

SARS-CoV-2 Rapid Antigen Test Based on a New Anti-Nucleocapsid Protein Monoclonal Antibody: Development and Real-Time Validation
 , , , , , ,
, , , , , ,  , , , ,
, , , ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Populational Study
2.3. Follow-Up Study
2.3.1. Sampling
2.3.2. RT-qPCR and RNA Sequencing
2.4. Expression and Purification of Nucleocapsid Recombinant Proteins DTC-N, N-Terminal Domain (Residues 47–177), and C-terminal Domain (Residues 210–419)
2.5. SARS-CoV-2 Nucleocapsid Monoclonal Antibody (mAb) Production and Characterization
2.5.1. mAb Sequencing
2.5.2. Epitope Characterization and Structure Analysis
2.6. SARS-CoV-2 Samples
2.7. Antigen Rapid Diagnostic Test (Ag-RDT)
2.7.1. Inactivation Buffer
2.7.2. Prototype 1
2.7.3. Prototype 2
2.7.4. Prototype 3
2.8. Assaying Ag-RDT Prototypes with SARS-CoV-2 Virus
2.9. Assaying Ag-RDT Prototypes with Patients’ Samples for Validation
2.10. Stability Assay
2.11. Statistical Analysis
3. Results
3.1. Population Characteristics
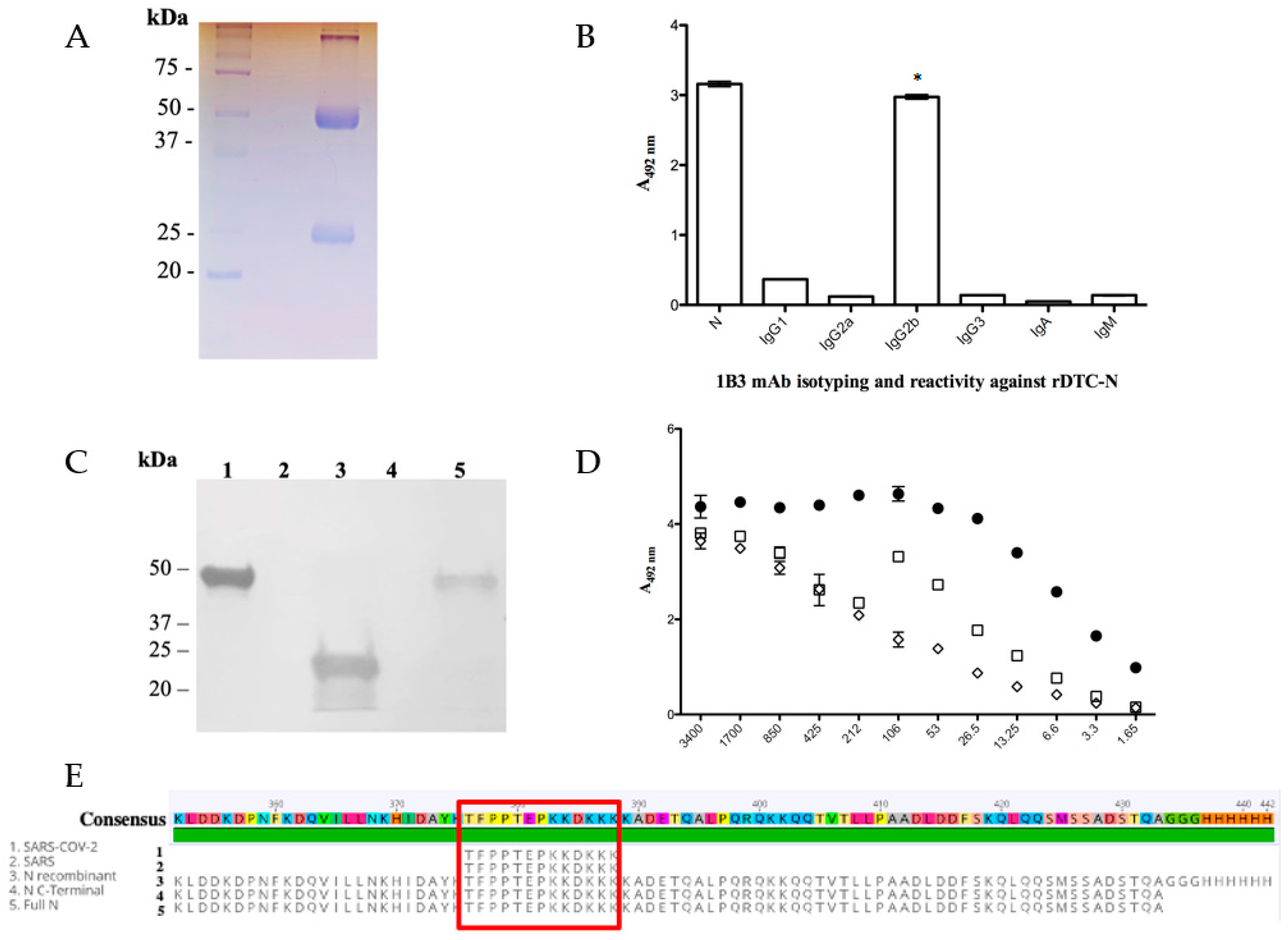
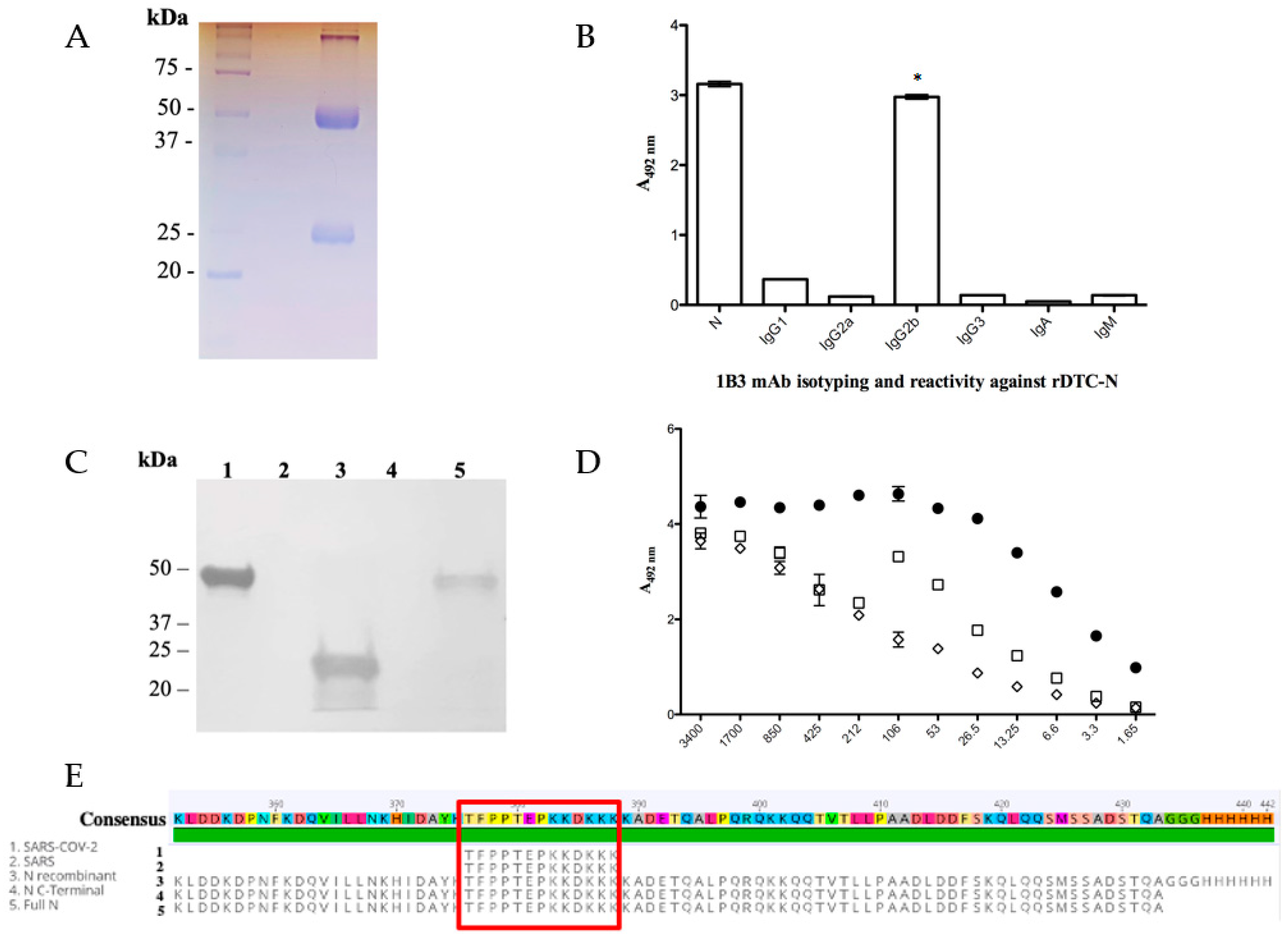
3.2. SARS-CoV-2 Nucleocapsid mAb (Anti-DTC-N 1B3 mAb) Functional Characterization
3.3. Ag-RDT Validation
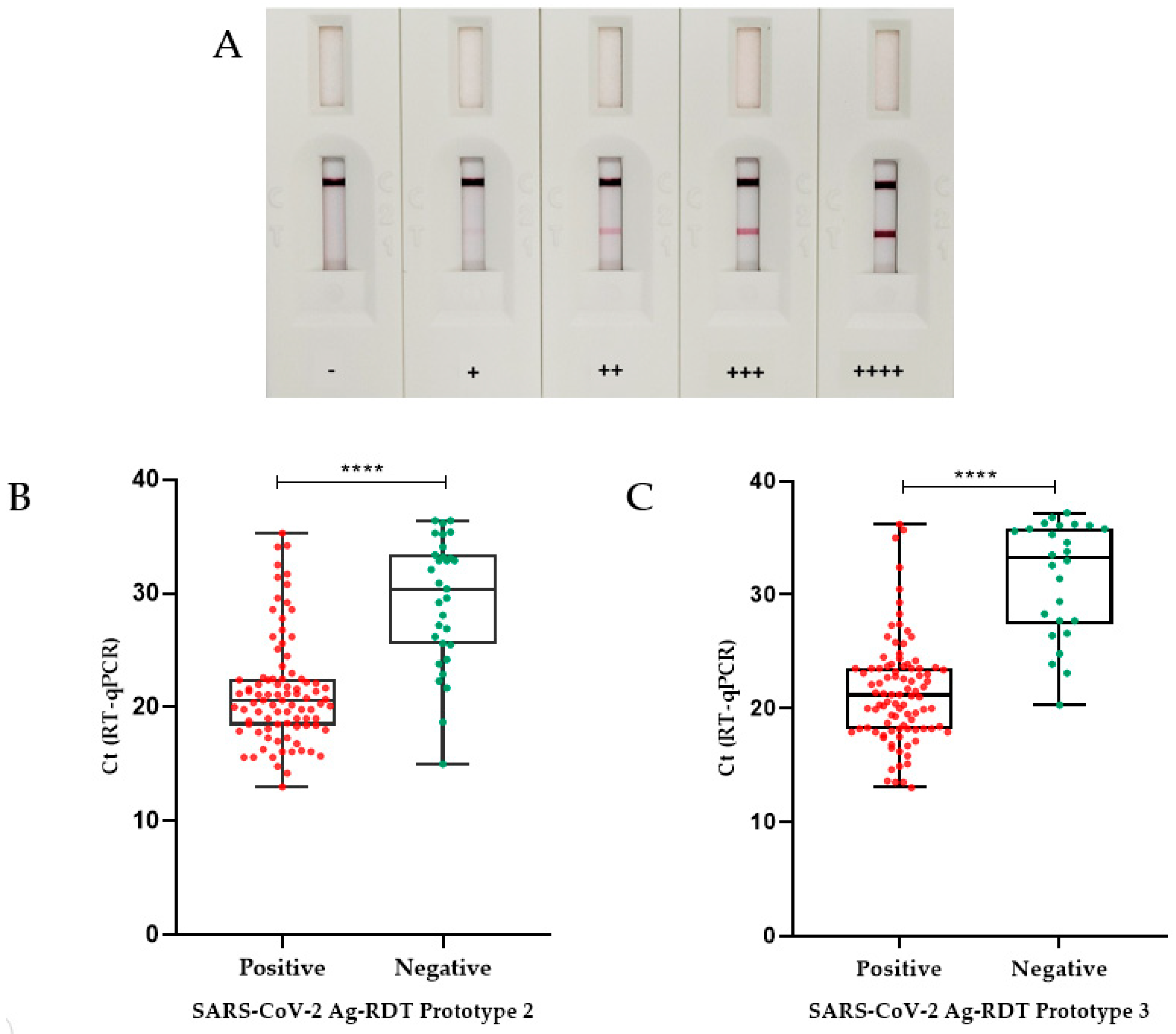
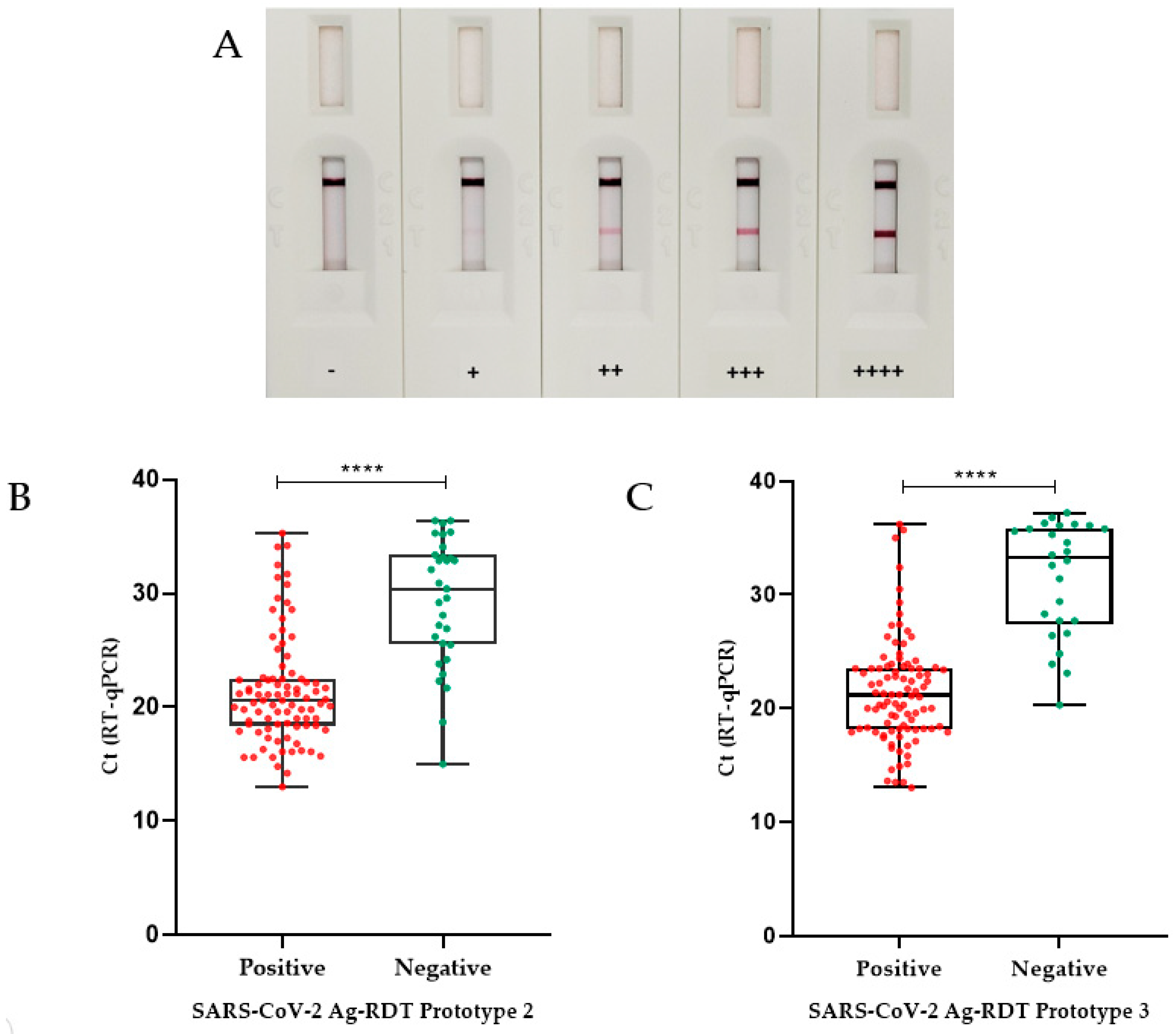
3.3.1. Prototype 1 Performance
3.3.2. Prototype 2 Performance
3.3.3. Prototype 3 Performance
3.4. Ag-RDT Follow-Up Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 4 September 2023).
- Rahman, S.; Rahman, M.M.; Miah, M.; Begum, M.N.; Sarmin, M.; Mahfuz, M.; Hossain, M.E.; Rahman, M.Z.; Chisti, M.J.; Ahmed, T. COVID-19 Reinfections among Naturally Infected and Vaccinated Individuals. Sci. Rep. 2022, 12, 1438. [Google Scholar] [CrossRef]
- Giovanetti, M.; Fonseca, V.; Wilkinson, E.; Tegally, H.; San, E.J.; Althaus, C.L.; Xavier, J.; Nanev Slavov, S.; Viala, V.L.; Ranieri Jerônimo Lima, A. Replacement of the Gamma by the Delta Variant in Brazil: Impact of Lineage Displacement on the Ongoing Pandemic. Virus Evol. 2022, 8, veac024. [Google Scholar] [CrossRef]
- Naveca, F.G.; Nascimento, V.; Souza, V.; Corado, A.L.; Nascimento, F.; Silva, G.; Mejía, M.C.; Brandão, M.J.; Costa, Á.; Duarte, D. Spread of Gamma (P. 1) Sub-Lineages Carrying Spike Mutations Close to the Furin Cleavage Site and Deletions in the N-Terminal Domain Drives Ongoing Transmission of SARS-CoV-2 in Amazonas, Brazil. medRxiv 2021, 10, e02366-21. [Google Scholar] [CrossRef]
- Baral, P.; Bhattarai, N.; Hossen, M.L.; Stebliankin, V.; Gerstman, B.S.; Narasimhan, G.; Chapagain, P.P. Mutation-Induced Changes in the Receptor-Binding Interface of the SARS-CoV-2 Delta Variant B. 1.617. 2 and Implications for Immune Evasion. Biochem. Biophys. Res. Commun. 2021, 574, 14–19. [Google Scholar] [CrossRef]
- Adamoski, D.; Baura, V.A.D.; Rodrigues, A.C.; Royer, C.A.; Aoki, M.N.; Tschá, M.K.; Bonatto, A.C.; Wassem, R.; Nogueira, M.B.; Raboni, S.M. SARS-CoV-2 Delta and Omicron Variants Surge in Curitiba, Southern Brazil, and Its Impact on Overall COVID-19 Lethality. Viruses 2022, 14, 809. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antigen-Detection in the Diagnosis of SARS-CoV-2 Infection: Interim Guidance, 6 October 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Ricks, S.; Kendall, E.A.; Dowdy, D.W.; Sacks, J.A.; Schumacher, S.G.; Arinaminpathy, N. Quantifying the Potential Value of Antigen-Detection Rapid Diagnostic Tests for COVID-19: A Modelling Analysis. BMC Med. 2021, 19, 75. [Google Scholar] [CrossRef]
- Peacock, W.F.; Soto-Ruiz, K.M.; House, S.L.; Cannon, C.M.; Headden, G.; Tiffany, B.; Motov, S.; Merchant-Borna, K.; Chang, A.M.; Pearson, C. Utility of COVID-19 Antigen Testing in the Emergency Department. J. Am. Coll. Emerg. Physicians Open 2022, 3, e12605. [Google Scholar] [CrossRef]
- Kim, J.; Sung, H.; Lee, H.; Kim, J.-S.; Shin, S.; Jeong, S.; Choi, M.; Lee, H.-J. Development Committee and Clinical Evidence Research Team in National Evidence-Based Healthcare Collaborating Agency. Clinical Performance of Rapid and Point-of-Care Antigen Tests for SARS-CoV-2 Variants of Concern: A Living Systematic Review and Meta-Analysis. Viruses 2022, 14, 1479. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Tsoi, H.; Chan, K.; Wong, B.H.; Che, X.; Tam, V.K.; Tam, S.C.; Cheng, V.C.; Hung, I.F. Relative Rates of Non-Pneumonic SARS Coronavirus Infection and SARS Coronavirus Pneumonia. Lancet 2004, 363, 841–845. [Google Scholar] [CrossRef]
- Weishampel, Z.A.; Young, J.; Fischl, M.; Fischer, R.J.; Donkor, I.O.; Riopelle, J.C.; Schulz, J.E.; Port, J.R.; Saturday, T.A.; van Doremalen, N. OraSure InteliSwabTM Rapid Antigen Test Performance with the SARS-CoV-2 Variants of Concern—Alpha, Beta, Gamma, Delta, and Omicron. Viruses 2022, 14, 543. [Google Scholar] [CrossRef]
- Prefeitura de Guaranésia. Available online: https://www.prefguaranesia.mg.gov.br/adm2017/ (accessed on 1 September 2022).
- Secretaria de Estado de Minas Gerais, S. Boletim Epidemiológico Coronavírus. Available online: https://coronavirus.saude.mg.gov.br/boletim (accessed on 1 September 2022).
- Carvalho, A.F.; Rocha, R.P.; Gonçalves, A.P.; Silva, T.; Sato, H.I.; Vuitika, L.; Bagno, F.F.; Sérgio, S.A.; Figueiredo, M.M.; Martins, R.B. The Use of Denaturing Solution as Collection and Transport Media to Improve SARS-CoV-2 RNA Detection and Reduce Infection of Laboratory Personnel. Braz. J. Microbiol. 2021, 52, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L. Detection of 2019 Novel Coronavirus (2019-NCoV) by Real-Time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef]
- Lu, X.; Wang, L.; Sakthivel, S.K.; Whitaker, B.; Murray, J.; Kamili, S.; Lynch, B.; Malapati, L.; Burke, S.A.; Harcourt, J. US CDC Real-Time Reverse Transcription PCR Panel for Detection of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1654. [Google Scholar] [CrossRef]
- Tyson, J.R.; James, P.; Stoddart, D.; Sparks, N.; Wickenhagen, A.; Hall, G.; Choi, J.H.; Lapointe, H.; Kamelian, K.; Smith, A.D. Improvements to the ARTIC Multiplex PCR Method for SARS-CoV-2 Genome Sequencing Using Nanopore. BioRxiv 2020. [Google Scholar] [CrossRef]
- Dorlass, E.G.; Lourenço, K.L.; Magalhaes, R.D.M.; Sato, H.; Fiorini, A.; Peixoto, R.; Coelho, H.P.; Telezynski, B.L.; Scagion, G.P.; Ometto, T. Survey of SARS-CoV-2 Genetic Diversity in Two Major Brazilian Cities Using a Fast and Affordable Sanger Sequencing Strategy. Genomics 2021, 113, 4109–4115. [Google Scholar] [CrossRef]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.D.M.; de Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; de Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 Isolation from the First Reported Patients in Brazil and Establishment of a Coordinated Task Network. Mem. Inst. Oswaldo Cruz 2020, 115, e200342. [Google Scholar] [CrossRef]
- Bagno, F.F.; Sérgio, S.A.R.; Figueiredo, M.M.; Godoi, L.C.; Andrade, L.A.F.; Salazar, N.C.; Soares, C.P.; Aguiar, A.; Almeida, F.J.; da Silva, E.D.; et al. Development and validation of an enzyme-linked immunoassay kit for diagnosis and surveillance of COVID-19. J. Clin. Virol. Plus 2022, 2, 100101. [Google Scholar] [CrossRef]
- Rocha, L.B.; Luz, D.E.; Moraes, C.T.; Caravelli, A.; Fernandes, I.; Guth, B.E.; Horton, D.S.; Piazza, R.M. Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities. Toxins 2012, 4, 729–747. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Alves, R.P.D.S.; Caetano, B.A.; Pereira, L.R.; Mitsunari, T.; Amorim, J.H.; Polatto, J.M.; Botosso, V.F.; Gallina, N.M.F.; Palacios, R. Epitope Sequences in Dengue Virus NS1 Protein Identified by Monoclonal Antibodies. Antibodies 2017, 6, 14. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous Cultures of Fused Cells Secreting Antibody of Predefined Specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Caldas, C.; Coelho, V.P.; Rigden, D.J.; Neschich, G.; Moro, A.M.; Brígido, M.M. Design and Synthesis of Germline-Based Hemi-Humanized Single-Chain Fv against the CD18 Surface Antigen. Protein Eng. 2000, 13, 353–360. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA Sequencing with Chain-Terminating Inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Frens, G. Controlled Nucleation for the Regulation of the Particle Size in Monodisperse Gold Suspensions. Nat. Phys. Sci. 1973, 241, 20–22. [Google Scholar] [CrossRef]
- Matejtschuk, P.; Phillips, P. Product Stability and Accelerated Degradation Studies. Med. Anim. Cell Cult. 2007, 503–522. [Google Scholar]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Begum, M.N.; Jubair, M.; Nahar, K.; Rahman, S.; Talha, M.; Sarker, M.S.; Uddin, A.N.; Khaled, S.; Uddin, M.S.; Li, Z. Factors Influencing the Performance of Rapid SARS-CoV-2 Antigen Tests under Field Condition. J. Clin. Lab. Anal. 2022, 36, e24203. [Google Scholar] [CrossRef]
- Rastawicki, W.; Gierczyński, R.; Juszczyk, G.; Mitura, K.; Henry, B.M. Evaluation of PCL Rapid Point of Care Antigen Test for Detection of SARS-CoV-2 in Nasopharyngeal Swabs. J. Med. Virol. 2021, 93, 1920–1922. [Google Scholar] [CrossRef]
- Pérez-García, F.; Romanyk, J.; Moya Gutiérrez, H.; Labrador Ballestero, A.; Pérez Ranz, I.; González Arroyo, J.; González Ventosa, V.; Pérez-Tanoira, R.; Domingo Cruz, C.; Cuadros-González, J. Comparative Evaluation of Panbio and SD Biosensor Antigen Rapid Diagnostic Tests for COVID-19 Diagnosis. J. Med. Virol. 2021, 93, 5650–5654. [Google Scholar] [CrossRef]
- Blairon, L.; Cupaiolo, R.; Thomas, I.; Piteüs, S.; Wilmet, A.; Beukinga, I.; Tré-Hardy, M. Efficacy Comparison of Three Rapid Antigen Tests for SARS-CoV-2 and How Viral Load Impact Their Performance. J. Med. Virol. 2021, 93, 5783–5788. [Google Scholar] [CrossRef]
- Sood, N.; Shetgiri, R.; Rodriguez, A.; Jimenez, D.; Treminino, S.; Daflos, A.; Simon, P. Evaluation of the Abbott BinaxNOW Rapid Antigen Test for SARS-CoV-2 Infection in Children: Implications for Screening in a School Setting. PLoS ONE 2021, 16, e0249710. [Google Scholar] [CrossRef]
- Choudhary, S.; Ishrat, A. Validation of Rapid SARS-COV-2 Antigen Detection Test as a Screening Tool for Detection of Covid-19 Infection at District Hospital in Northern India. Asian J. Med. Sci. 2022, 13, 7–10. [Google Scholar] [CrossRef]
- Nóra, M.; Déri, D.; Veres, D.S.; Kis, Z.; Barcsay, E.; Pályi, B. Evaluating the Field Performance of Multiple SARS-Cov-2 Antigen Rapid Tests Using Nasopharyngeal Swab Samples. PLoS ONE 2022, 17, e0262399. [Google Scholar] [CrossRef] [PubMed]
- Wegrzynska, K.; Walory, J.; Charkiewicz, R.; Lewandowska, M.A.; Wasko, I.; Kozinska, A.; Majewski, P.; Baraniak, A. Clinical Validation of GenBody COVID-19 Ag, Nasal and Nasopharyngeal Rapid Antigen Tests for Detection of SARS-CoV-2 in European Adult Population. Biomedicines 2023, 11, 493. [Google Scholar] [CrossRef]
- SARS-CoV-2 Variants of Concern as of 24 August 2023. Available online: https://www.ecdc.europa.eu/en/covid-19/variants-concern (accessed on 25 August 2023).
- Abavisani, M.; Rahimian, K.; Mahdavi, B.; Tokhanbigli, S.; Mollapour Siasakht, M.; Farhadi, A.; Kodori, M.; Mahmanzar, M.; Meshkat, Z. Mutations in SARS-CoV-2 Structural Proteins: A Global Analysis. Virol. J. 2022, 19, 220. [Google Scholar] [CrossRef]
- Liotti, F.M.; Menchinelli, G.; Lalle, E.; Palucci, I.; Marchetti, S.; Colavita, F.; La Sorda, M.; Sberna, G.; Bordi, L.; Sanguinetti, M. Performance of a Novel Diagnostic Assay for Rapid SARS-CoV-2 Antigen Detection in Nasopharynx Samples. Clin. Microbiol. Infect. 2021, 27, 487–488. [Google Scholar] [CrossRef]
- Diao, B.; Wen, K.; Zhang, J.; Chen, J.; Han, C.; Chen, Y.; Wang, S.; Deng, G.; Zhou, H.; Wu, Y. Accuracy of a Nucleocapsid Protein Antigen Rapid Test in the Diagnosis of SARS-CoV-2 Infection. Clin. Microbiol. Infect. 2021, 27, 289.e1–289.e4. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 Entry into Cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Deerain, J.; Druce, J.; Tran, T.; Batty, M.; Yoga, Y.; Fennell, M.; Dwyer, D.E.; Kok, J.; Williamson, D.A. Assessment of the Analytical Sensitivity of 10 Lateral Flow Devices against the SARS-CoV-2 Omicron Variant. J. Clin. Microbiol. 2022, 60, e02479-21. [Google Scholar] [CrossRef] [PubMed]
- Rohde, J.; Himmel, W.; Hofinger, C.; Lâm, T.-T.; Schrader, H.; Wallstabe, J.; Kurzai, O.; Gágyor, I. Diagnostic Accuracy and Feasibility of a Rapid SARS-CoV-2 Antigen Test in General Practice–a Prospective Multicenter Validation and Implementation Study. BMC Prim. Care 2022, 23, 149. [Google Scholar] [CrossRef]
- Batra, A.; Clark, J.R.; Kang, A.K.; Ali, S.; Patel, T.R.; Shlobin, N.A.; Hoffman, S.C.; Lim, P.H.; Orban, Z.S.; Visvabharathy, L. Persistent Viral RNA Shedding of SARS-CoV-2 Is Associated with Delirium Incidence and Six-Month Mortality in Hospitalized COVID-19 Patients. GeroScience 2022, 44, 1241–1254. [Google Scholar] [CrossRef]
- Badu, K.; Oyebola, K.; Zahouli, J.Z.; Fagbamigbe, A.F.; de Souza, D.K.; Dukhi, N.; Amankwaa, E.F.; Tolba, M.F.; Sylverken, A.A.; Mosi, L. SARS-CoV-2 Viral Shedding and Transmission Dynamics: Implications of WHO COVID-19 Discharge Guidelines. Front. Med. 2021, 8, 648660. [Google Scholar] [CrossRef] [PubMed]
- Binnicker, M.J. Can Testing Predict SARS-CoV-2 Infectivity? The Potential for Certain Methods to Be Surrogates for Replication-Competent Virus. J. Clin. Microbiol. 2021, 59, e00469-21. [Google Scholar] [CrossRef] [PubMed]
- Ronchini, C.; Gandini, S.; Pasqualato, S.; Mazzarella, L.; Facciotti, F.; Mapelli, M.; IEO Covid Team; Frige’, G.; Passerini, R.; Pase, L.; et al. Lower Probability and Shorter Duration of Infections after COVID-19 Vaccine Correlate with Anti-SARS-CoV-2 Circulating IgGs. PLoS ONE 2022, 17, e0263014. [Google Scholar] [CrossRef] [PubMed]
- Moderbacher, C.R.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Kalimuddin, S.; Tham, C.Y.; Qui, M.; de Alwis, R.; Sim, J.X.; Lim, J.M.; Tan, H.-C.; Syenina, A.; Zhang, S.L.; Le Bert, N. Early T Cell and Binding Antibody Responses Are Associated with COVID-19 RNA Vaccine Efficacy Onset. Med 2021, 2, 682–688.e4. [Google Scholar] [CrossRef]
- Mazzoni, A.; Vanni, A.; Spinicci, M.; Lamacchia, G.; Kiros, S.T.; Rocca, A.; Capone, M.; Di Lauria, N.; Salvati, L.; Carnasciali, A.; et al. SARS-CoV-2 Infection and Vaccination Trigger Long-Lived B and CD4+ T Lymphocytes with Implications for Booster Strategies. J. Clin. Investig. 2022, 132, e157990. [Google Scholar] [CrossRef]
- Azevedo, P.O.; Hojo-Souza, N.S.; Faustino, L.P.; Fumagalli, M.J.; Hirako, I.C.; Oliveira, E.R.; Figueiredo, M.M.; Carvalho, A.F.; Doro, D.; Benevides, L. Differential Requirement of Neutralizing Antibodies and T Cells on Protective Immunity to SARS-CoV-2 Variants of Concern. NPJ Vaccines 2023, 8, 15. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Prototype 2 | ||||||
|---|---|---|---|---|---|---|
| RT-qPCR Positive Samples | Ct | Ag-RDT Positive | Ag-RDT Negative | Sensitivity (95% CI) | Specificity (95% CI) | Accuracy (95% CI) |
| 79 | Ct ≤ 25 | 72 | 7 | 91.1% (82.6–96.4%) | 90% (79.5–96.2%) 54/60 | 90.6% (83.5–94.9%) |
| 18 | 25 < Ct ≤ 30 | 10 | 8 | 55.6% (30.8–78.5%) | 82.1% (71.7–89.8%) | |
| 23 | Ct > 30 | 7 | 16 | 30.4% (13.2–52.9%) | 71.2% (62.7–82.6%) | |
| 120 | Total | 89 | 31 | 74.2% (65.4–81.7%) | 79.4% (72.8–85.1%) | |
| Prototype 3 | ||||||
| RT-qPCR positive sample | Ct | Ag-RDT positive | Ag-RDT negative | Sensitivity (95% CI) | Specificity (95% CI) | Accuracy (95% CI) |
| 84 | Ct ≤ 25 | 80 | 4 | 95.2% (88.2–98.7%) | 98.3% (91.1–99.9%) 59/60 | 96.5% (92.1–98.8%) |
| 15 | 25 < Ct ≤ 30 | 9 | 6 | 60.0% (32.3–83.6%) | 90.7% (81.7–96.2%) | |
| 21 | Ct > 30 | 5 | 16 | 23.8% (8.2–47.2%) | 79.0% (68.5–87.3%) | |
| 120 | Total | 94 | 26 | 78.3% (69.9–85.3%) | 85.0% (78.9–89.9%) | |
| Days of Symptoms Onset | RT-qPCR Positive Samples | Mean Ct | Ag-RDT Positive Samples | Sensitivity (95% CI) |
|---|---|---|---|---|
| 0 to 3 | 25 | 21.5 | 25 | 100.0% (86.3–100.0%) |
| 4 to 7 | 45 | 25.2 | 29 | 64.4% (48.9–78.1%) |
| >7 | 42 | 30.8 | 18 | 42.8% (27.7–59.0%) |
| Author | Country | Samples (n) | Commercial Kit | Sensitivity | Specificity | ||
|---|---|---|---|---|---|---|---|
| + | − | Ct ≤ 25 | Overall | ||||
| Rastawicki, 2021 [31] | Poland | 95 | 46 | PCL COVID-19 Ag Rapid FIA (ROK) | 92.9% | 38.9% | 83.3% |
| Pérez-García, 2021 [32] | Spain | 186 | 170 | Panbio COVID-19 Ag Rapid Test Abbott (USA) | 98.9% | 60.0% | 100% |
| SD Biosensor Ag (ROK) | 97.4% | 66.5% | 97.3% | ||||
| Blairon, 2021 [33] | Belgium | 150 | 49 | Coronavirus Ag Rapid Test Cassette Bio-Rad (USA) | 97.1% | 60.0% | 100% |
| GSD NovaGen SARS-CoV-2 Antigen Rapid Test (China) | 95.7% | 59.3% | 85.7% | ||||
| Aegle Coronavirus Ag Rapid Test Cassette LumiraDx (UK) | 97.1% | 61.1% | 100% | ||||
| Sood, 2021 [34] | USA | 226 | 548 | BinaxNOW™ Abbott (USA) | 93.8% | 56.2% | 98.4% |
| Choudhary, 2022 [35] | India | 129 | 627 | SD Biosensor, Inc. (ROK) | NR | 55.0% | 99.2% |
| Nóra, 2022 [36] | Hungary | 40 | 58 | GenBody COVID-19 Ag (ROK) | 93.8% | 62.0% | 86.4% |
| Wegrzynska, 2023 [37] | Poland | 103 | 301 | GenBody COVID-19 Ag (ROK) | 100% | 97.1% | 100% |
| Prototypes | Brazil | 120 | 60 | SARS-CoV-2 Ag RDT Prototype 2 (Brazil) | 92.0% | 74.1% | 90.0% |
| 120 | 60 | SARS-CoV-2 Ag RDT Prototype 3 (Brazil) | 95.2% | 78.3% | 98.3% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, F.F.; da Silva, M.A.; Lopes, T.B.; Polatto, J.M.; de Castro, N.S.; Andrade, L.A.F.; Lourenço, K.L.; Sato, H.I.; de Carvalho, A.F.; Coelho, H.P.; et al. SARS-CoV-2 Rapid Antigen Test Based on a New Anti-Nucleocapsid Protein Monoclonal Antibody: Development and Real-Time Validation. Microorganisms 2023, 11, 2422. https://doi.org/10.3390/microorganisms11102422
Coelho FF, da Silva MA, Lopes TB, Polatto JM, de Castro NS, Andrade LAF, Lourenço KL, Sato HI, de Carvalho AF, Coelho HP, et al. SARS-CoV-2 Rapid Antigen Test Based on a New Anti-Nucleocapsid Protein Monoclonal Antibody: Development and Real-Time Validation. Microorganisms. 2023; 11(10):2422. https://doi.org/10.3390/microorganisms11102422
Chicago/Turabian StyleCoelho, Fabiana Fioravante, Miriam Aparecida da Silva, Thiciany Blener Lopes, Juliana Moutinho Polatto, Natália Salazar de Castro, Luis Adan Flores Andrade, Karine Lima Lourenço, Hugo Itaru Sato, Alex Fiorini de Carvalho, Helena Perez Coelho, and et al. 2023. "SARS-CoV-2 Rapid Antigen Test Based on a New Anti-Nucleocapsid Protein Monoclonal Antibody: Development and Real-Time Validation" Microorganisms 11, no. 10: 2422. https://doi.org/10.3390/microorganisms11102422
APA StyleCoelho, F. F., da Silva, M. A., Lopes, T. B., Polatto, J. M., de Castro, N. S., Andrade, L. A. F., Lourenço, K. L., Sato, H. I., de Carvalho, A. F., Coelho, H. P., Bagno, F. F., Luz, D., Viala, V. L., Cattony, P. Q., Melo, B. d. S., Moro, A. M., Quintilio, W., Barbosa, A. P., Bomfim, C. G., ... Fernandes, A. P. (2023). SARS-CoV-2 Rapid Antigen Test Based on a New Anti-Nucleocapsid Protein Monoclonal Antibody: Development and Real-Time Validation. Microorganisms, 11(10), 2422. https://doi.org/10.3390/microorganisms11102422