
Geo-Spatial Characteristics of 567 Places of Tick-Borne Encephalitis Infection in Southern Germany, 2018–2020

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Places of Infection
2.2. Confirmed Natural TBE Foci
2.3. Comparator Places
2.4. Covariates
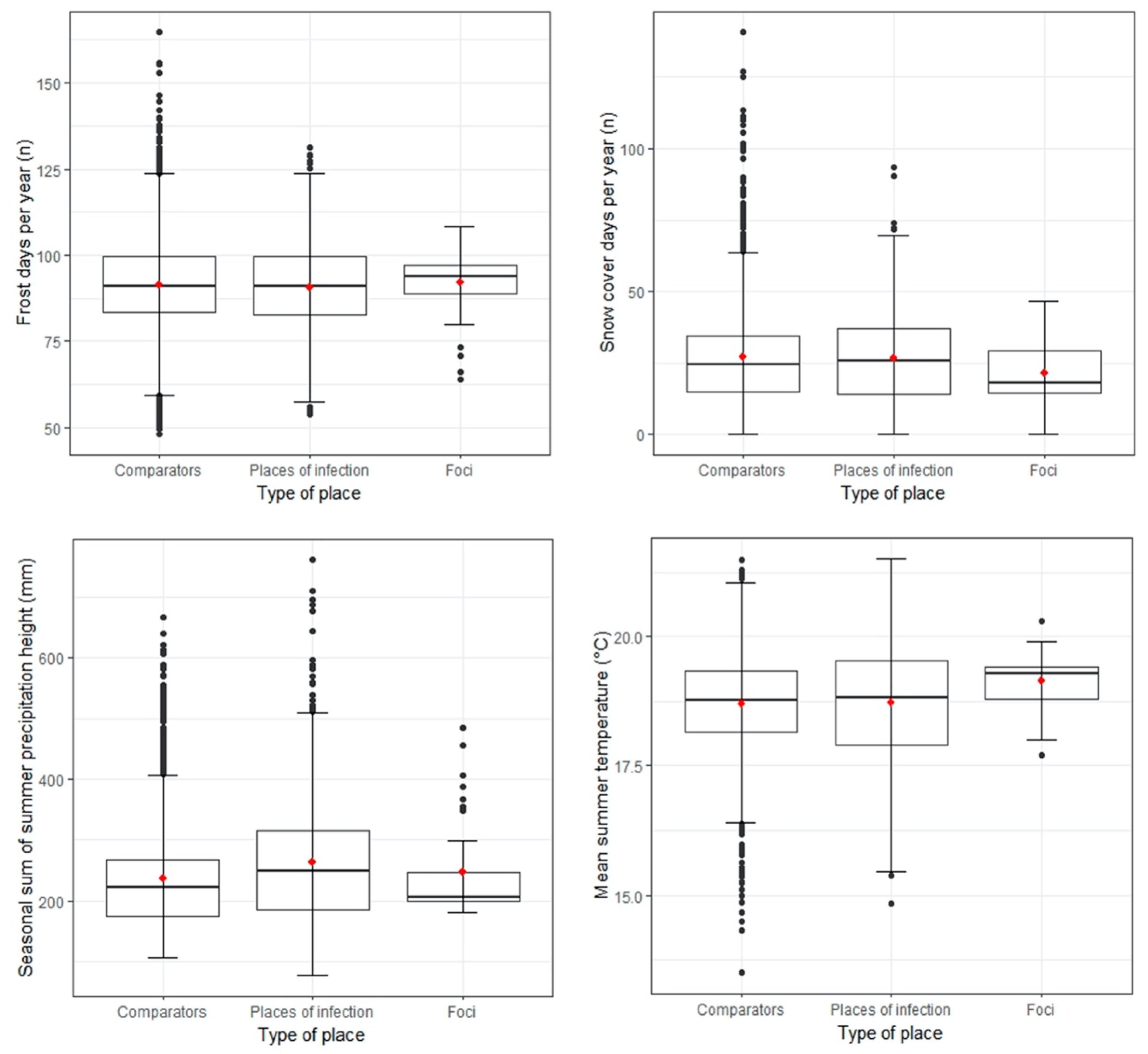
2.4.1. Abiotic Factors
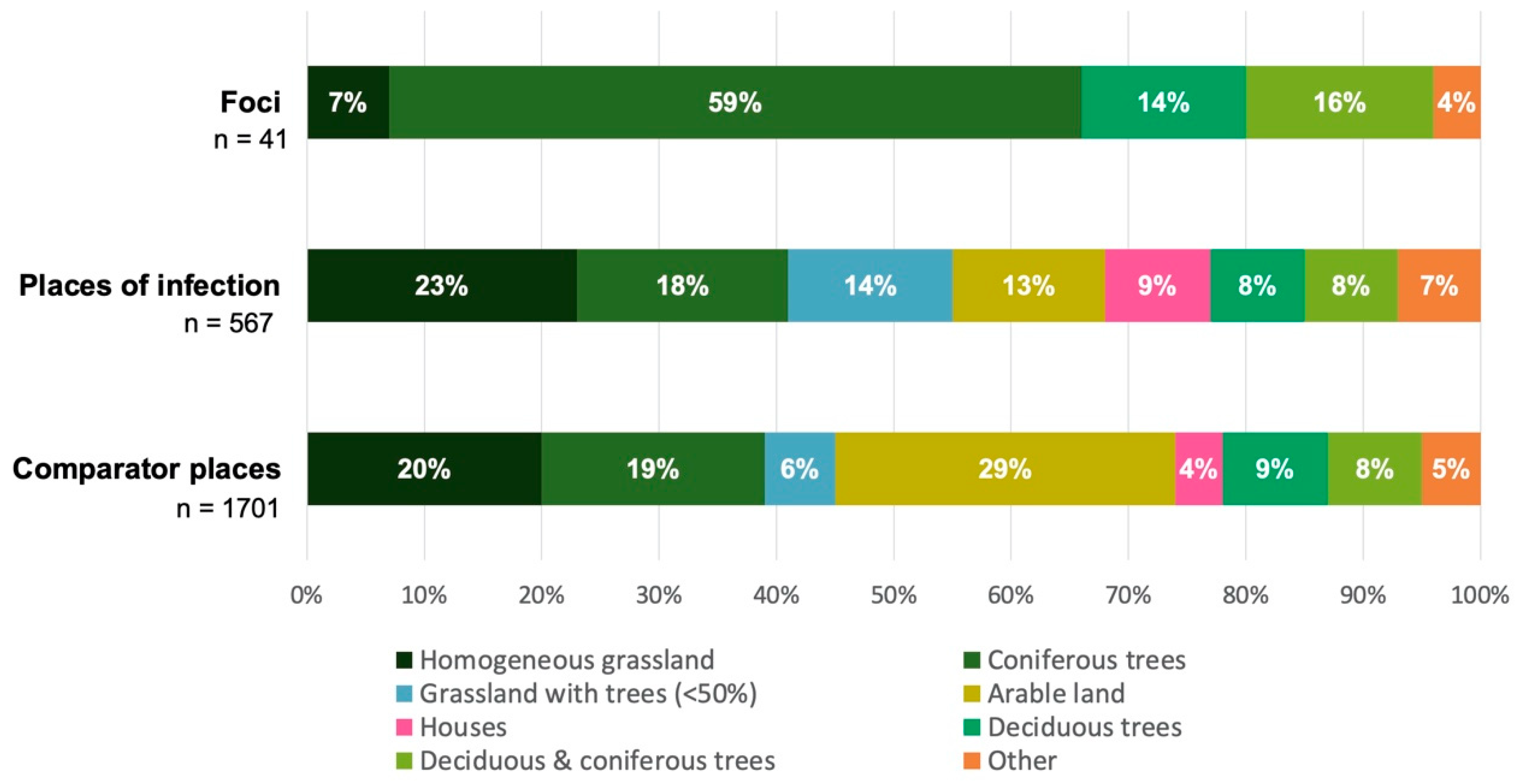
2.4.2. Biotic and Anthropogenic Factors
2.5. Spatial Analysis
3. Results
3.1. Epidemiological Overview and Descriptive Statistics
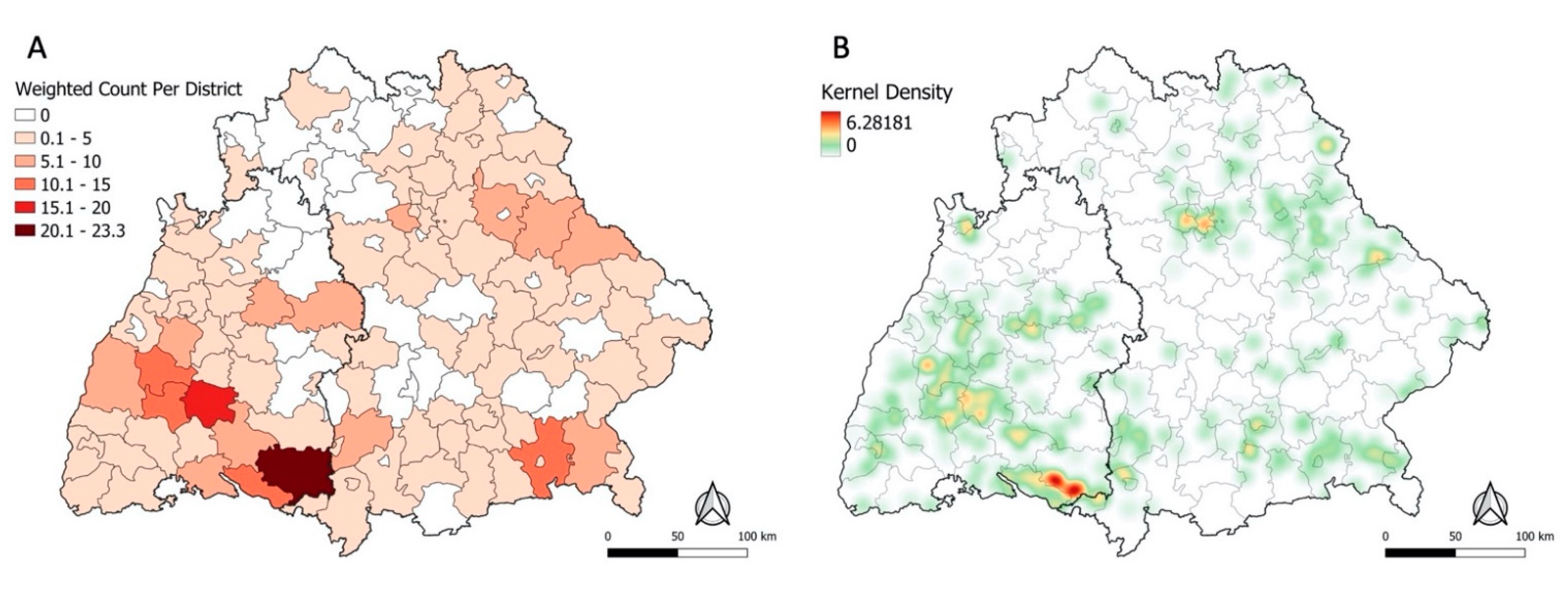
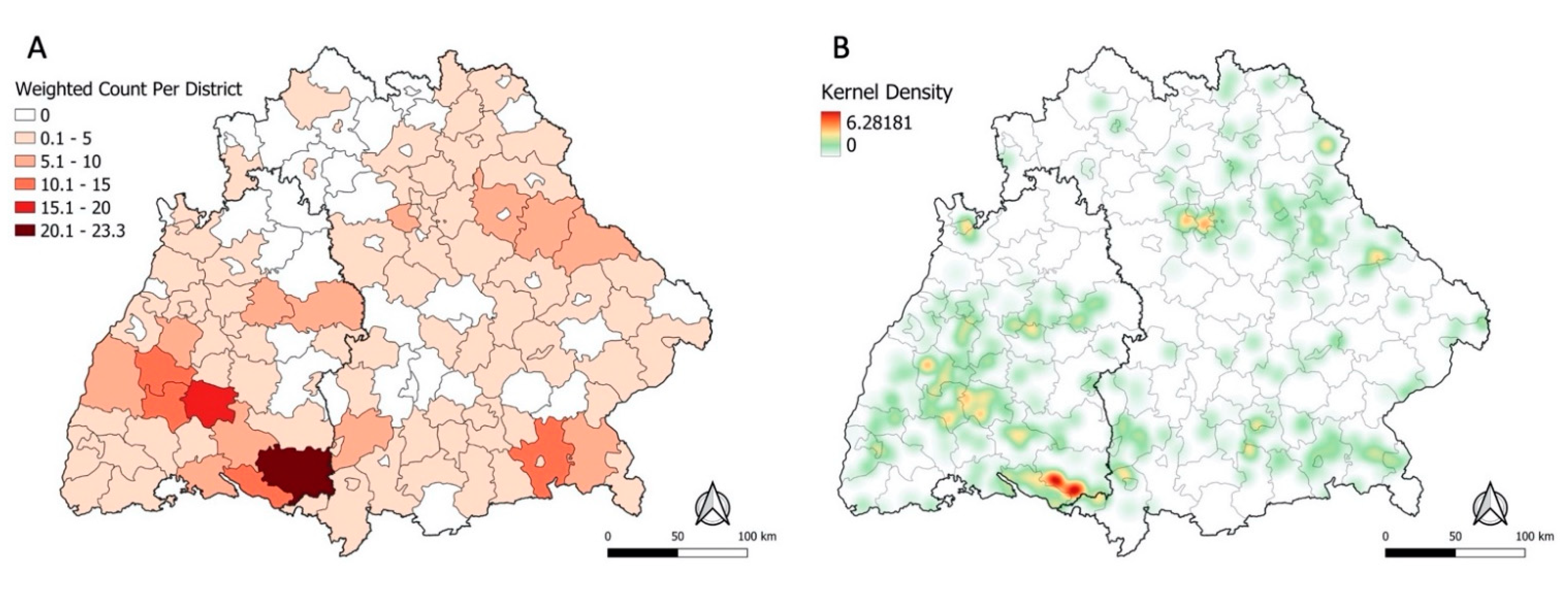
3.2. Spatial Clustering of Places of TBE Infection
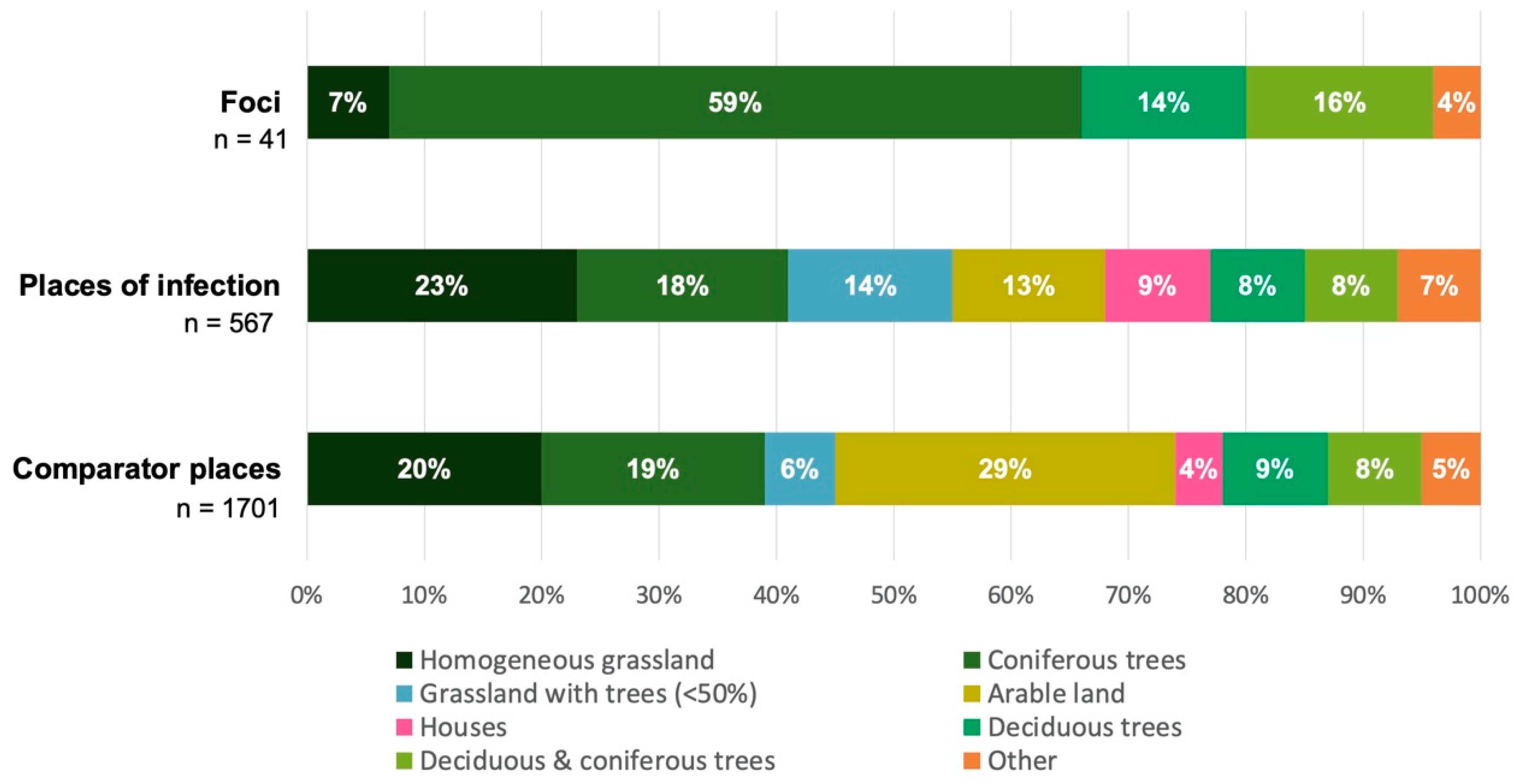
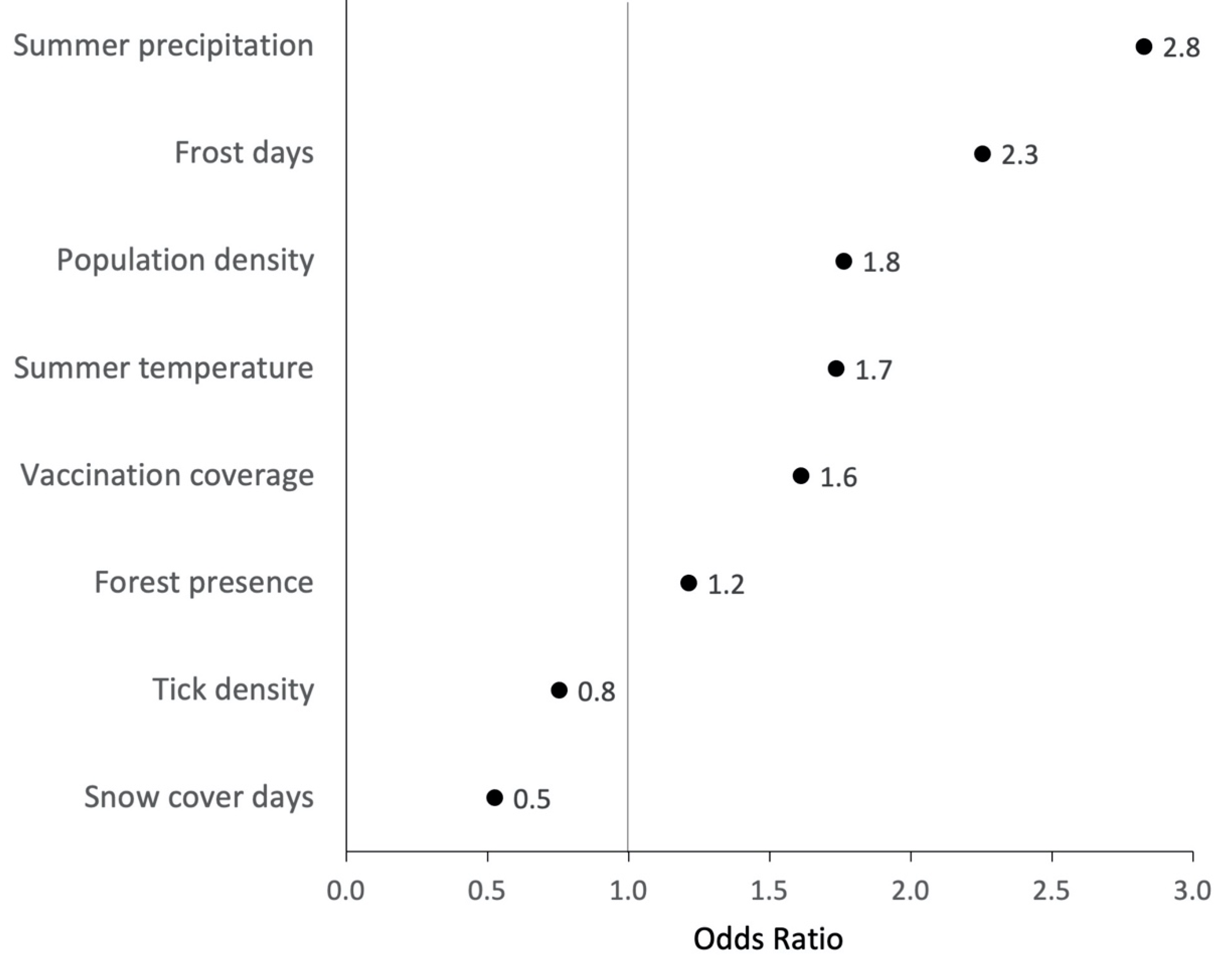
3.3. Individual Environmental Characteristics of Places of TBE Infection
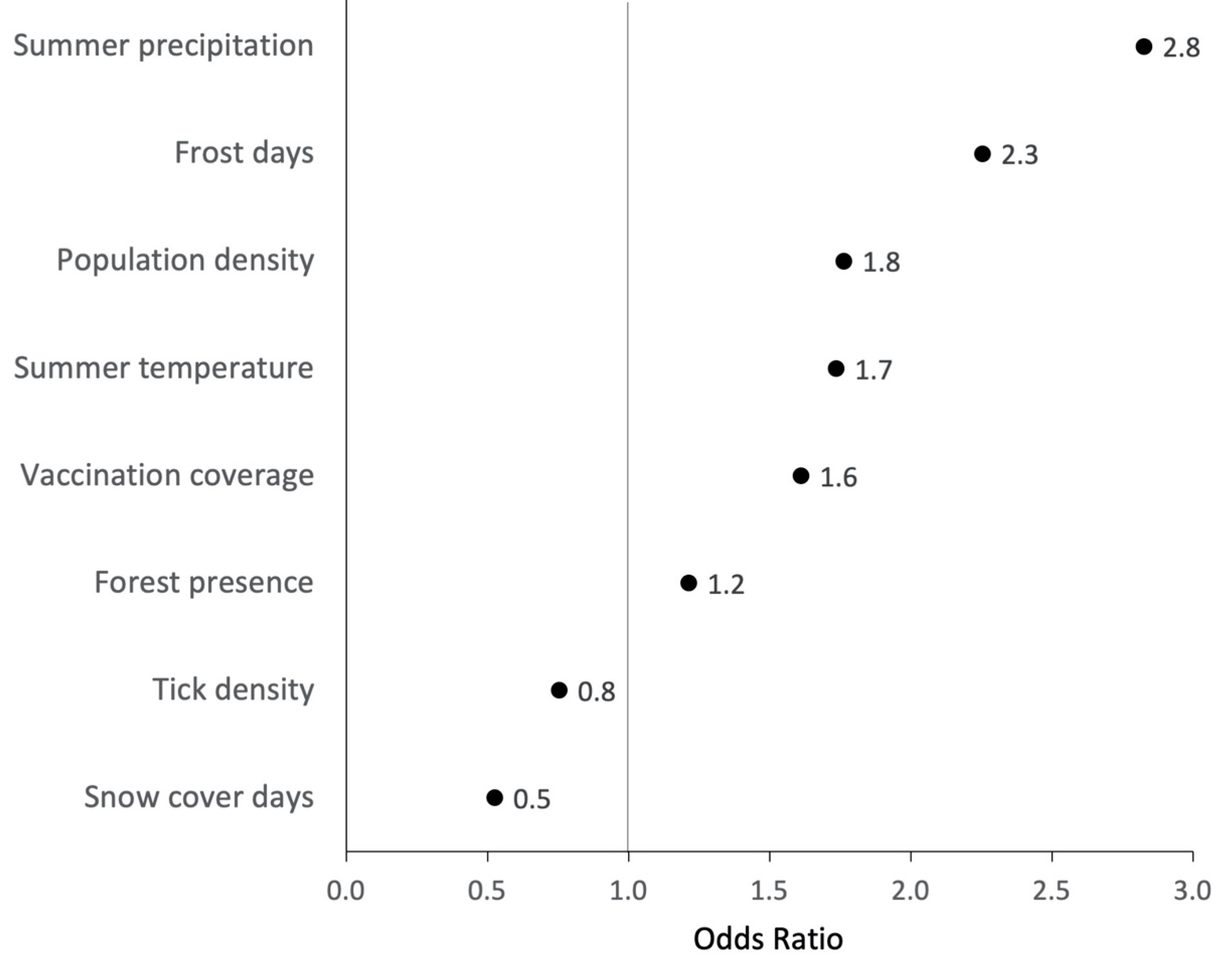
3.4. Ecological Niche Model
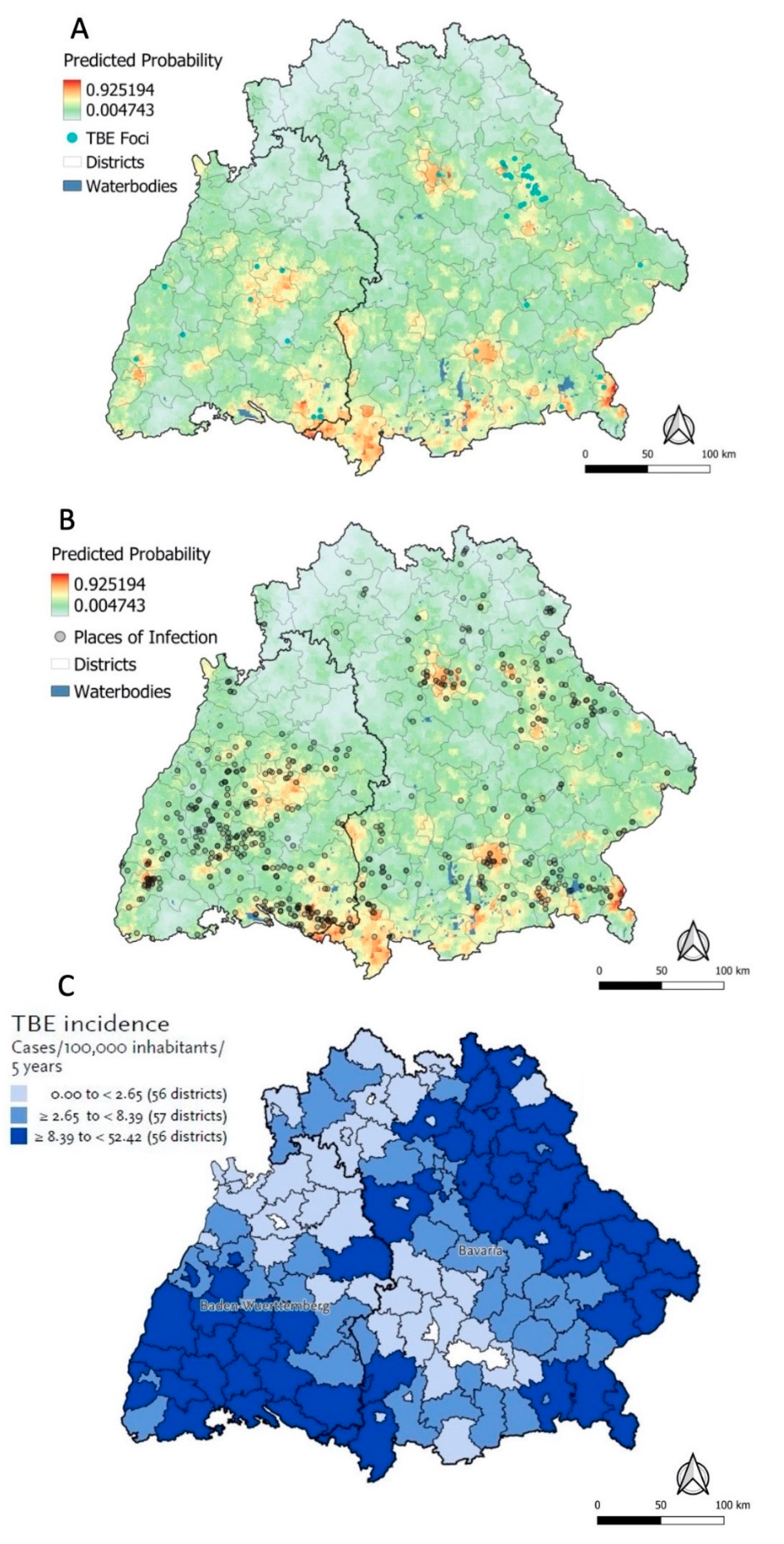
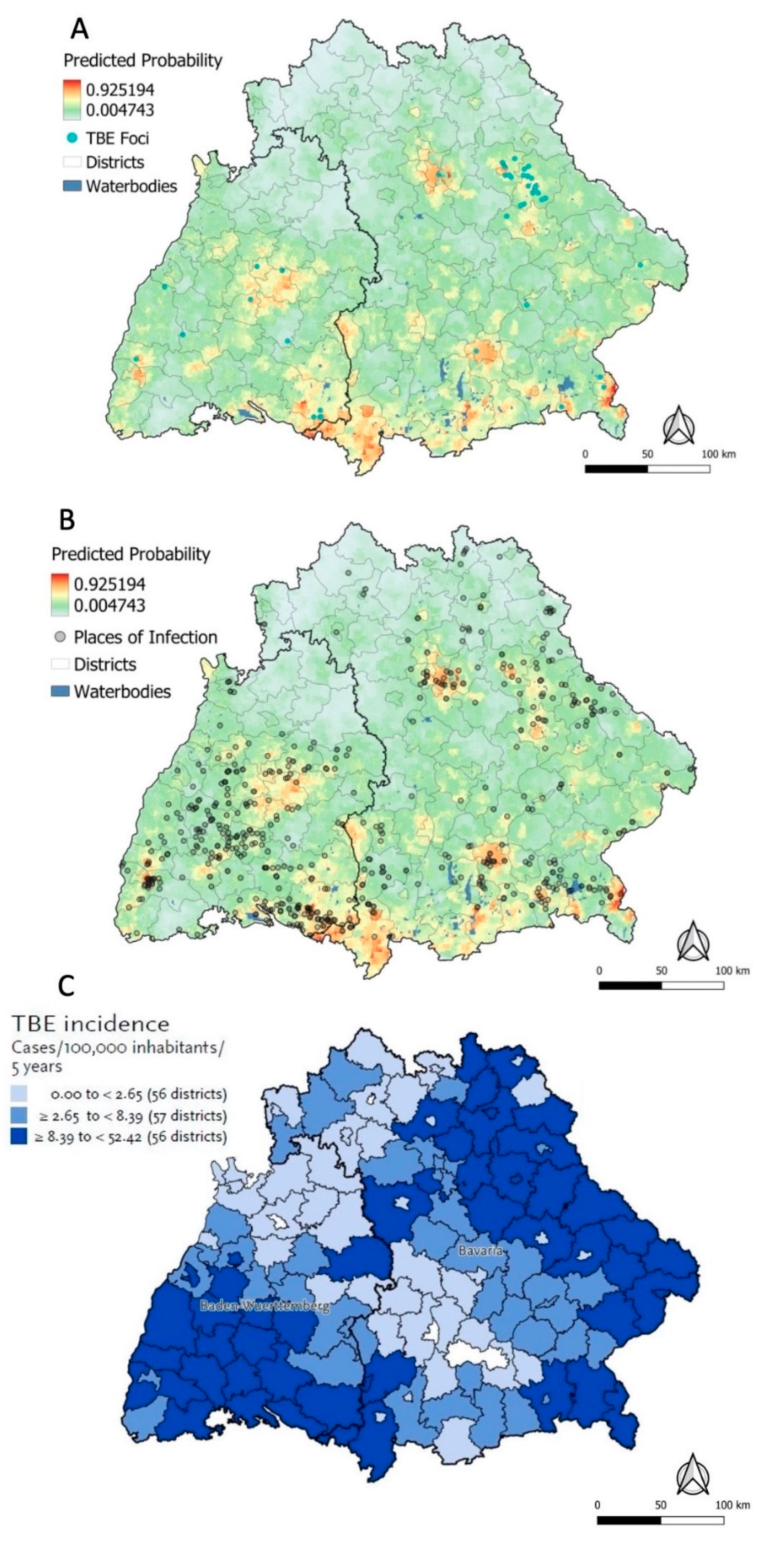
3.4.1. Predicted Probabilities for Places of TBE Infection
3.4.2. Sensitivity Analysis and Kernel Density Estimation
3.4.3. RKI Risk Map and Ecological Niche Predictions
4. Discussion
4.1. Strengths
4.2. Limitations and Recommendations for Future Research
4.3. Public Health Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, R.-X.; Lai, S.-J.; Yang, Y.; Li, X.-L.; Liu, K.; Yao, H.-W.; Zhou, H.; Li, Y.; Wang, L.-P.; Mu, D.; et al. Mapping the distribution of tick-borne encephalitis in mainland China. Ticks Tick Borne Dis. 2017, 8, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Bogovic, P. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, W.; Kreusch, T.; Böhmer, M.M.; Wagner-Wiening, C.; Dobler, G.; Wichmann, O.; Altmann, D. Epidemiology of Tick-Borne Encephalitis (TBE) in Germany, 2001–2018. Pathogens 2019, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert Koch Institut. Frühsommer-Meningoenzephalitis (FSME) und Verwandte Virusenzephalitiden (TBE, Tick-Borne Encephalitis). Available online: https://www.rki.de/DE/Content/Infekt/EpidBull/Merkblaetter/Ratgeber_FSME.html;jsessionid=36F76FE0F81E448D6F53E0EF2E18EBFB.internet072#doc2381918bodyText7 (accessed on 8 September 2021).
- World Health Organization. Tick-Borne Encephalitis. Available online: https://www.who.int/health-topics/tick-borne-encephalitis#tab=tab_1 (accessed on 9 September 2021).
- Beauté, J.; Spiteri, G.; Warns-Petit, E.; Zeller, H. Tick-borne encephalitis in Europe, 2012 to 2016. Eurosurveillance 2018, 23, 1800201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert Koch Institut. FSME Risikogebiete in Deutschland (Stand: Januar 2022). Epidemiol. Bull. 2022, 9, 3–21. [Google Scholar] [CrossRef]
- Robert Koch Institut. Zahl der FSME-Erkrankungen in Aktueller Zeckensaison höher als in den Vorjahren. Available online: https://www.rki.de/DE/Content/InfAZ/F/FSME/FSME_2020.html;jsessionid=B455A023EDF815721A2E5FC456170155.internet111 (accessed on 3 August 2021).
- Rekord an FSME-Infektionen: Corona Könnte eine Rolle Spielen, Langfristiger Trend Befürchtet. Available online: https://www.aerzteblatt.de/nachrichten/121901/Rekord-an-FSME-Infektionen-Corona-koennte-eine-Rolle-spielen-langfristiger-Trend-befuerchtet (accessed on 5 September 2021).
- Nygren, T.M.; Pilic, A.; Böhmer, M.M.; Wagner-Wiening, C.; Wichmann, O.; Hellenbrand, W. Tick-Borne Encephalitis: Acute Clinical Manifestations, Severity, and Quality of Life in 581 Cases from Germany, 2018–2020. 2022; manuscript submitted for publication. [Google Scholar]
- Jaenson, T.G.T.; Petersson, E.H.; Jaenson, D.G.E.; Kindberg, J.; Pettersson, J.H.-O.; Hjertqvist, M.; Medlock, J.M.; Bengtsson, H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: Incidence of human TBE correlates with abundance of deer and hares. Parasites Vectors 2018, 11, 477. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; de la Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef]
- Daniel, M.; Danielová, V.; Fialová, A.; Malý, M.; Kříž, B.; Nuttall, P.A. Increased Relative Risk of Tick-Borne Encephalitis in Warmer Weather. Front. Cell. Infect. Microbiol. 2018, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.; Materna, J.; Honig, V.; Metelka, L.; Danielová, V.; Harcarik, J.; Kliegrová, S.; Grubhoffer, L. Vertical Distribution of the Tick Ixodes ricinus and Tick-borne Pathogens in the Northern Moravian Mountains Correlated with Climate Warming (Jeseníky Mts., Czech Republic). Central Eur. J. Public Health 2009, 17, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Hauser, G.; Rais, O.; Cadenas, F.M.; Gonseth, Y.; Bouzelboudjen, M.; Gern, L. Influence of climatic factors on Ixodes ricinus nymph abundance and phenology over a long-term monthly observation in Switzerland (2000–2014). Parasites Vectors 2018, 11, 289. [Google Scholar] [CrossRef]
- Gray, J.S. Ixodes ricinus seasonal activity: Implications of global warming indicated by revisiting tick and weather data. Int. J. Med. Microbiol. 2008, 298 (Suppl. S1), 19–24. [Google Scholar] [CrossRef]
- Knap, N.; Županc, T.A. Factors affecting the ecology of tick-borne encephalitis in Slovenia. Epidemiol. Infect. 2015, 143, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Stefanoff, P.; Rubikowska, B.; Bratkowski, J.; Ustrnul, Z.; Vanwambeke, S.O.; Rosińska, M. A Predictive Model Has Identified Tick-Borne Encephalitis High-Risk Areas in Regions Where No Cases Were Reported Previously, Poland, 1999–2012. Int. J. Environ. Res. Public Health 2018, 15, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uusitalo, R.; Siljander, M.; Dub, T.; Sane, J.; Sormunen, J.J.; Pellikka, P.; Vapalahti, O. Modelling habitat suitability for occurrence of human tick-borne encephalitis (TBE) cases in Finland. Ticks Tick-Borne Dis. 2020, 11, 101457. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Green, R.M.; Peacey, M.F.; Rogers, D.J. Seasonal synchrony: The key to tick-borne encephalitis foci identified by satellite data. Parasitology 2000, 121, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, S.E. The shifting landscape of tick-borne zoonoses: Tick-borne encephalitis and Lyme borreliosis in Europe. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanoff, P.; Zielicka-Hardy, A.; Hlebowicz, M.; Konior, R.; Lipowski, D.; Szenborn, L.; Siennicka, J.; Orlikova, H.; TBE Enhanced Surveillance Working Group. New endemic foci of tick-borne encephalitis (TBE) identified in districts where testing for TBE was not available before 2009 in Poland. Parasites Vectors 2013, 6, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Rubel, F.; Brugger, K. Operational TBE incidence forecasts for Austria, Germany, and Switzerland 2019–2021. Ticks Tick-Borne Dis. 2021, 12, 101579. [Google Scholar] [CrossRef]
- Brugger, K.; Boehnke, D.; Petney, T.; Dobler, G.; Pfeffer, M.; Silaghi, C.; Schaub, G.A.; Pinior, B.; Dautel, H.; Kahl, O.; et al. A Density Map of the Tick-Borne Encephalitis and Lyme Borreliosis Vector Ixodes ricinus (Acari: Ixodidae) for Germany. J. Med. Entomol. 2016, 53, 1292–1302. [Google Scholar] [CrossRef]
- Robert Koch Institut. Karte der FSME-Risikogebiete. Available online: https://www.rki.de/DE/Content/InfAZ/F/FSME/Karte_Tab.html (accessed on 22 September 2021).
- QGIS Development Team. QGIS Geographic Information System. Available online: https://www.qgis.org/de/site/ (accessed on 15 July 2021).
- Dobler, G.; Gniel, D.; Petermann, R.; Pfeffer, M. Epidemiology and distribution of tick-borne encephalitis. Wien. Med. Wochenschr. 2012, 162, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Dobler, G.; Hufert, F.; Pfeffer, M.; Essbauer, S. Tick-Borne Encephalitis: From Microfocus to Human Disease. In Progress in Parasitology; Mehlhorn, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 323–331. [Google Scholar]
- Lindau, A. Untersuchung Viraler, Zoonotischer Erreger in Baden-Württemberg. Ph.D. Thesis, Universität Hohenheim, Stuttgart, Germany, 2019. [Google Scholar]
- Materna, J.; Daniel, M.; Danielová, V. Altitudinal distribution limit of the tick Ixodes ricinus shifted considerably towards higher altitudes in Central Europe: Results of three years monitoring in the Krkonoše Mts. (Czech Republic). Cent. Eur. J. Public Health 2005, 13, 24–28. [Google Scholar] [PubMed]
- Jore, S.; Vanwambeke, S.O.; Slunge, D.; Boman, A.; Krogfelt, K.A.; Jepsen, M.T.; Vold, L. Spatial tick bite exposure and associated risk factors in Scandinavia. Infect. Ecol. Epidemiol. 2020, 10, 1764693. [Google Scholar] [CrossRef] [PubMed]
- Clarke-Crespo, E.; Moreno-Arzate, C.N.; López-González, C.A. Ecological Niche Models of Four Hard Tick Genera (Ixodidae) in Mexico. Animals 2020, 10, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, S.; van Vliet, A.J.; Bron, W.A.; Gassner, F.; Takken, W. High Risk of Tick Bites in Dutch Gardens. Vector-Borne Zoonotic Dis. 2013, 13, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Maier, W.A.; Kistemann, T.; Kampen, H. Analysis of the distribution of the tick Ixodes ricinus L. (Acari: Ixodidae) in a nature reserve of western Germany using Geographic Information Systems. Int. J. Hyg. Environ. Health 2009, 212, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, D.; Brugger, K.; Pfäffle, M.; Sebastian, P.; Norra, S.; Petney, T.; Oehme, R.; Littwin, N.; Lebl, K.; Raith, J.; et al. Estimating Ixodes ricinus densities on the landscape scale. Int. J. Health Geogr. 2015, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Schulz, M.; Mahling, M.; Pfister, K. Abundance and seasonal activity of questing Ixodes Ricinus ticks in their natural habitats in southern Germany in 2011. J. Vector Ecol. 2014, 39, 56–65. [Google Scholar] [CrossRef]
- Dautel, H.; Knülle, W. Cold hardiness, supercooling ability and causes of low-temperature mortality in the soft tick, Argas reflexus, and the hard tick, Ixodes ricinus (Acari: Ixodoidea) from Central Europe. J. Insect Physiol. 1997, 43, 843–854. [Google Scholar] [CrossRef]
- Rieck, T.; Steffen, A.; Schmid-Küpke, N.; Feig, M.; Wichmann, O.; Siedler, A. Impfquoten bei Erwachsenen in Deutschland—Aktuelles aus der KV-Impfsurveillance und der Onlinebefragung von Krankenhauspersonal OKaPII. Epidemiol. Bull. 2020, 47, 3–26. [Google Scholar] [CrossRef]
- Bundesamt für Kartographie und Geodäsie. Verwaltungsgebiete 1:250,000 Aktualisiert, Stand 2019. Available online: https://www.bkg.bund.de/SharedDocs/Produktinformationen/BKG/DE/P-2021/210127_VG250.html (accessed on 23 November 2021).
- Bundesamt für Kartographie und Geodäsie. Digitales Geländemodell Gitterweite 200 m (DGM200). Available online: https://gdz.bkg.bund.de/index.php/default/digitales-gelandemodell-gitterweite-200-m-dgm200.html (accessed on 20 July 2021).
- Bundesamt für Kartographie und Geodäsie. Digitales Landbedeckungsmodell für Deutschland (LBM-DE2018). Available online: https://gdz.bkg.bund.de/index.php/default/digitales-landbedeckungsmodell-fur-deutschland-stand-2018-lbm-de2018.html (accessed on 10 July 2021).
- Deutscher Wetterdienst. Open Data Bereich des Climate Data Center. Available online: https://www.dwd.de/DE/leistungen/cdc/climate-data-center.html;jsessionid=9E22FEE9BE12F436B7A14F110C7FB81D.live11042?nn=17626 (accessed on 30 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 10 July 2021).
- Hair, J.F.J.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Multivariate Data Analysis, 3rd ed.; Macmillan: New York, NY, USA, 1995. [Google Scholar]
- Zajic, A. Introduction to AIC—Akaike Information Criterion. Available online: https://towardsdatascience.com/introduction-to-aic-akaike-information-criterion-9c9ba1c96ced (accessed on 23 September 2021).
- Melo, F. Area under the ROC Curve. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.-H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013; pp. 38–39. [Google Scholar]
- Google Developers. Machine Learning Crash Course: Classification: ROC Curve and AUC. Available online: https://developers.google.com/machine-learning/crash-course/classification/roc-and-auc (accessed on 13 September 2021).
- RDocumentation. Predict: Model Prediction. Available online: https://www.rdocumentation.org/packages/car/versions/3.0-12/topics/Predict (accessed on 22 February 2022).
- Walter, M.; Vogelgesang, J.R.; Rubel, F.; Brugger, K. Tick-Borne Encephalitis Virus and Its European Distribution in Ticks and Endothermic Mammals. Microorganisms 2020, 8, 1065. [Google Scholar] [CrossRef] [PubMed]
- Jore, S.; Vanwambeke, S.O.; Viljugrein, H.; Isaksen, K.; Kristoffersen, A.B.; Woldehiwet, Z.; Johansen, B.; Brun, E.; Brun-Hansen, H.; Westermann, S.; et al. Climate and environmental change drives Ixodes ricinus geographical expansion at the northern range margin. Parasites Vectors 2014, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Place Indicated | Study Participants (n = 359) |
| Indicated points only | 163 (45%) |
| Indicated polygons only | 128 (36%) |
| Indicated both | 68 (19%) |
| Number of Places Indicated | Study Participants (n = 359) |
| Indicated 1 place | 235 (66%) |
| Indicated 2–5 places | 119 (33%) |
| Indicated 6 or more places | 5 (1%) |
| Tick Bite yes or no | Study Participants (n = 359) |
| None | 120 (33%) |
| One or more | 239 (67%) |
| Buffered Points (n = 320) | Polygons (n = 247) | TBE Foci (n = 41) | Comparator Points (n = 960) | Comparator Polygons (n = 741) | |
|---|---|---|---|---|---|
| Min. | 0.28 | 0.09 | <0.01 | 0.28 | 2.30 |
| Max. | 0.50 | 5.34 | 0.09 | 0.50 | 2.30 |
| Median | 0.50 | 1.93 | 0.01 | 0.50 | 2.30 |
| Mean | 0.44 | 2.29 | 0.03 | 0.43 | 2.30 |
| Standard Deviation | 0.07 | 1.45 | 0.02 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedsam, A.M.; Brady, O.J.; Pilic, A.; Dobler, G.; Hellenbrand, W.; Nygren, T.M. Geo-Spatial Characteristics of 567 Places of Tick-Borne Encephalitis Infection in Southern Germany, 2018–2020. Microorganisms 2022, 10, 643. https://doi.org/10.3390/microorganisms10030643
Friedsam AM, Brady OJ, Pilic A, Dobler G, Hellenbrand W, Nygren TM. Geo-Spatial Characteristics of 567 Places of Tick-Borne Encephalitis Infection in Southern Germany, 2018–2020. Microorganisms. 2022; 10(3):643. https://doi.org/10.3390/microorganisms10030643
Chicago/Turabian StyleFriedsam, Amelie M., Oliver J. Brady, Antonia Pilic, Gerhard Dobler, Wiebke Hellenbrand, and Teresa M. Nygren. 2022. "Geo-Spatial Characteristics of 567 Places of Tick-Borne Encephalitis Infection in Southern Germany, 2018–2020" Microorganisms 10, no. 3: 643. https://doi.org/10.3390/microorganisms10030643
APA StyleFriedsam, A. M., Brady, O. J., Pilic, A., Dobler, G., Hellenbrand, W., & Nygren, T. M. (2022). Geo-Spatial Characteristics of 567 Places of Tick-Borne Encephalitis Infection in Southern Germany, 2018–2020. Microorganisms, 10(3), 643. https://doi.org/10.3390/microorganisms10030643