
The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
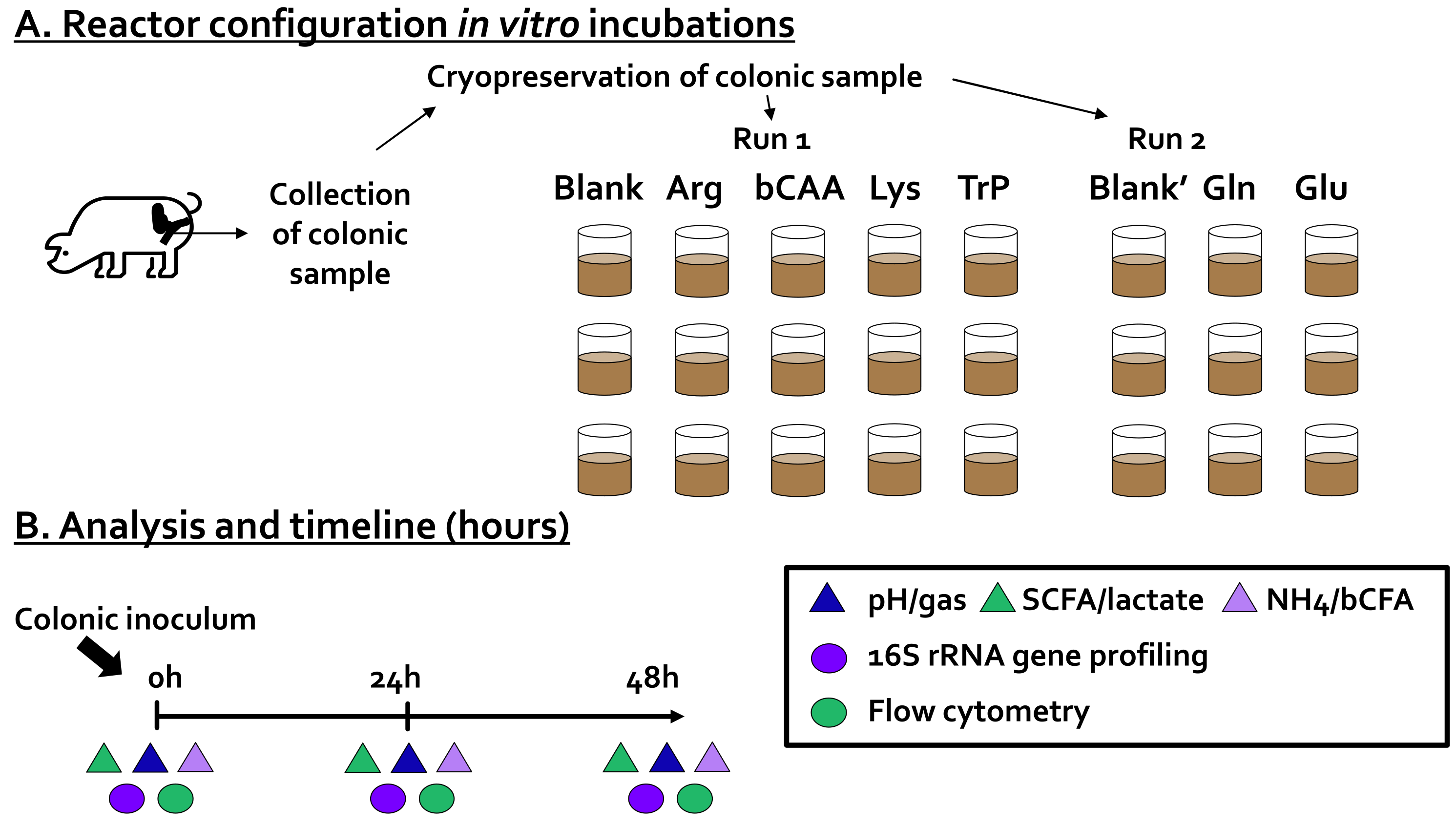
2.2. Incubation Strategy
2.3. Microbial Activity Analysis
2.4. Microbial Composition Analysis
2.5. Statistics
3. Results
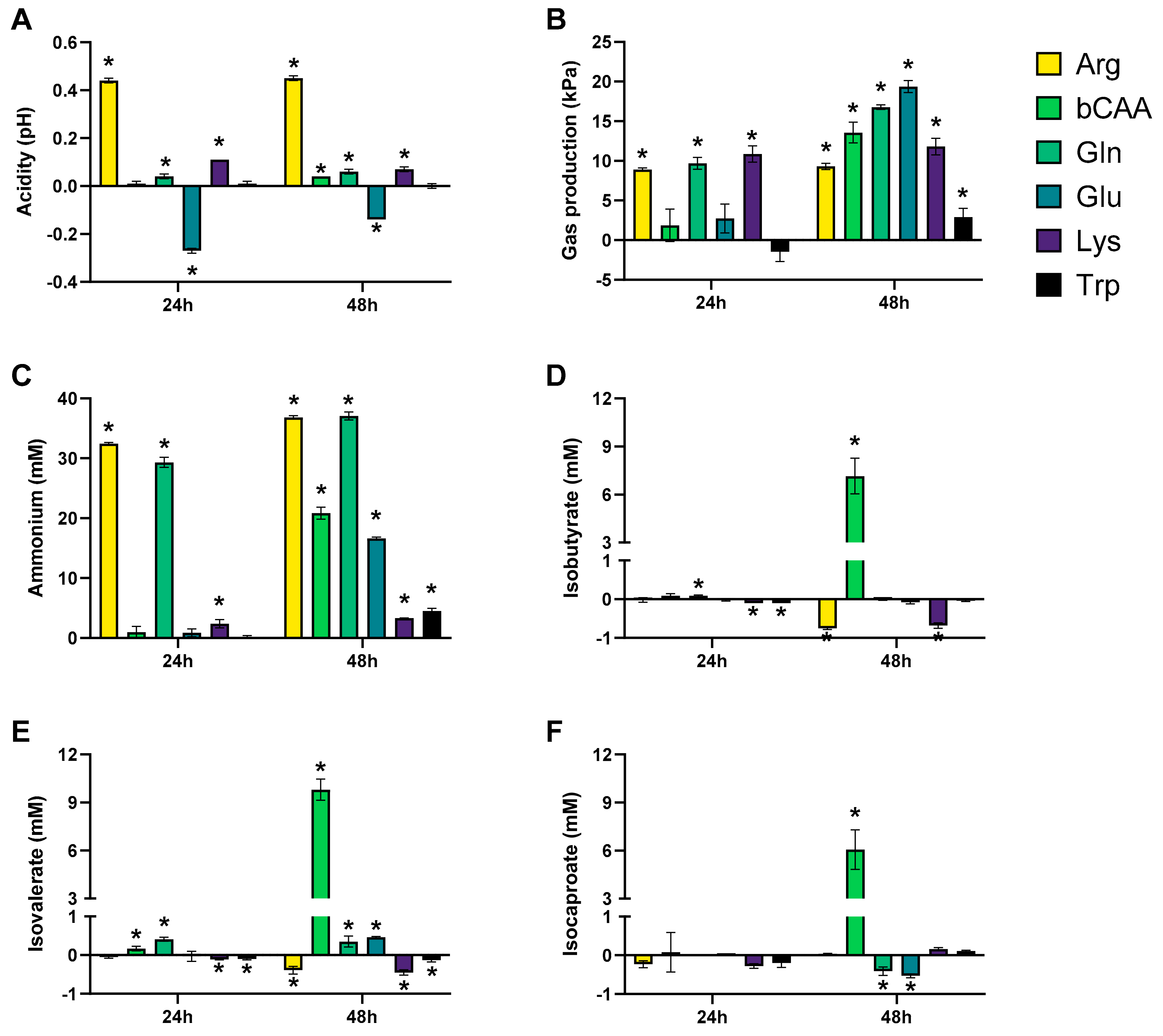
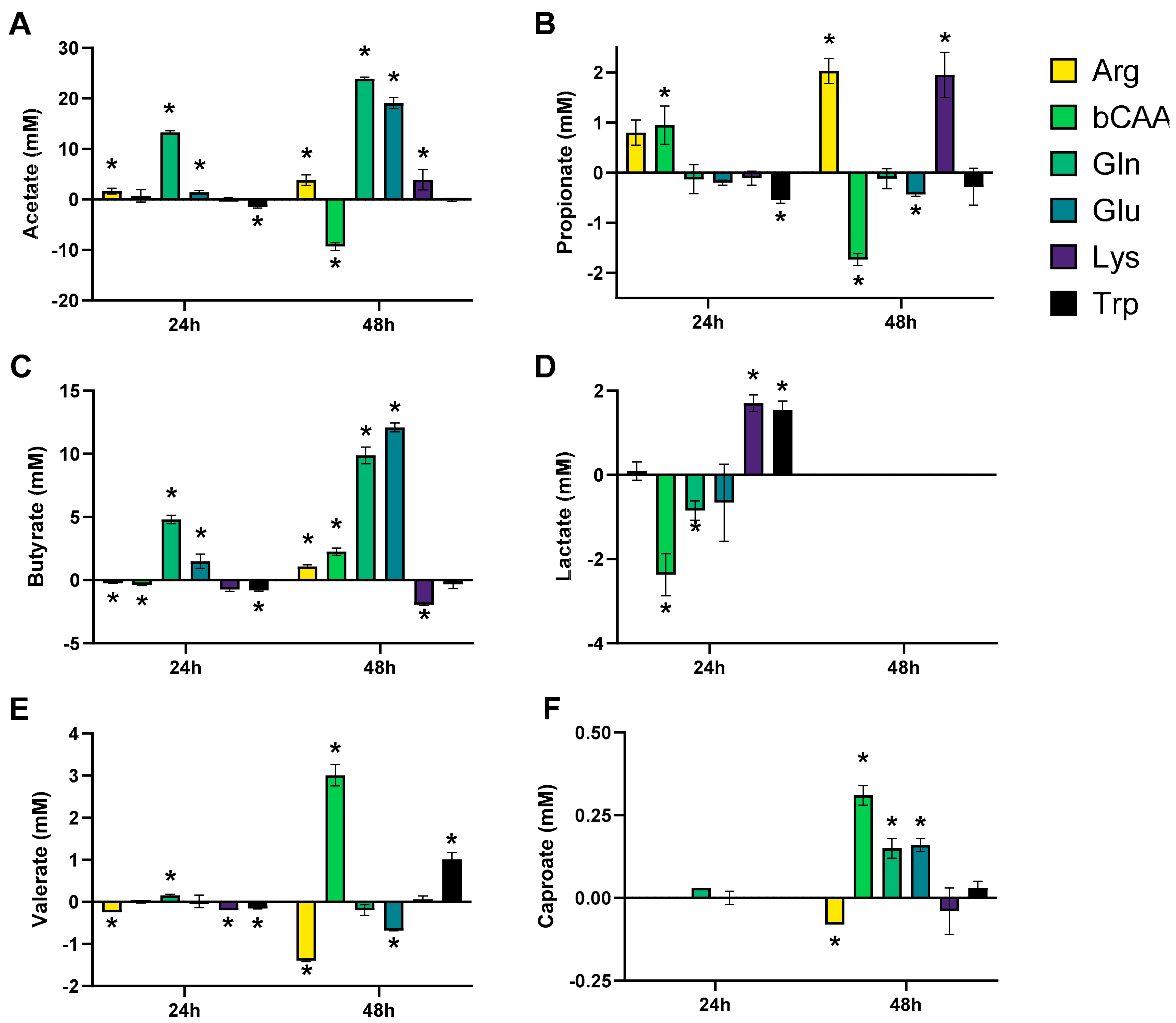
3.1. Amino Acids Differentially Stimulated Microbial Activity of the Porcine Colonic Microbiota
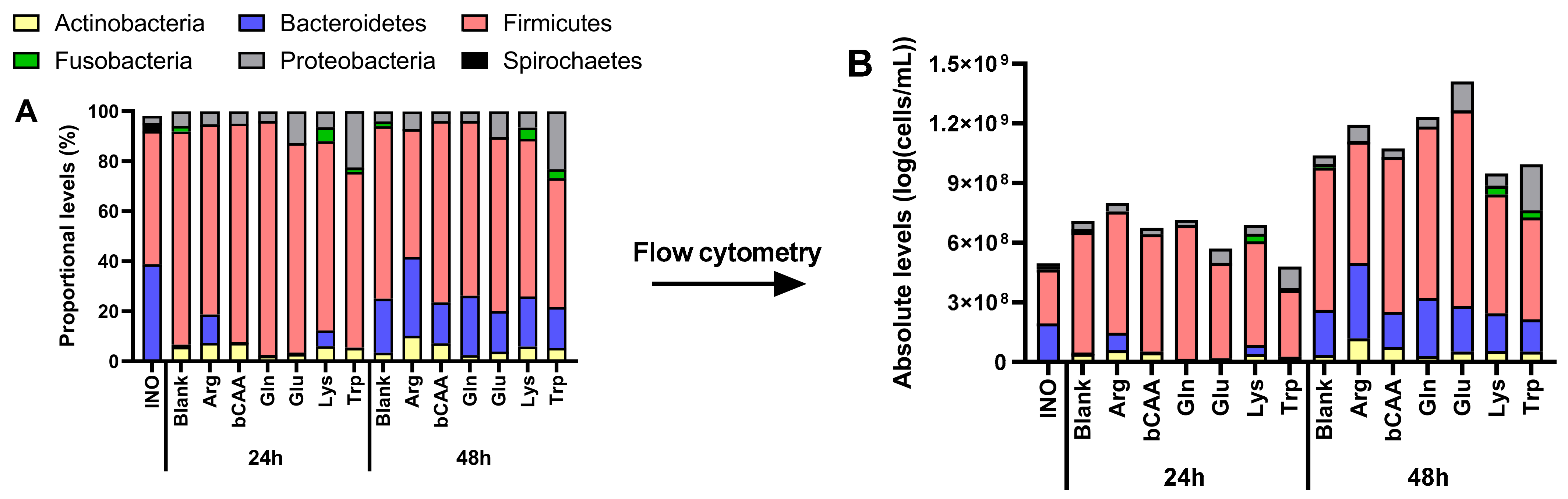
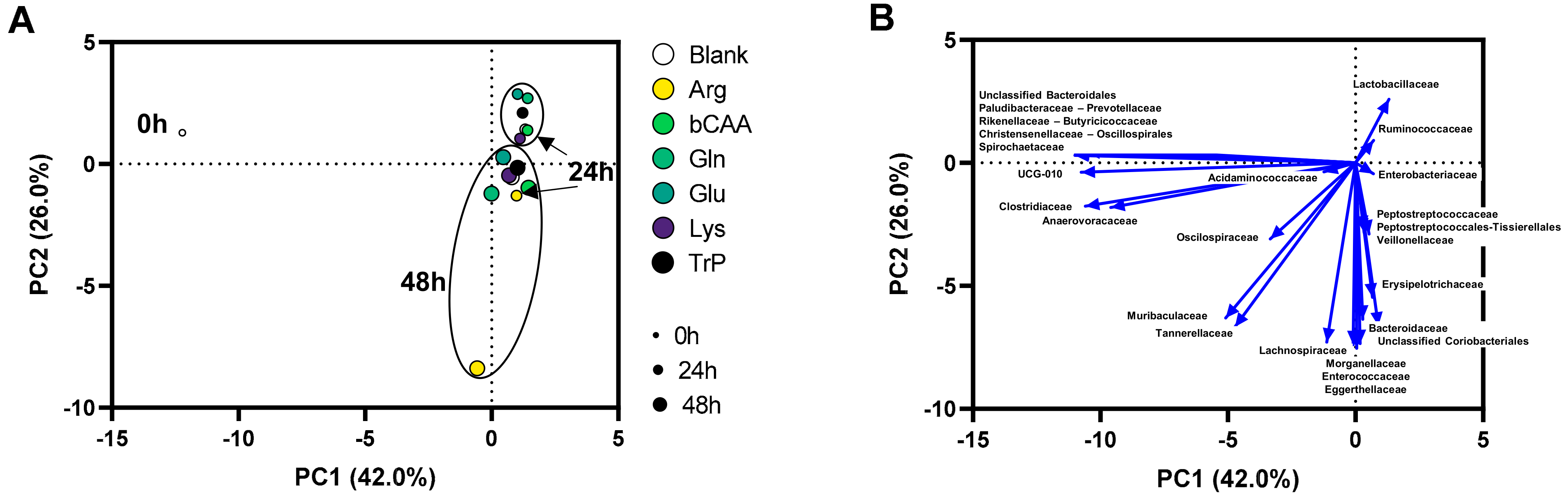
3.2. Amino Acids Differentially Altered Microbial Composition of the Porcine Colonic Microbiota
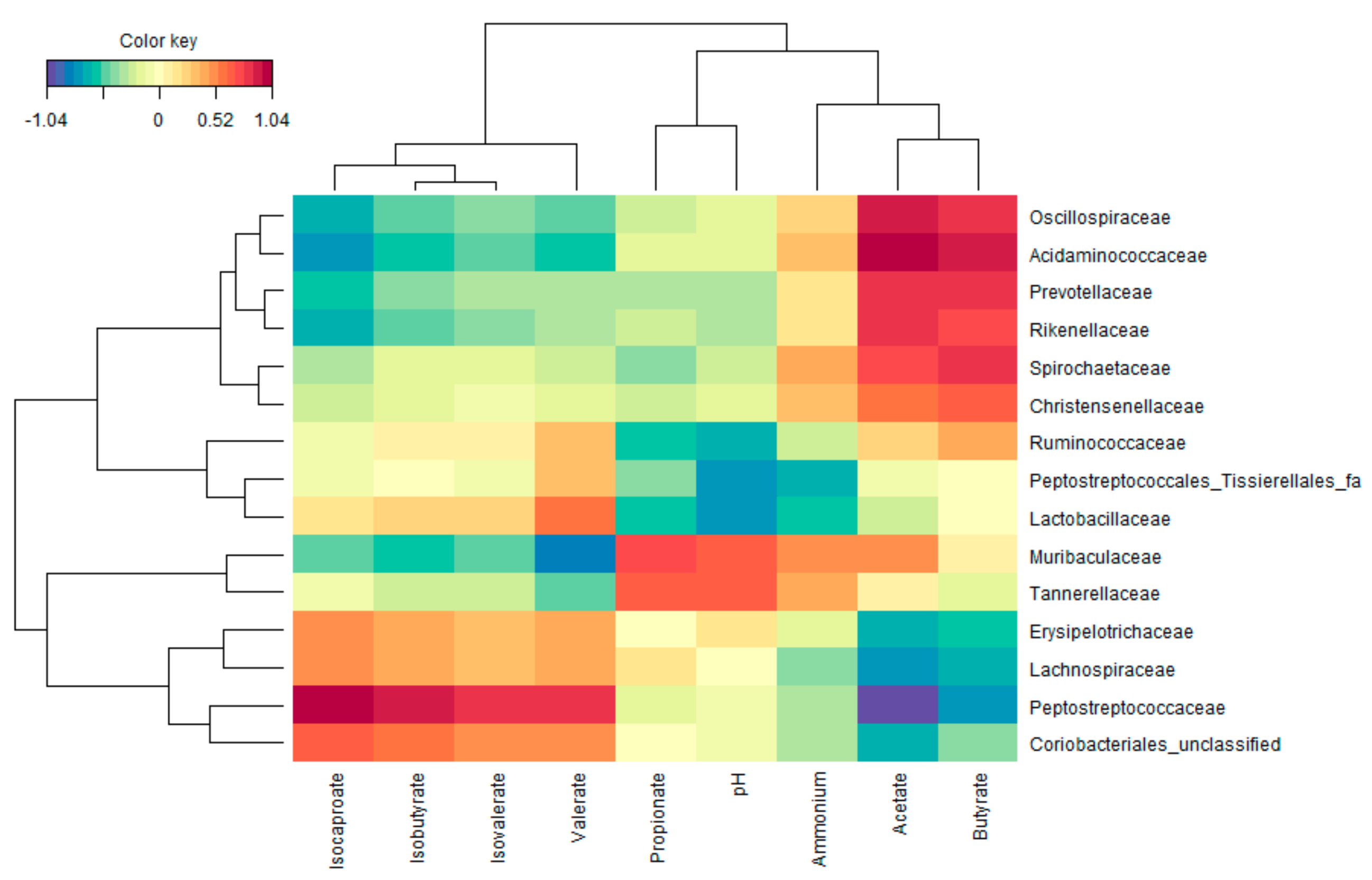
3.3. Functional Populations Involved in the Fermentation of Amino Acids
3.4. In Vitro Microbiota Composition in Comparison with the Original Porcine Colonic Inoculum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chalvon-Demersay, T.; Luise, D.; Le Floc’h, N.; Tesseraud, S.; Lambert, W.; Bosi, P.; Trevisi, P.; Beaumont, M.; Corrent, E. Functional Amino Acids in Pigs and Chickens: Implication for Gut Health. Front. Vet. Sci. 2021, 8, 496. [Google Scholar] [CrossRef] [PubMed]
- Eugenio, F.A.; van Milgen, J.; Duperray, J.; Sergheraert, R.; Le Floc’h, N. Feeding Intact Proteins, Peptides, or Free Amino Acids to Monogastric Farm Animals. Amino Acids 2022, 54, 157–168. [Google Scholar] [CrossRef]
- Ren, W.; Chen, S.; Yin, J.; Duan, J.; Li, T.; Liu, G.; Feng, Z.; Tan, B.; Yin, Y.; Wu, G. Dietary Arginine Supplementation of Mice Alters the Microbial Population and Activates Intestinal Innate Immunity. J. Nutr. 2014, 144, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Duan, J.; Yin, J.; Liu, G.; Cao, Z.; Xiong, X.; Chen, S.; Li, T.; Yin, Y.; Hou, Y.; et al. Dietary L-Glutamine Supplementation Modulates Microbial Community and Activates Innate Immunity in the Mouse Intestine. Amino Acids 2014, 46, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Huang, S.; Zou, D.; Dong, D.; He, X.; Liu, N.; Liu, W.; Huang, L. Metabolic Shifts and Structural Changes in the Gut Microbiota upon Branched-Chain Amino Acid Supplementation in Middle-Aged Mice. Amino Acids 2016, 48, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Bertocchi, M.; Bosi, P.; Correa, F.; Spinelli, E.; Trevisi, P. Contribution of L-Arginine Supplementation during Gestation on Sow Productive Performance and on Sow Microbial Faecal Profile. Ital. J. Anim. Sci. 2020, 19, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.-M.; Li, T.-J.; Wu, L.; Xiao, D.-F.; Blachier, F.; Yin, Y.-L. Monosodium L-Glutamate and Dietary Fat Differently Modify the Composition of the Intestinal Microbiota in Growing Pigs. Obes. Facts 2015, 8, 87–100. [Google Scholar] [CrossRef]
- Zhang, B.; Lv, Z.; Li, Z.; Wang, W.; Li, G.; Guo, Y. Dietary L-Arginine Supplementation Alleviates the Intestinal Injury and Modulates the Gut Microbiota in Broiler Chickens Challenged by Clostridium Perfringens. Front. Microbiol. 2018, 9, 1716. [Google Scholar] [CrossRef] [Green Version]
- Bello, A.U.; Idrus, Z.; Meng, G.Y.; Awad, E.A.; Farjam, A.S. Gut Microbiota and Transportation Stress Response Affected by Tryptophan Supplementation in Broiler Chickens. Ital. J. Anim. Sci. 2018, 17, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Adhikari, P.; Oakley, B.; Kim, W.K. Changes in Cecum Microbial Community in Response to Total Sulfur Amino Acid (TSAA: DL-Methionine) in Antibiotic-Free and Supplemented Poultry Birds. Poult. Sci. 2019, 98, 5809–5819. [Google Scholar] [CrossRef]
- Suenderhauf, C.; Tuffin, G.; Lorentsen, H.; Grimm, H.-P.; Flament, C.; Parrott, N. Pharmacokinetics of Paracetamol in Göttingen Minipigs: In Vivo Studies and Modeling to Elucidate Physiological Determinants of Absorption. Pharm. Res. 2014, 31, 2696–2707. [Google Scholar] [CrossRef]
- Hossain, M.; Abramowitz, W.; Watrous, B.J.; Szpunar, G.J.; Ayres, J.W. Gastrointestinal Transit of Nondisintegrating, Nonerodible Oral Dosage Forms in Pigs. Pharm. Res. 1990, 7, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Chen, X.; Fang, C.; Zhao, L.; Chen, F. The Maturing Development of Gut Microbiota in Commercial Piglets during the Weaning Transition. Front. Microbiol. 2017, 8, 1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.-Y.; Williams, B.; Konstantinov, S.; Tamminga, S.; De Vos, W.; Akkermans, A. Analysis of 16S RDNA Reveals Bacterial Shift during in Vitro Fermentation of Fermentable Carbohydrate Using Piglet Faeces as Inoculum. Anaerobe 2003, 9, 175–180. [Google Scholar] [CrossRef]
- Messens, W.; Goris, J.; Dierick, N.; Herman, L.; Heyndrickx, M. Inhibition of Salmonella Typhimurium by Medium-Chain Fatty Acids in an in Vitro Simulation of the Porcine Cecum. Vet. Microbiol. 2010, 141, 73–80. [Google Scholar] [CrossRef]
- Tanner, S.A.; Berner, A.Z.; Rigozzi, E.; Grattepanche, F.; Chassard, C.; Lacroix, C. In Vitro Continuous Fermentation Model (PolyFermS) of the Swine Proximal Colon for Simultaneous Testing on the Same Gut Microbiota. PLoS ONE 2014, 9, e94123. [Google Scholar] [CrossRef] [Green Version]
- Vandeputte, D.; Kathagen, G.; D’hoe, K.; Vieira-Silva, S.; Valles-Colomer, M.; Sabino, J.; Wang, J.; Tito, R.Y.; De Commer, L.; Darzi, Y.; et al. Quantitative Microbiome Profiling Links Gut Community Variation to Microbial Load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef]
- Props, R.; Kerckhof, F.-M.; Rubbens, P.; De Vrieze, J.; Sanabria, E.H.; Waegeman, W.; Monsieurs, P.; Hammes, F.; Boon, N. Absolute Quantification of Microbial Taxon Abundances. ISME J. 2017, 11, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Van den Abbeele, P.; Moens, F.; Pignataro, G.; Schnurr, J.; Ribecco, C.; Gramenzi, A.; Marzorati, M. Yeast-Derived Formulations Are Differentially Fermented by the Canine and Feline Microbiome as Assessed in a Novel In Vitro Colonic Fermentation Model. J. Agric. Food Chem. 2020, 68, 13102–13110. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Verstrepen, L.; Ghyselinck, J.; Albers, R.; Marzorati, M.; Mercenier, A. A Novel Non-Digestible, Carrot-Derived Polysaccharide (CRG-I) Selectively Modulates the Human Gut Microbiota While Promoting Gut Barrier Integrity: An Integrated in Vitro Approach. Nutrients 2020, 12, 1917. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Duysburgh, C.; Ghyselinck, J.; Goltz, S.; Berezhnaya, Y.; Boileau, T.; De Blaiser, A.; Marzorati, M. Fructans with Varying Degree of Polymerization Enhance the Selective Growth of Bifidobacterium Animalis Subsp. Lactis BB-12 in the Human Gut Microbiome In Vitro. Appl. Sci. 2021, 11, 598. [Google Scholar] [CrossRef]
- Marsaux, B.; Van den Abbeele, P.; Ghyselinck, J.; Prioult, G.; Marzorati, M.; Bogićević, B. Synbiotic Effect of Bifidobacterium Lactis CNCM I-3446 and Bovine Milk-Derived Oligosaccharides on Infant Gut Microbiota. Nutrients 2020, 12, 2268. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Taminiau, B.; Pinheiro, I.; Duysburgh, C.; Jacobs, H.; Pijls, L.; Marzorati, M. Arabinoxylo-Oligosaccharides and Inulin Impact Inter-Individual Variation on Microbial Metabolism and Composition, Which Immunomodulates Human Cells. J. Agric. Food Chem. 2018, 66, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Van Noten, N.; Degroote, J.; Van Liefferinge, E.; Taminiau, B.; De Smet, S.; Desmet, T.; Michiels, J. Effects of Thymol and Thymol α-D-Glucopyranoside on Intestinal Function and Microbiota of Weaned Pigs. Animals 2020, 10, 329. [Google Scholar] [CrossRef] [Green Version]
- Hoefman, S.; Pommerening-Röser, A.; Samyn, E.; De Vos, P.; Heylen, K. Efficient Cryopreservation Protocol Enables Accessibility of a Broad Range of Ammonia-Oxidizing Bacteria for the Scientific Community. Res. Microbiol. 2013, 164, 288–292. [Google Scholar] [CrossRef]
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.W.; Boschker, H.T.S.; Verstraete, W.; Van de Wiele, T. Human Faecal Microbiota Display Variable Patterns of Glycerol Metabolism. FEMS Microbiol. Ecol. 2010, 74, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Van de Wiele, T.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Prebiotic Effects of Chicory Inulin in the Simulator of the Human Intestinal Microbial Ecosystem. FEMS Microbiol. Ecol. 2004, 51, 143–153. [Google Scholar] [CrossRef]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a Tool to Protect the Structure and Function of an Activated-Sludge Microbial Community against a 3-Chloroaniline Shock Load. Appl. Environ. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Duysburgh, C.; Van den Abbeele, P.; Krishnan, K.; Bayne, T.F.; Marzorati, M. A Synbiotic Concept Containing Spore-Forming Bacillus Strains and a Prebiotic Fiber Blend Consistently Enhanced Metabolic Activity by Modulation of the Gut Microbiome in Vitro. Int. J. Pharm. X 2019, 1, 100021. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L. Assessing and Improving Methods Used in Operational Taxonomic Unit-Based Approaches for 16S RRNA Gene Sequence Analysis. Appl. Environ. Microbiol. 2011, 77, 3219–3226. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved Alignments and New Tools for RRNA Analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Lee, S.; Lee, D.K. What Is the Proper Way to Apply the Multiple Comparison Test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.-A.L. MixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Engelhardt, W.; Ronnau, K.; Rechkemmer, G.; Sakata, T. Absorption of Short-Chain Fatty Acids and Their Role in the Hindgut of Monogastric Animals. Anim. Feed Sci. Technol. 1989, 23, 43–53. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Chapelet, G.; Javaudin, F.; Lepelletier, D.; Batard, E.; Montassier, E. The Effects of Inulin on Gut Microbial Composition: A Systematic Review of Evidence from Human Studies. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 403–413. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, T.R.; Cotta, M.A.; Falsen, E.; Moore, E.; Lawson, P.A. Peptostreptococcus Russellii Sp. Nov., Isolated from a Swine-Manure Storage Pit. Int. J. Syst. Evol. Microbiol. 2011, 61, 1875–1879. [Google Scholar] [CrossRef]
- Hinnebusch, B.F.; Meng, S.; Wu, J.T.; Archer, S.Y.; Hodin, R.A. The Effects of Short-Chain Fatty Acids on Human Colon Cancer Cell Phenotype Are Associated with Histone Hyperacetylation. J. Nutr. 2002, 132, 1012–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, J.A.K.; Mullish, B.H.; Pechlivanis, A.; Liu, Z.; Brignardello, J.; Kao, D.; Holmes, E.; Li, J.V.; Clarke, T.B.; Thursz, M.R.; et al. Inhibiting Growth of Clostridioides Difficile by Restoring Valerate, Produced by the Intestinal Microbiota. Gastroenterology 2018, 155, 1495–1507.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachier, F.; Mariotti, F.; Huneau, J.F.; Tomé, D. Effects of Amino Acid-Derived Luminal Metabolites on the Colonic Epithelium and Physiopathological Consequences. Amino Acids 2007, 33, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Jamin, A.; Chatelais, L.; Gras-Le Guen, C.; Michel, C.; Le Huërou-Luron, I. Dietary Protein Excess during Neonatal Life Alters Colonic Microbiota and Mucosal Response to Inflammatory Mediators Later in Life in Female Pigs. J. Nutr. 2013, 143, 1225–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falony, G.; Vlachou, A.; Verbrugghe, K.; Vuyst, L.D. Cross-Feeding between Bifidobacterium Longum BB536 and Acetate-Converting, Butyrate-Producing Colon Bacteria during Growth on Oligofructose. Appl. Environ. Microbiol. 2006, 72, 7835–7841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckel, W. Unusual Enzymes Involved in Five Pathways of Glutamate Fermentation. Appl. Microbiol. Biotechnol. 2001, 57, 263–273. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Morotomi, M.; Nagai, F.; Watanabe, Y. Description of Christensenella Minuta Gen. Nov., Sp. Nov., Isolated from Human Faeces, Which Forms a Distinct Branch in the Order Clostridiales, and Proposal of Christensenellaceae Fam. Nov. Int. J. Syst. Evol. Microbiol. 2011, 62, 144–149. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A Candidate for the next-Generation Probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and Cultivation Study of Muribaculaceae Reveals Novel Species, Host Preference, and Functional Potential of This yet Undescribed Family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ormerod, K.L.; Wood, D.L.A.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.O.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic Characterization of the Uncultured Bacteroidales Family S24-7 Inhabiting the Guts of Homeothermic Animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van de Wiele, T. Propionate as a Health-Promoting Microbial Metabolite in the Human Gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of Protein Fermentation to Gut Health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Family | Arg | bCAA | Gln | Glu | Lys | Trp |
|---|---|---|---|---|---|---|---|
| Actinobacteria | Unclassified Coriobacteriales | 0.61 | 0.46 | −0.18 | 0.03 | 0.32 | 0.32 |
| Eggerthellaceae | 1.30 | 0.59 | 0.17 | 0.04 | −0.08 | 0.07 | |
| Bacteroidetes | Bacteroidaceae | 0.27 | 0.00 | 0.01 | −0.09 | 0.01 | −0.03 |
| Unclassified Bacteroidales | 0.22 | −0.03 | 0.06 | −0.05 | −0.03 | −0.03 | |
| Muribaculaceae | 2.17 | −0.14 | 0.82 | 0.04 | 1.48 | −0.43 | |
| Paludibacteraceae | −0.14 | −0.34 | 0.27 | 0.59 | −0.24 | −0.34 | |
| Prevotellaceae | 0.11 | −0.24 | 0.32 | 0.20 | −0.08 | −0.32 | |
| Rikenellaceae | −0.06 | −0.53 | 0.26 | 0.10 | −0.23 | −0.48 | |
| Tannerellaceae | 0.93 | −0.63 | 0.34 | −0.34 | 0.12 | −1.35 | |
| Firmicutes | Acidaminococcaceae | 0.24 | −0.60 | 1.07 | 1.27 | 0.14 | −0.46 |
| Anaerovoracaceae | 0.40 | −0.18 | 0.46 | 0.22 | −0.01 | 0.51 | |
| Butyricicoccaceae | 0.07 | −0.16 | 0.10 | 0.35 | −0.28 | 0.06 | |
| Christensenellaceae | 0.26 | −0.08 | 0.13 | −0.10 | −0.01 | −0.46 | |
| Clostridiaceae | 0.29 | −0.34 | 0.08 | 0.29 | −0.20 | −0.16 | |
| Enterococcaceae | 1.54 | 0.00 | −0.16 | −0.44 | −0.30 | −0.12 | |
| Erysipelotrichaceae | 0.25 | −0.01 | −0.01 | −1.04 | −0.23 | −0.25 | |
| Lachnospiraceae | 0.26 | −0.06 | −0.02 | −0.39 | −0.03 | −0.15 | |
| Lactobacillaceae | −0.55 | −0.13 | −0.09 | 0.02 | −0.26 | 0.05 | |
| Oscillospiraceae | 0.44 | −0.21 | 0.49 | 0.52 | −0.04 | 0.05 | |
| Oscillospirales | 0.37 | −0.14 | 0.25 | −0.19 | −0.14 | −0.02 | |
| Peptostreptococcaceae | 0.60 | 1.18 | −0.43 | −0.71 | 0.33 | 0.61 | |
| Peptostreptococcales−Tissierellales | −0.36 | −0.26 | −0.11 | −0.22 | −0.14 | −0.02 | |
| Ruminococcaceae | −0.26 | −0.14 | 0.26 | 0.44 | −0.23 | −0.16 | |
| UCG−010 | −0.03 | −0.40 | −0.07 | −0.60 | 0.09 | −0.37 | |
| Veillonellaceae | −0.24 | 0.00 | −0.03 | 0.06 | 0.07 | −0.52 | |
| Proteobacteria | Enterobacteriaceae | 0.27 | 0.02 | 0.00 | 0.52 | 0.22 | 0.79 |
| Morganellaceae | 0.73 | 0.52 | −0.49 | −1.03 | −0.17 | 0.43 | |
| Spirochaetes | Spirochaetaceae | 0.35 | −0.03 | 0.14 | 0.07 | −0.06 | −0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van den Abbeele, P.; Ghyselinck, J.; Marzorati, M.; Koch, A.-M.; Lambert, W.; Michiels, J.; Chalvon-Demersay, T. The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota. Microorganisms 2022, 10, 762. https://doi.org/10.3390/microorganisms10040762
Van den Abbeele P, Ghyselinck J, Marzorati M, Koch A-M, Lambert W, Michiels J, Chalvon-Demersay T. The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota. Microorganisms. 2022; 10(4):762. https://doi.org/10.3390/microorganisms10040762
Chicago/Turabian StyleVan den Abbeele, Pieter, Jonas Ghyselinck, Massimo Marzorati, Anna-Maria Koch, William Lambert, Joris Michiels, and Tristan Chalvon-Demersay. 2022. "The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota" Microorganisms 10, no. 4: 762. https://doi.org/10.3390/microorganisms10040762
APA StyleVan den Abbeele, P., Ghyselinck, J., Marzorati, M., Koch, A.-M., Lambert, W., Michiels, J., & Chalvon-Demersay, T. (2022). The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota. Microorganisms, 10(4), 762. https://doi.org/10.3390/microorganisms10040762