthase Activity of the Mitochondrial Molybdopterin Enzymes MARC1 and MARC2. J. Biol. Chem. 2014, 289, 10345–10358. [Google Scholar] [CrossRef]
- Bender, D.; Schwarz, G. Nitrite-Dependent Nitric Oxide Synthesis by Molybdenum Enzymes. FEBS Lett. 2018, 592, 2126–2139. [Google Scholar] [CrossRef]
- Rockel, P.; Strube, F.; Rockel, A.; Wildt, J.; Kaiser, W.M. Regulation of Nitric Oxide (NO) Production by Plant Nitrate Reductase In Vivo and In Vitro. J. Exp. Bot. 2002, 53, 103–110. [Google Scholar] [CrossRef]
- Maiber, L.; Koprivova, A.; Bender, D.; Kopriva, S.; Fischer-Schrader, K. Characterization of the Amidoxime Reducing Components ARC1 and ARC2 from Arabidopsis thaliana. FEBS J. 2022, 289, 5656–5669. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Llamas, A.; Fernandez, E.; Sanz-Luque, E.; Galvan, A. Nitrous Oxide Emissions from Nitrite Are Highly Dependent on Nitrate Reductase in the Microalga Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2022, 23, 9412. [Google Scholar] [CrossRef]
- Llamas, A.; Chamizo-Ampudia, A.; Tejada-Jimenez, M.; Galvan, A.; Fernandez, E. The Molybdenum Cofactor Enzyme MARC: Moonlighting or Promiscuous Enzyme? BioFactors 2017, 43, 486–494. [Google Scholar] [CrossRef]
- Gupta, M.N.; Uversky, V.N. Moonlighting Enzymes: When Cellular Context Defines Specificity. Cell. Mol. Life Sci. 2023, 80, 130. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejada-Jimenez, M.; Leon-Miranda, E.; Llamas, A. Chlamydomonas reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism. Microorganisms 2023, 11, 1671. https://doi.org/10.3390/microorganisms11071671
Tejada-Jimenez M, Leon-Miranda E, Llamas A. Chlamydomonas reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism. Microorganisms. 2023; 11(7):1671. https://doi.org/10.3390/microorganisms11071671
Chicago/Turabian StyleTejada-Jimenez, Manuel, Esperanza Leon-Miranda, and Angel Llamas. 2023. "Chlamydomonas reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism" Microorganisms 11, no. 7: 1671. https://doi.org/10.3390/microorganisms11071671
APA StyleTejada-Jimenez, M., Leon-Miranda, E., & Llamas, A. (2023). Chlamydomonas reinhardtii—A Reference Microorganism for Eukaryotic Molybdenum Metabolism. Microorganisms, 11(7), 1671. https://doi.org/10.3390/microorganisms11071671