
Metagenomic Analysis Reveals Variations in Gut Microbiomes of the Schistosoma mansoni-Transmitting Snails Biomphalaria straminea and Biomphalaria glabrata

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Background
2. Methods
2.1. Samples Collection
2.2. DNA Extraction
2.3. Metagenomic Sequencing, Quality Control, Assembly and Annotation
2.4. Bacterial Data Analysis
2.5. Parasite Detection
2.6. Statistical Analysis
3. Results
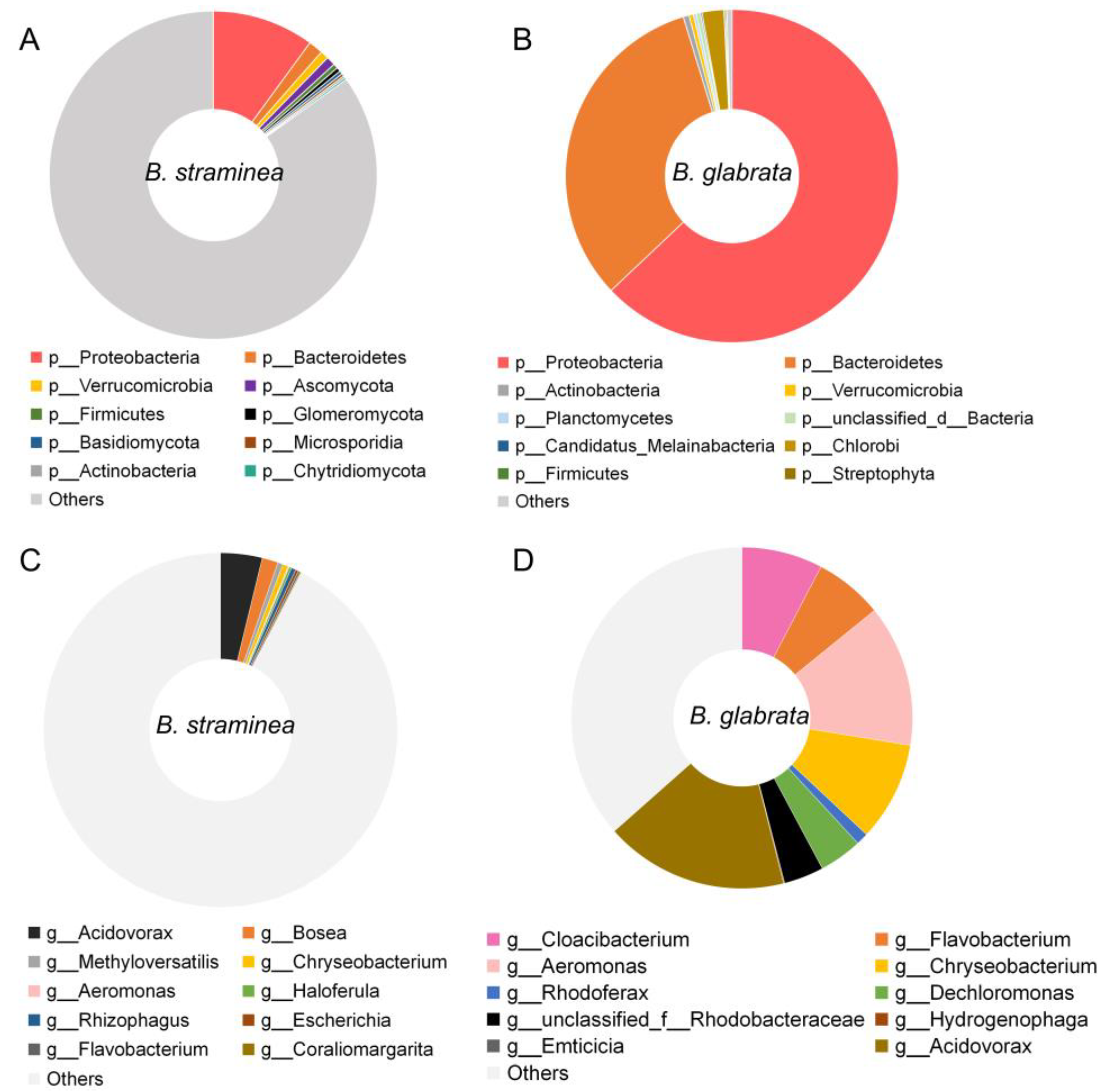
3.1. The Composition of Gut Microbes of B. straminea and B. glabrata
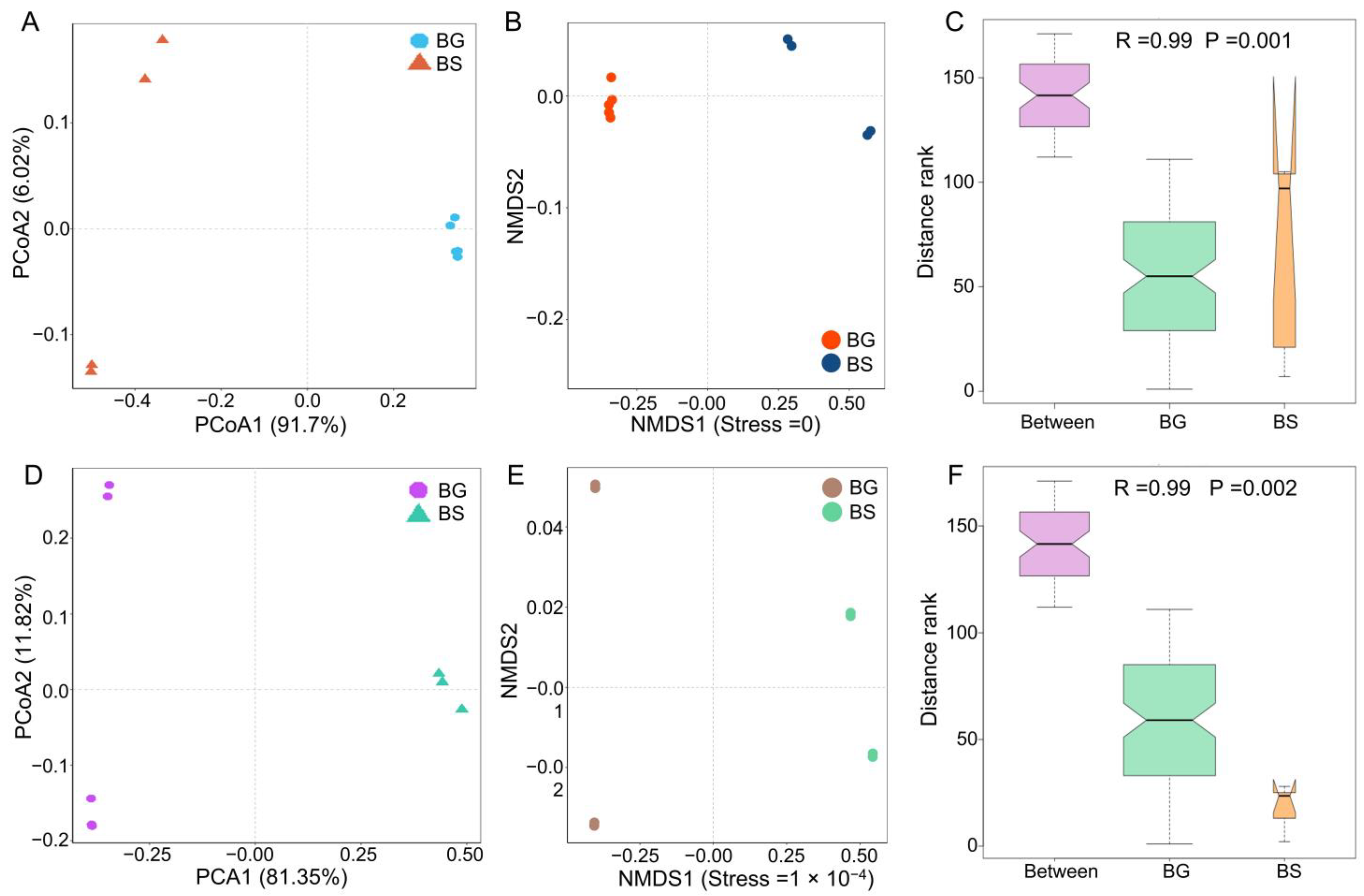
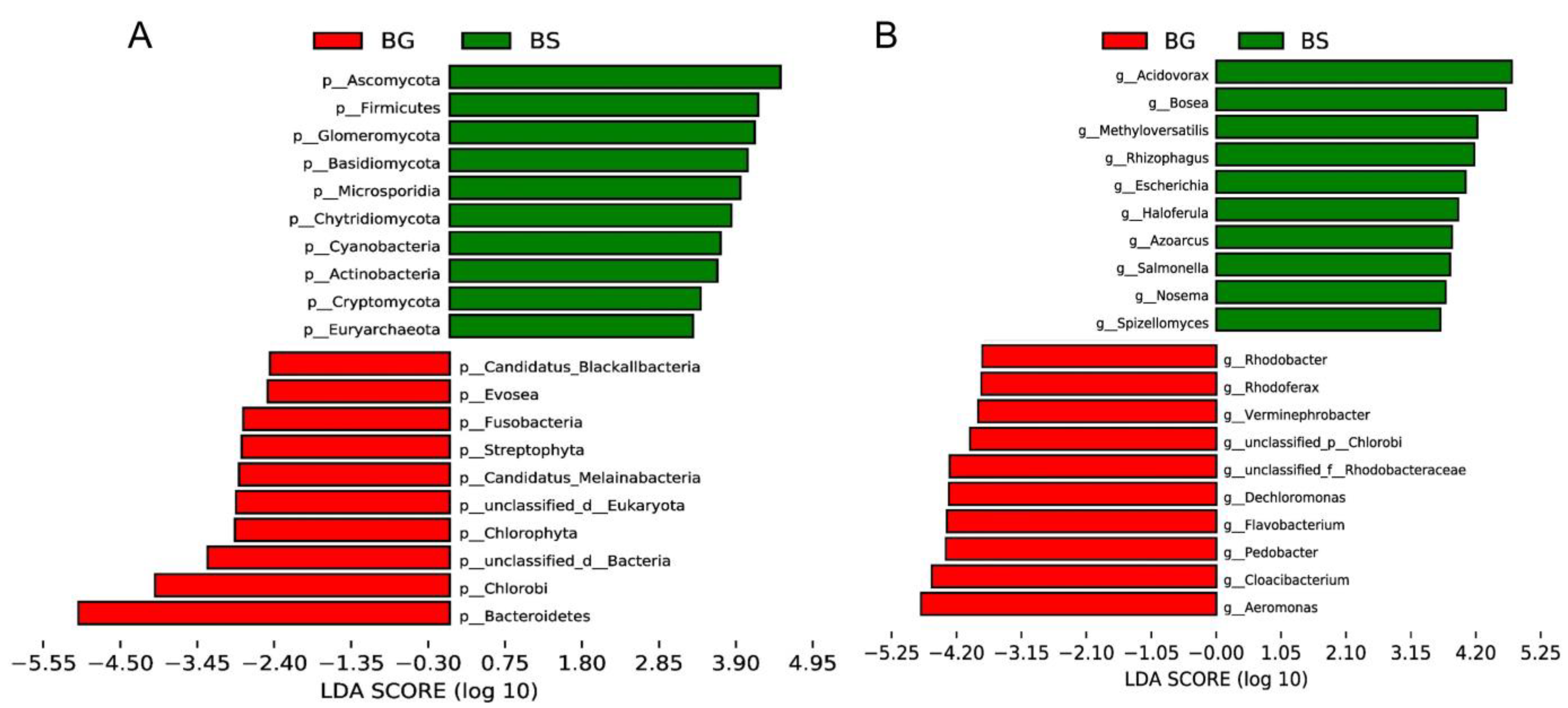
3.2. Microbiome Composition Shifts Associated with Host Species
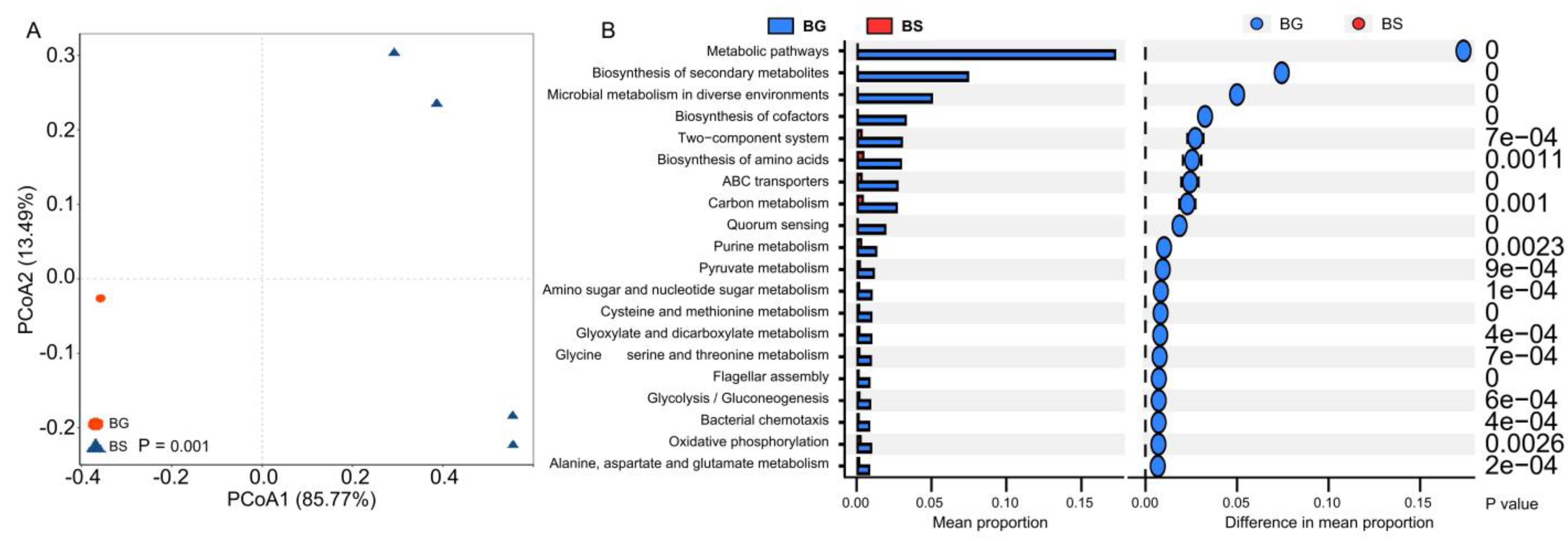
3.3. Bacterial Functional Shifts Associated with Host Species
3.4. The Distribution of ARGs of the Gut Microbiota of B. straminea and B. glabrata
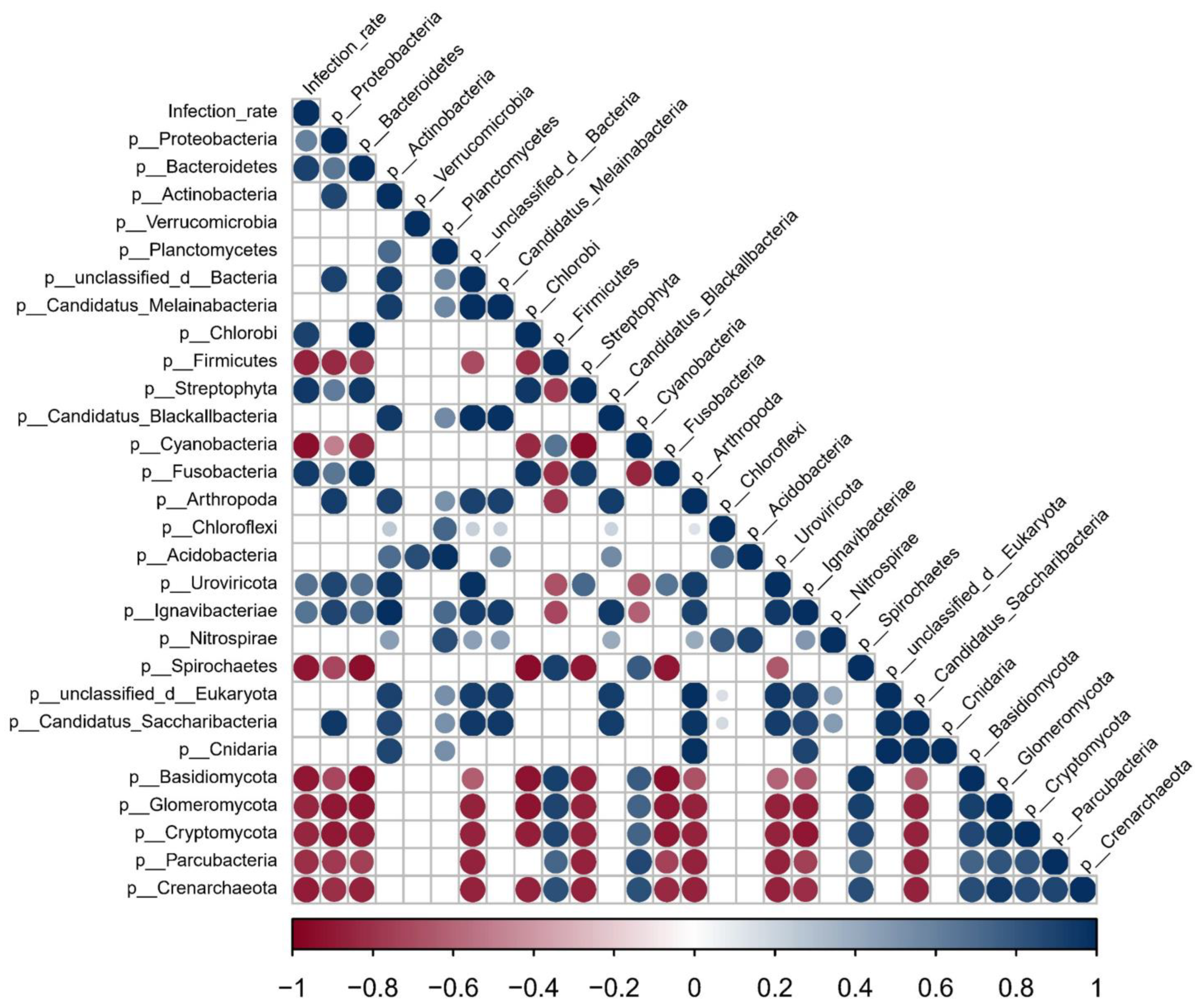
3.5. Potential Associations between Snail Gut Microbiota Features and Infection Rate of S. mansoni
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Barnett, R. Schistosomiasis. Lancet 2018, 392, 2431. [Google Scholar] [CrossRef] [PubMed]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef] [PubMed]
- Arican-Goktas, H.D.; Ittiprasert, W.; Bridger, J.M.; Knight, M. Differential spatial repositioning of activated genes in Biomphalaria glabrata snails infected with Schistosoma mansoni. PLoS Negl. Trop. Dis. 2014, 8, e3013. [Google Scholar] [CrossRef]
- Lin, D.; Xiang, S.; Sanogo, B.; Liang, Y.; Sun, X.; Wu, Z. Molecular Characterization of Rotifers and Their Potential Use in the Biological Control of Biomphalaria. Front. Cell Infect. Microbiol. 2021, 11, 744352. [Google Scholar] [CrossRef]
- Lin, D.; Zeng, X.; Sanogo, B.; He, P.; Xiang, S.; Du, S.; Zhang, Y.; Wang, L.; Wan, S.; Zeng, X.; et al. The potential risk of Schistosoma mansoni transmission by the invasive freshwater snail Biomphalaria straminea in South China. PLoS Negl. Trop. Dis. 2020, 14, e8310. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Xu, Y.; Zhou, H.; Huang, P.; Ma, Y.; Gao, M.; Cheng, S.; Zhou, H.; Lv, Z. Alterations of Gut Microbiome and Metabolite Profiling in Mice Infected by Schistosoma japonicum. Front. Immunol. 2020, 11, 569727. [Google Scholar] [CrossRef]
- Huang, P.; Zhou, M.; Cheng, S.; Hu, Y.; Gao, M.; Ma, Y.; Limpanont, Y.; Zhou, H.; Dekumyoy, P.; Cheng, Y.; et al. Myricetin Possesses Anthelmintic Activity and Attenuates Hepatic Fibrosis via Modulating TGFbeta1 and Akt Signaling and Shifting Th1/Th2 Balance in Schistosoma japonicum-Infected Mice. Front. Immunol. 2020, 11, 593. [Google Scholar] [CrossRef]
- Lin, D.; Song, Q.; Zhang, Y.; Liu, J.; Chen, F.; Du, S.; Xiang, S.; Wang, L.; Wu, X.; Sun, X. Bacillus subtilis Attenuates Hepatic and Intestinal Injuries and Modulates Gut Microbiota and Gene Expression Profiles in Mice Infected with Schistosoma japonicum. Front. Cell Dev. Biol. 2021, 9, 766205. [Google Scholar] [CrossRef]
- Lin, D.; Song, Q.; Liu, J.; Chen, F.; Zhang, Y.; Wu, Z.; Sun, X.; Wu, X. Potential Gut Microbiota Features for Non-Invasive Detection of Schistosomiasis. Front. Immunol. 2022, 13, 941530. [Google Scholar] [CrossRef]
- Madureira, A.C. Programmed Cell Death-Ligand-1 expression in Bladder Schistosomal Squamous Cell Carcinoma—There’s room for Immune Checkpoint Blockage? Front. Immunol. 2022, 13, 955000. [Google Scholar] [CrossRef]
- Trippler, L.; Ali, S.M.; Ame, S.M.; Hattendorf, J.; Suleiman, K.R.; Ali, M.N.; Juma, S.; Kabole, F.; Knopp, S. Fine-scale-mapping of Schistosoma haematobium infections at the school and community levels and intermediate host snail abundance in the north of Pemba Island: Baseline cross-sectional survey findings before the onset of a 3-year intervention study. Parasit. Vectors 2022, 15, 292. [Google Scholar] [CrossRef]
- Cesari, I.M.; Ballén, D.E.; Mendoza, L.; Ferrer, A.; Pointier, J.-P.; Kombila, M.; Richard-Lenoble, D.; Théron, A. Comparative evaluation of Schistosoma mansoni, Schistosoma intercalatum, and Schistosoma haematobium alkaline phosphatase antigenicity by the alkaline phosphatase immunoassay (APIA). Parasitol. Res. 2014, 113, 1395–1403. [Google Scholar] [CrossRef]
- Chienwichai, P.; Tipthara, P.; Tarning, J.; Limpanont, Y.; Chusongsang, P.; Chusongsang, Y.; Adisakwattana, P.; Reamtong, O. Metabolomics reveal alterations in arachidonic acid metabolism in Schistosoma mekongi after exposure to praziquantel. PLoS Negl. Trop. Dis. 2021, 15, e9706. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.O.; Sassa, M.; Parvin, N.; Islam, M.R.; Yajima, A.; Ota, E. Diagnostic test accuracy for detecting Schistosoma japonicum and S. mekongi in humans: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2021, 15, e9244. [Google Scholar] [CrossRef] [PubMed]
- Chienwichai, P.; Nogrado, K.; Tipthara, P.; Tarning, J.; Limpanont, Y.; Chusongsang, P.; Chusongsang, Y.; Tanasarnprasert, K.; Adisakwattana, P.; Reamtong, O. Untargeted serum metabolomic profiling for early detection of Schistosoma mekongi infection in mouse model. Front. Cell Infect. Microbiol. 2022, 12, 910177. [Google Scholar] [CrossRef] [PubMed]
- Brodin, P. Immune-microbe interactions early in life: A determinant of health and disease long term. Science 2022, 376, 945–950. [Google Scholar] [CrossRef]
- Du, S.; Sun, X.; Zhang, J.; Lin, D.; Chen, R.; Cui, Y.; Xiang, S.; Wu, Z.; Ding, T. Metagenome-Assembled Genomes Reveal Mechanisms of Carbohydrate and Nitrogen Metabolism of Schistosomiasis-Transmitting Vector Biomphalaria Glabrata. Microbiol. Spectr. 2022, 10, e184321. [Google Scholar] [CrossRef]
- Chevalier, C.; Stojanović, O.; Colin, D.J.; Suarez-Zamorano, N.; Tarallo, V.; Veyrat-Durebex, C.; Rigo, D.; Fabbiano, S.; Stevanović, A.; Hagemann, S.; et al. Gut Microbiota Orchestrates Energy Homeostasis during Cold. Cell 2015, 163, 1360–1374. [Google Scholar] [CrossRef]
- Videvall, E.; Song, S.J.; Bensch, H.M.; Strandh, M.; Engelbrecht, A.; Serfontein, N.; Hellgren, O.; Olivier, A.; Cloete, S.; Knight, R.; et al. Early-life gut dysbiosis linked to juvenile mortality in ostriches. Microbiome 2020, 8, 147. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Sukhchuluun, G.; Bo, T.B.; Chi, Q.S.; Yang, J.J.; Chen, B.; Zhang, L.; Wang, D.-H. Huddling remodels gut microbiota to reduce energy requirements in a small mammal species during cold exposure. Microbiome 2018, 6, 103. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, D.; Wang, L.; Hao, J.; Wang, J.; Zhou, X.; Wang, W.; Qiu, Q.; Huang, X.; Zhou, J.; et al. Convergent Evolution of Rumen Microbiomes in High-Altitude Mammals. Curr. Biol. 2016, 26, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Minich, J.J.; Härer, A.; Vechinski, J.; Frable, B.W.; Skelton, Z.R.; Kunselman, E.; Shane, M.A.; Perry, D.S.; Gonzalez, A.; McDonald, D.; et al. Host biology, ecology and the environment influence microbial biomass and diversity in 101 marine fish species. Nat. Commun. 2022, 13, 6978. [Google Scholar] [CrossRef] [PubMed]
- Baniel, A.; Amato, K.R.; Beehner, J.C.; Bergman, T.J.; Mercer, A.; Perlman, R.F.; Petrullo, L.; Reitsema, L.; Sams, S.; Lu, A.; et al. Seasonal shifts in the gut microbiome indicate plastic responses to diet in wild geladas. Microbiome 2021, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.S.; Shin, N.R.; Lee, J.B.; Kim, M.S.; Whon, T.W.; Hyun, D.W.; Yun, J.-H.; Jung, M.-J.; Kim, Y.J.; Bae, J.W. Host habitat is the major determinant of the gut microbiome of fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Luo, T.; Li, Y.; Zhang, W.; Liu, J.; Shi, H. Rumen and fecal microbiota profiles associated with immunity of young and adult goats. Front. Immunol. 2022, 13, 978402. [Google Scholar] [CrossRef]
- Ranasinghe, K.; Gunathilaka, N.; Amarasinghe, D.; Rodrigo, W.; Udayanga, L. Diversity of midgut bacteria in larvae and females of Aedes aegypti and Aedes albopictus from Gampaha District, Sri Lanka. Parasit. Vectors 2021, 14, 433. [Google Scholar] [CrossRef]
- Wan, S.; Sun, X.; Wu, F.; Yu, Z.; Wang, L.; Lin, D.; Li, Z.; Wu, Z.; Sun, X. Chi3l3: A potential key orchestrator of eosinophil recruitment in meningitis induced by Angiostrongylus cantonensis. J. Neuroinflammation. 2018, 15, 31. [Google Scholar] [CrossRef]
- Ji, L.; Yiyue, X.; Xujin, H.; Minghui, Z.; Mengying, Z.; Yue, H.; Yanqi, W.; Langui, S.; Xin, Z.; Datao, L.; et al. Study on the tolerance and adaptation of rats to Angiostrongylus cantonensis infection. Parasitol. Res. 2017, 116, 1937–1945. [Google Scholar] [CrossRef]
- Song, L.G.; Zheng, X.Y.; Lin, D.T.; Wang, G.X.; Wu, Z.D. Parasitology should not be abandoned: Data from outpatient parasitological testing in Guangdong, China. Infect. Dis. Poverty. 2017, 6, 119. [Google Scholar] [CrossRef]
- Pinheiro, G.L.; Correa, R.F.; Cunha, R.S.; Cardoso, A.M.; Chaia, C.; Clementino, M.M.; Garcia, E.S.; de Souza, W.; Frasés, S. Isolation of aerobic cultivable cellulolytic bacteria from different regions of the gastrointestinal tract of giant land snail Achatina fulica. Front. Microbiol. 2015, 6, 860. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-H.; Lv, S.; Lu, Y.; Bi, D.-Q.; Guo, Y.-H.; Wu, J.-T.; Yue, Z.-Y.; Mao, G.-Y.; Guo, Z.-X.; Zhang, Y.; et al. Spatial structure of the microbiome in the gut of Pomacea canaliculata. BMC Microbiol. 2019, 19, 273. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, S.; Xiao, Q.; Lin, Y.; Li, X.; Qu, Y.; Wu, G.; Li, H. Composition and diversity of gut microbiota in Pomacea canaliculata in sexes and between developmental stages. BMC Microbiol. 2021, 21, 200. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lin, L.; Xie, F.; Jin, W.; Zhu, W.; Wang, M.; Qiu, Q.; Li, Z.; Liu, J.; Mao, S. Metagenomic insights into the microbe-mediated B and K(2) vitamin biosynthesis in the gastrointestinal microbiome of ruminants. Microbiome 2022, 10, 109. [Google Scholar] [CrossRef]
- Salazar, C.; Giménez, M.; Riera, N.; Parada, A.; Puig, J.; Galiana, A.; Grill, F.; Vieytes, M.; Mason, C.E.; Antelo, V.; et al. Human microbiota drives hospital-associated antimicrobial resistance dissemination in the urban environment and mirrors patient case rates. Microbiome 2022, 10, 208. [Google Scholar] [CrossRef]
- Zhang, X.-L.; Deng, Y.-P.; Yang, T.; Li, L.-Y.; Cheng, T.-Y.; Liu, G.-H.; Duan, D.-Y. Metagenomics of the midgut microbiome of Rhipicephalus microplus from China. Parasit. Vectors 2022, 15, 48. [Google Scholar] [CrossRef]
- Isaac, S.; Flor-Duro, A.; Carruana, G.; Puchades-Carrasco, L.; Quirant, A.; Lopez-Nogueroles, M.; Pineda-Lucena, A.; Garcia-Garcera, M.; Ubeda, C. Microbiome-mediated fructose depletion restricts murine gut colonization by vancomycin-resistant Enterococcus. Nat. Commun. 2022, 13, 7718. [Google Scholar] [CrossRef]
- Qu, G.; Wang, W.; Lu, X.; Dai, J.; Li, X.; Liang, Y. Evaluating the risk of Schistosoma mansoni transmission in mainland China. Parasitol. Res. 2016, 115, 4711–4713. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Noguchi, H.; Park, J.; Takagi, T. MetaGene: Prokaryotic gene finding from environmental genome shotgun sequences. Nucleic Acids Res. 2006, 34, 5623–5630. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Avershina, E.; Frisli, T.; Rudi, K. De novo semi-alignment of 16S rRNA gene sequences for deep phylogenetic characterization of next generation sequencing data. Microbes Environ. 2013, 28, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.A.; Thiengo, S.C. Susceptibility of Biomphalaria straminea (Dunker, 1848) from Serra da Mesa Dam, Goias, Brazil to infection with three strains of Schistosoma mansoni Sambon, 1907. Mem. Inst. Oswaldo. Cruz. 2002, 97 (Suppl. 1), 59–60. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Ye, J.; Sun, Z.; He, Y.; Zhao, Y.; Ren, S.; Zhang, G.; Liu, M.; Zheng, P.; et al. Multi-omics reveal microbial determinants impacting the treatment outcome of antidepressants in major depressive disorder. Microbiome 2023, 11, 195. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Larsson, D.G. Antibiotic resistance genes in the environment: Prioritizing risks. Nat. Rev. Microbiol. 2015, 13, 396. [Google Scholar] [CrossRef]
- Young, C.C.W.; Karmacharya, D.; Bista, M.; Sharma, A.N.; Goldstein, T.; Mazet, J.A.K.; Johnson, C.K. Antibiotic resistance genes of public health importance in livestock and humans in an informal urban community in Nepal. Sci. Rep. 2022, 12, 13808. [Google Scholar] [CrossRef]
- Adema, C.M.; Hillier, L.W.; Jones, C.S.; Loker, E.S.; Knight, M.; Minx, P.; Oliveira, G.; Raghavan, N.; Shedlock, A.; do Amaral, L.R.; et al. Whole genome analysis of a schistosomiasis-transmitting freshwater snail. Nat. Commun. 2017, 8, 15451. [Google Scholar] [CrossRef] [PubMed]
- Osório, J.B.; Pereira, L.d.M.; Giongo, A.; Marconatto, L.; Potriquet, J.; Candido, R.R.F.; Mulvenna, J.; Jones, M.; Graeff-Teixeira, C.; Morassutti, A.L. Mollusk microbiota shift during Angiostrongylus cantonensis infection in the freshwater snail Biomphalaria glabrata and the terrestrial slug Phillocaulis soleiformis. Parasitol. Res. 2020, 119, 2495–2503. [Google Scholar] [CrossRef] [PubMed]
- Portet, A.; Toulza, E.; Lokmer, A.; Huot, C.; Duval, D.; Galinier, R.; Gourbal, B. Experimental Infection of the Biomphalaria glabrata Vector Snail by Schistosoma mansoni Parasites Drives Snail Microbiota Dysbiosis. Microorganisms 2021, 9, 1084. [Google Scholar] [CrossRef] [PubMed]
- Worthmann, A.; John, C.; Rühlemann, M.C.; Baguhl, M.; Heinsen, F.-A.; Schaltenberg, N.; Heine, M.; Schlein, C.; Evangelakos, I.; Mineo, C.; et al. Cold-induced conversion of cholesterol to bile acids in mice shapes the gut microbiome and promotes adaptive thermogenesis. Nat. Med. 2017, 23, 839–849. [Google Scholar] [CrossRef]
- Lin, D.; Zheng, X.; Sanogo, B.; Ding, T.; Sun, X.; Wu, Z. Bacterial composition of midgut and entire body of laboratory colonies of Aedes aegypti and Aedes albopictus from Southern China. Parasit. Vectors 2021, 14, 586. [Google Scholar] [CrossRef]
- Kokou, F.; Sasson, G.; Friedman, J.; Eyal, S.; Ovadia, O.; Harpaz, S.; Cnaani, A.; Mizrahi, I. Core gut microbial communities are maintained by beneficial interactions and strain variability in fish. Nat. Microbiol. 2019, 4, 2456–2465. [Google Scholar] [CrossRef]
- Stephens, W.Z.; Burns, A.R.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J.M. The composition of the zebrafish intestinal microbial community varies across development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef]
- Cicala, F.; Cisterna-Celiz, J.A.; Moore, J.D.; Rocha-Olivares, A. Structure, dynamics and predicted functional role of the gut microbiota of the blue (Haliotis fulgens) and yellow (H. corrugata) abalone from Baja California Sur, Mexico. PeerJ 2018, 6, e5830. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, C.; Li, G.; Yi, X. The influence of species identity and geographic locations on gut microbiota of small rodents. Front. Microbiol. 2022, 13, 983660. [Google Scholar] [CrossRef]
- Lyu, T.; Zhu, J.; Yang, X.; Yang, W.; Zheng, Z. Responses of Gut Microbial Community Composition and Function of the Freshwater Gastropod Bellamya aeruginosa to Cyanobacterial Bloom. Front. Microbiol. 2022, 13, 906278. [Google Scholar] [CrossRef]
- Hao, Y.; Guan, W.; Wu, H.; Li, L.; Abe, E.M.; Xue, J.; Qin, Z.; Wang, Q.; Lv, S.; Xu, J.; et al. Intestinal microbiome profiles in Oncomelania hupensis in mainland China. Acta Trop. 2020, 201, 105202. [Google Scholar] [CrossRef] [PubMed]
- de Melo, E.S.; Brayner, F.A.; Junior, N.; Franca, I.; Alves, L.C. Investigation of defense response and immune priming in Biomphalaria glabrata and Biomphalaria straminea, two species with different susceptibility to Schistosoma mansoni. Parasitol. Res. 2020, 119, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Nong, W.; Yu, Y.; Aase-Remedios, M.E.; Xie, Y.; So, W.L.; Li, Y.; Wong, C.F.; Baril, T.; Law, S.T.S.; Lai, S.Y.; et al. Genome of the ramshorn snail Biomphalaria straminea—An obligate intermediate host of schistosomiasis. Gigascience 2022, 11, giac012. [Google Scholar] [PubMed]
- Cui, E.; Zhou, Z.; Gao, F.; Chen, H.; Li, J. Roles of substrates in removing antibiotics and antibiotic resistance genes in constructed wetlands: A review. Sci. Total Environ. 2023, 859 Pt 1, 160257. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Mu, X.; Zhang, K.; Lang, H.; Su, Q.; Li, X.; Zhou, X.; Zhang, X.; Zheng, H. Geographical resistome profiling in the honeybee microbiome reveals resistance gene transfer conferred by mobilizable plasmids. Microbiome 2022, 10, 69. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Liu, F.; Cao, J.; Lv, N.; Zhu, B.; Zhang, G.; Gao, G.F. Integrated metagenomic and metatranscriptomic profiling reveals differentially expressed resistomes in human, chicken, and pig gut microbiomes. Environ. Int. 2020, 138, 105649. [Google Scholar] [CrossRef]
- Tan, R.; Jin, M.; Shao, Y.; Yin, J.; Li, H.; Chen, T.; Shi, D.; Zhou, S.; Li, J.; Yang, D. High-sugar, high-fat, and high-protein diets promote antibiotic resistance gene spreading in the mouse intestinal microbiota. Gut Microbes. 2022, 14, 2022442. [Google Scholar] [CrossRef]
- Zhang, M.; Hou, L.; Zhu, Y.; Zhang, C.; Li, W.; Lai, X.; Yang, J.; Li, S.; Shu, H. Composition and distribution of bacterial communities and antibiotic resistance genes in fish of four mariculture systems. Environ. Pollut. 2022, 311, 119934. [Google Scholar] [CrossRef]
- Dutra, H.L.; Rocha, M.N.; Dias, F.B.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia Blocks Currently Circulating Zika Virus Isolates in Brazilian Aedes aegypti Mosquitoes. Cell Host Microbe. 2016, 19, 771–774. [Google Scholar] [CrossRef]
- Wang, S.; Dos-Santos, A.L.A.; Huang, W.; Liu, K.C.; Oshaghi, M.A.; Wei, G.; Agre, P.; Jacobs-Lorena, M. Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science 2017, 357, 1399–1402. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef] [PubMed]
- Liberti, J.; Engel, P. The gut microbiota—Brain axis of insects. Curr. Opin. Insect Sci. 2020, 39, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, J.; Cryan, J.F. Microbiota-brain interactions: Moving toward mechanisms in model organisms. Neuron 2021, 109, 3930–3953. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Hong, J.; Wu, M.; Yuan, Z.; Li, D.; Wu, Z.; Sun, X.; Lin, D. Metagenomic Analysis Reveals Variations in Gut Microbiomes of the Schistosoma mansoni-Transmitting Snails Biomphalaria straminea and Biomphalaria glabrata. Microorganisms 2023, 11, 2419. https://doi.org/10.3390/microorganisms11102419
Li P, Hong J, Wu M, Yuan Z, Li D, Wu Z, Sun X, Lin D. Metagenomic Analysis Reveals Variations in Gut Microbiomes of the Schistosoma mansoni-Transmitting Snails Biomphalaria straminea and Biomphalaria glabrata. Microorganisms. 2023; 11(10):2419. https://doi.org/10.3390/microorganisms11102419
Chicago/Turabian StyleLi, Peipei, Jinni Hong, Mingrou Wu, Zhanhong Yuan, Dinghao Li, Zhongdao Wu, Xi Sun, and Datao Lin. 2023. "Metagenomic Analysis Reveals Variations in Gut Microbiomes of the Schistosoma mansoni-Transmitting Snails Biomphalaria straminea and Biomphalaria glabrata" Microorganisms 11, no. 10: 2419. https://doi.org/10.3390/microorganisms11102419
APA StyleLi, P., Hong, J., Wu, M., Yuan, Z., Li, D., Wu, Z., Sun, X., & Lin, D. (2023). Metagenomic Analysis Reveals Variations in Gut Microbiomes of the Schistosoma mansoni-Transmitting Snails Biomphalaria straminea and Biomphalaria glabrata. Microorganisms, 11(10), 2419. https://doi.org/10.3390/microorganisms11102419