

Effects of Cold-Surge-Induced Nearshore Seawater Icing on the Eukaryotic Microalgal Community in Aoshan Bay, Qingdao


Abstract
1. Introduction
2. Materials and Methods
2.1. Station and Sample Collection
2.2. Sample Processing and Analysis
2.3. DNA Extraction, Amplification and Sequencing
2.4. Bioinformatics and Statistical Analysis
3. Results
3.1. Environment Parameters
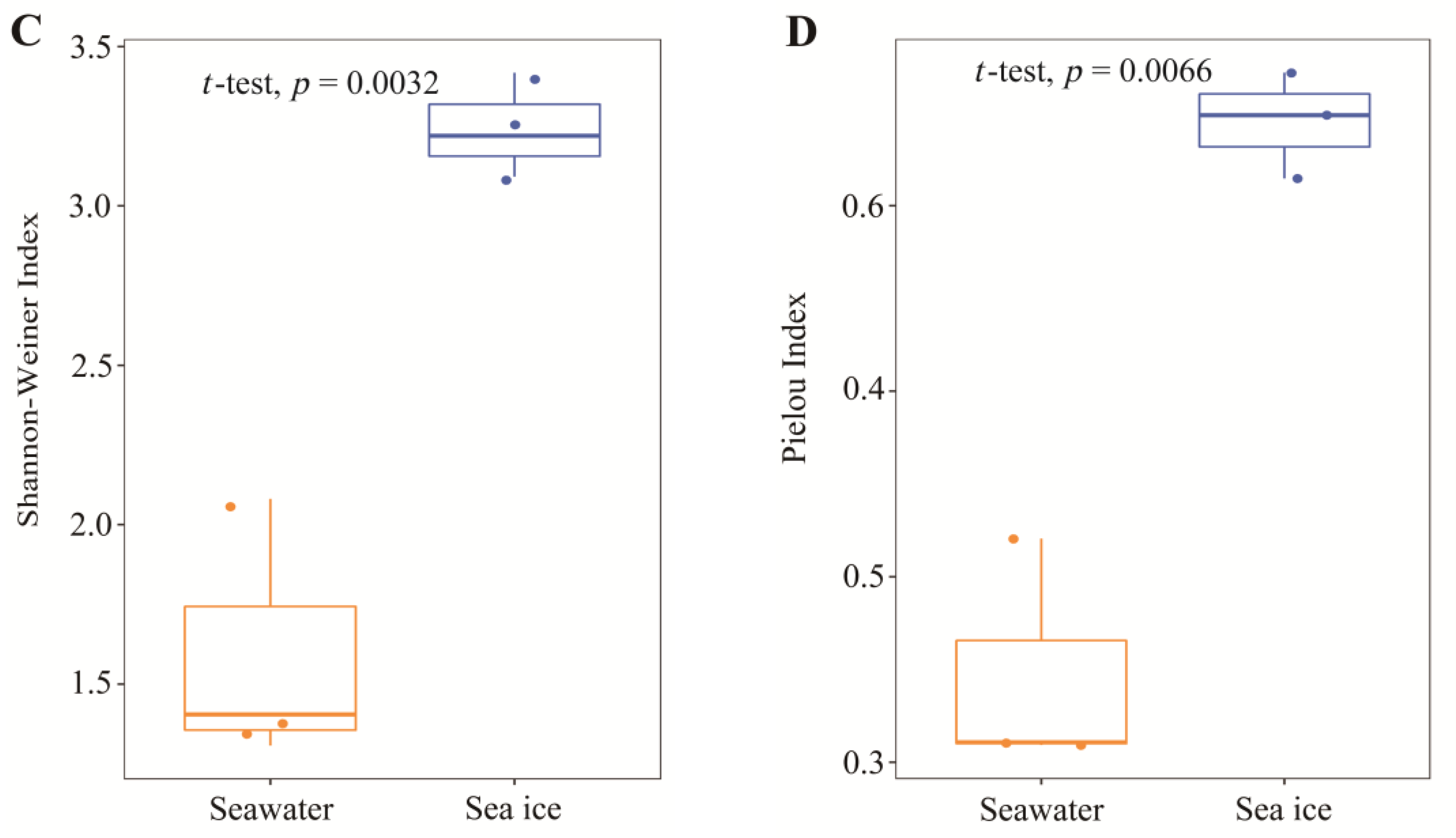
3.2. Eukaryotic Microalgal Diversity and Community Composition
3.3. Differences of Eukaryotic Microalgal Community in Seawater and Sea Ice
3.4. Correlation of Eukaryotic Microalgae Genera with Environmental Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gillett, N.P.; Stone, D.A.; Stott, P.A.; Nozawa, T.; Karpechko, A.Y.; Hegerl, G.C.; Wehner, M.F.; Jones, P.D. Attribution of polar warming to human influence. Nat. Geosci. 2008, 1, 750–754. [Google Scholar] [CrossRef]
- Najafi, M.R.; Zwiers, F.W.; Gillett, N.P. Attribution of Arctic temperature change to greenhouse-gas and aerosol influences. Nat. Clim. Chang. 2015, 5, 246–249. [Google Scholar] [CrossRef]
- Dai, G.K.; Li, C.X.; Han, Z.; Luo, D.H.; Yao, Y. The nature and predictability of the East Asian extreme cold events of 2020/21. Adv. Atmos. Sci. 2022, 39, 566–575. [Google Scholar] [CrossRef]
- Yao, Y.; Zhang, W.Q.; Luo, D.H.; Zhong, L.H.; Pei, L. Seasonal cumulative effect of Ural Blocking Episodes on the frequent cold events in China during the early winter of 2020/21. Adv. Atmos. Sci. 2022, 39, 609–624. [Google Scholar] [CrossRef]
- Wu, B.Y.; Su, J.Z.; Zhang, R.H. Effects of autumn-winter Arctic sea ice on winter Siberian High. Chin. Sci. Bull. 2011, 56, 3220–3228. [Google Scholar] [CrossRef]
- Mori, M.; Watanabe, M.; Shiogama, H.; Inoue, J.; Kimoto, M. Robust Arctic sea-ice influence on the frequent Eurasian cold winters in past decades. Nat. Geosci. 2014, 7, 869–873. [Google Scholar] [CrossRef]
- Liu, J.P.; Curry, J.A.; Wang, H.J.; Song, M.R.; Horton, R.M. Impact of declining Arctic sea ice on winter snowfall. Proc. Natl. Acad. Sci. USA 2012, 109, 4074–4079. [Google Scholar] [CrossRef]
- Kurczyn, J.A.; Appendini, C.M.; Beier, E.; Sosa-Lopez, A.; Lopez-Gonzalez, J.; Posada-Vanegas, G. Oceanic and atmospheric impact of central American cold surges (Nortes) in the Gulf of Mexico. Int. J. Climatol. 2021, 41, E1450–E1468. [Google Scholar] [CrossRef]
- Xie, X.; Zhou, R.; Deng, Z. Overcompensation and competitive effects of Microcystis aeruginosa and Scenedesmus obliquus after low temperature and light stresses. Acta Ecol. Sin. 2014, 34, 1224–1234. [Google Scholar]
- Grossmann, S.; Gleitz, M. Microbial response to experimental sea-ice formation: Implications for the establishment of Antarctic sea-ice communities. J. Exp. Mar. Biol. Ecol. 1993, 173, 273–289. [Google Scholar] [CrossRef]
- Worster, M.G.; Jones, D.W.R. Sea-ice thermodynamics and brine drainage. Philos. Trans. R. Soc. A-Math. Phys. Eng. Sci. 2015, 373, 20140166. [Google Scholar] [CrossRef]
- Niedrauer, T.M.; Martin, S. An experimental study of brine drainage and convection in young sea ice. J. Geophys. Res.-Ocean. 1979, 84, 1176–1186. [Google Scholar] [CrossRef]
- Campbell, K.; Mundy, C.J.; Belzile, C.; Delaforge, A.; Rysgaard, S. Seasonal dynamics of algal and bacterial communities in Arctic sea ice under variable snow cover. Polar Biol. 2018, 41, 41–58. [Google Scholar] [CrossRef]
- Lund-Hansen, L.; Sgaard, D.; Sorrell, B.; Gradinger, R.; Meiners, K. Arctic Sea Ice Ecology: Seasonal Dynamics in Algal and Bacterial Productivity; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Piontek, J.; Galgani, L.; Nothig, E.M.; Peeken, I.; Engel, A. Organic matter composition and heterotrophic bacterial activity at declining summer sea ice in the central Arctic Ocean. Limnol. Oceanogr. 2021, 66, S343–S362. [Google Scholar] [CrossRef]
- Delille, B.; Vancoppenolle, M.; Geilfus, N.X.; Tilbrook, B.; Lannuzel, D.; Schoemann, V.; Becquevort, S.; Carnat, G.; Delille, D.; Lancelot, C.; et al. Southern Ocean CO2 sink: The contribution of the sea ice. J. Geophys. Res.-Ocean. 2014, 119, 6340–6355. [Google Scholar] [CrossRef]
- Ligowski, R.; Jordan, R.W.; Assmy, P. Morphological adaptation of a planktonic diatom to growth in Antarctic sea ice. Mar. Biol. 2012, 159, 817–827. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kovaleva, G.V.; Povazhnyy, V.V.; Zolotareva, A.E.; Makarevich, P.R.; Matishov, D.G. Temporary ice microalgae community in Taganrog Bay of the Azov Sea. Oceanology 2014, 54, 618–623. [Google Scholar] [CrossRef]
- Freyria, N.J.; Kuo, A.; Chovatia, M.; Johnson, J.; Lipzen, A.; Barry, K.W.; Grigoriev, I.V.; Lovejoy, C. Salinity tolerance mechanisms of an Arctic Pelagophyte using comparative transcriptomic and gene expression analysis. Commun. Biol. 2022, 5, 13. [Google Scholar] [CrossRef]
- Molazadeh, M.; Ahmadzadeh, H.; Pourianfar, H.R.; Lyon, S.; Rampelotto, P.H. The use of microalgae for coupling wastewater treatment with CO2 biofixation. Front. Bioeng. Biotechnol. 2019, 7, 42. [Google Scholar] [CrossRef]
- Singh, U.B.; Ahluwalia, A.S. Microalgae: A promising tool for carbon sequestration. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 73–95. [Google Scholar] [CrossRef]
- Guo, Y.L.; Xiong, X.J.; Chen, C.; Yu, L.; Guo, Y.Q. Hydrometeorological interpretation and cause analysis on Qingdao’s warmer winter. Coast. Eng. 2014, 33, 1–11. (In Chinese) [Google Scholar]
- Hu, Y.B.; Wang, F.Y.; Boone, W.; Barber, D.; Rysgaard, S. Assessment and improvement of the sea ice processing for dissolved inorganic carbon analysis. Limnol. Oceanogr.-Methods 2018, 16, 83–91. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View. Software. 2021. Available online: https://odv.awi.de (accessed on 15 September 2022).
- Pierrot, D.; Lewis, E.; Wallace, R.; Wallace, D.; Wallace, W.; Wallace, D.W.R. MS Excel Program Developed for CO2 System Calculations; ORNL/CDIAC-105a; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy: Oak Ridge, TN, USA, 2006. [Google Scholar] [CrossRef]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicz, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep-Sea Res. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- Dickson, A.G. Standard potential of the reaction: AgCl (s) + 1/2H2 (g) = Ag (s)+HCl (aq), and the standard acidity constant of the ion HSO4− in synthetic sea water from 273.15 to 318.15 K. J. Chem. Thermodyn. 1990, 22, 113–127. [Google Scholar] [CrossRef]
- Uppström, L.R. The Boron/chlorinity ratio of deep-sea water from Pacific Ocean. Deep-Sea Res. 1974, 21, 161–162. [Google Scholar] [CrossRef]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; Wiley-VCH: Weinheim, Germany, 1999; 600p. [Google Scholar]
- Cox, G.F.N.; Weeks, W.F. Equations for determining the gas and brine volumes in sea-ice samples. J. Glaciol. 1983, 29, 306–316. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, P.; Xu, L.; Qu, L.Y.; Han, Y.J.; Zheng, L.W.; Gong, X.Z. Disproportionate responses between free-living and particle-attached bacteria during the transition to oxygen-deficient zones in the Bohai Seawater. Sci. Total Environ. 2021, 791, 148097. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Robeson, M.S.; O’Rourke, D.R.; Kaehler, B.D.; Ziemski, M.; Dillon, M.R.; Foster, J.T.; Bokulich, N.A. RESCRIPt: Reproducible sequence taxonomy reference database management for the masses. bioRxiv 2020. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Hu, K. Become competent in generating RNA-Seq Heat Maps in one day for novices without prior R experience. In Nuclear Reprogramming: Methods and Protocols; Hu, K., Ed.; Methods in Molecular Biology; Humana: New York, NY, USA, 2021; Volume 2239, pp. 269–303. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Villanueva, R.A.M.; Chen, Z.J. ggplot2: Elegant graphics for data analysis, 2nd edition. Meas.-Interdiscip. Res. Perspect. 2019, 17, 160–167. [Google Scholar] [CrossRef]
- Yadav, M.L.; Roychoudhury, B. Handling missing values: A study of popular imputation packages in R. Knowl.-Based Syst. 2018, 160, 104–118. [Google Scholar] [CrossRef]
- Ren, H.W.; Liu, J.H.; Li, G.; Hu, Y.B. Bacterial community in cold surge-caused sea ice differs from seawater in mid-latitude region: A case study in Aoshan Bay, Southern Yellow Sea. Front. Mar. Sci. 2022, 9, 856110. [Google Scholar] [CrossRef]
- Rozanska, M.; Poulin, M.; Gosselin, M. Protist entrapment in newly formed sea ice in the Coastal Arctic Ocean. J. Mar. Syst. 2008, 74, 887–901. [Google Scholar] [CrossRef]
- De Villiërs Louw, S.; Walker, D.R.; Fawcett, S.E. Factors influencing sea-ice algae abundance, community composition, and distribution in the marginal ice zone of the Southern Ocean during winter. Deep-Sea Res. Part I-Oceanogr. Res. Pap. 2022, 185, 103805. [Google Scholar] [CrossRef]
- Moro, I.; Paccagnella, R.; Barbante, C.; Andreoli, C. Microalgal communities of the sea ice, ice-covered and ice-free waters of Wood Bay (Ross Sea, Antarctica) during the austral summer 1993-94. Mar. Ecol.-Pubbl. Della Stn. Zool. Napoli I 2000, 21, 233–245. [Google Scholar] [CrossRef]
- Joli, N.; Monier, A.; Logares, R.; Lovejoy, C. Seasonal patterns in Arctic prasinophytes and inferred ecology of Bathycoccus unveiled in an Arctic winter metagenome. ISME J. 2017, 11, 1372–1385. [Google Scholar] [CrossRef]
- Trombetta, T.; Bouget, F.Y.; Felix, C.; Mostajir, B.; Vidussi, F. Microbial diversity in a North Western Mediterranean Sea Shallow Coastal Lagoon under contrasting Water temperature conditions. Front. Mar. Sci. 2022, 9, 858744. [Google Scholar] [CrossRef]
- Vaulot, D.; Lepere, C.; Toulza, E.; De la Iglesia, R.; Poulain, J.; Gaboyer, F.; Moreau, H.; Vandepoele, K.; Ulloa, O.; Gavory, F.; et al. Metagenomes of the Picoalga Bathycoccus from the Chile Coastal Upwelling. PLoS ONE 2012, 7, e39648. [Google Scholar] [CrossRef]
- Monier, A.; Sudek, S.; Fast, N.M.; Worden, A.Z. Gene invasion in distant eukaryotic lineages: Discovery of mutually exclusive genetic elements reveals marine biodiversity. ISME J. 2013, 7, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ruiz, M.; Prieto, A.; Barber-Lluch, E.; Teira, E. Amino acid utilization by eukaryotic picophytoplankton in a coastal upwelling system. Mar. Ecol. Prog. Ser. 2018, 588, 43–57. [Google Scholar] [CrossRef]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Goryunov, D.V.; Troitsky, A.V. Photosynthetic Picoeukaryotes in the Land-Fast Ice of the White Sea, Russia. Microb. Ecol. 2018, 75, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.; Schulz, K.G.; Riebesell, U.; Bellerby, R.; Delille, B.; Schartau, M. Effects of CO2 on particle size distribution and phytoplankton abundance during a mesocosm bloom experiment (PeECE II). Biogeosciences 2008, 5, 509–521. [Google Scholar] [CrossRef]
- Meakin, N.G.; Wyman, M. Rapid shifts in picoeukaryote community structure in response to ocean acidification. ISME J. 2011, 5, 1397–1405. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | S | T (°C) | pH | pCO2 | NO2− | NO3− | NH4+ | SiO32− | PO43− |
|---|---|---|---|---|---|---|---|---|---|
| S1sw | 40.4 | −1.0 | 8.16 | 346 | 0.5 | 3.8 | 2.8 | 5.1 | 0.1 |
| S2sw | 33.1 | −1.2 | 8.20 | 277 | 0.2 | 16.4 | 1.4 | 5.8 | 0.1 |
| S3sw | 32.2 | −1.0 | 8.24 | 239 | 0.2 | 12.1 | 0.6 | 4.8 | 0.0 |
| S1ice | 39.1 | −2.2 | 8.26 | 279 | 0.8 | 4.7 | 16.7 | 5.7 | 0.2 |
| S2ice | 55.8 | −3.2 | 8.29 | 374 | 2.9 | 15.0 | 33.2 | 20.4 | 0.9 |
| S3ice | 35.6 | −2.0 | 8.28 | 252 | 1.2 | - | 6.5 | 11.8 | 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, H.; Guo, X.; Xu, Y.; Hu, Y. Effects of Cold-Surge-Induced Nearshore Seawater Icing on the Eukaryotic Microalgal Community in Aoshan Bay, Qingdao. Microorganisms 2023, 11, 108. https://doi.org/10.3390/microorganisms11010108
Bian H, Guo X, Xu Y, Hu Y. Effects of Cold-Surge-Induced Nearshore Seawater Icing on the Eukaryotic Microalgal Community in Aoshan Bay, Qingdao. Microorganisms. 2023; 11(1):108. https://doi.org/10.3390/microorganisms11010108
Chicago/Turabian StyleBian, Haizhen, Xin Guo, Yanqiang Xu, and Yubin Hu. 2023. "Effects of Cold-Surge-Induced Nearshore Seawater Icing on the Eukaryotic Microalgal Community in Aoshan Bay, Qingdao" Microorganisms 11, no. 1: 108. https://doi.org/10.3390/microorganisms11010108
APA StyleBian, H., Guo, X., Xu, Y., & Hu, Y. (2023). Effects of Cold-Surge-Induced Nearshore Seawater Icing on the Eukaryotic Microalgal Community in Aoshan Bay, Qingdao. Microorganisms, 11(1), 108. https://doi.org/10.3390/microorganisms11010108