
Interaction and Assembly of Bacterial Communities in High-Latitude Coral Habitat Associated Seawater


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
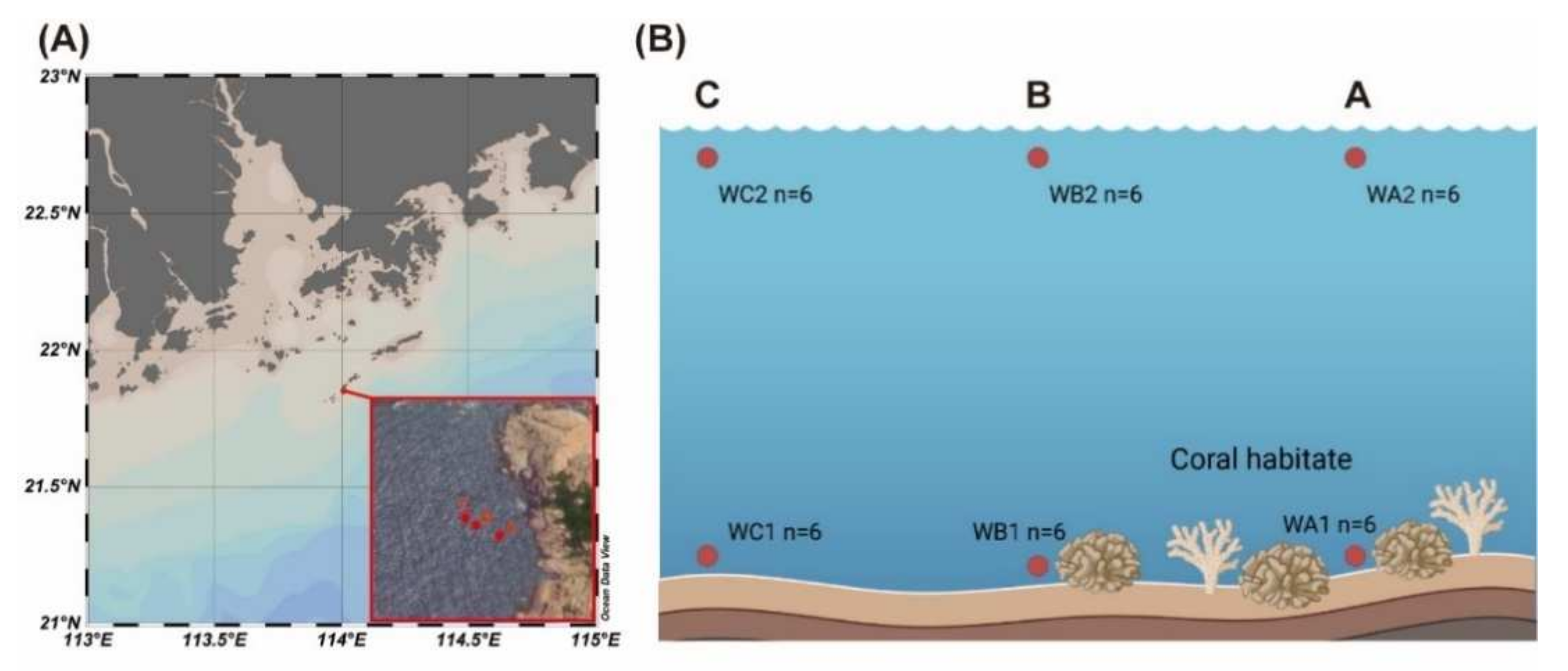
2.1. Sample Collection
2.2. DNA Extraction, Amplification, and Sequence Analysis
2.3. Statistical Analysis
3. Results
3.1. Alpha Diversity of Bacteria and Carbon-Fixation Bacteria
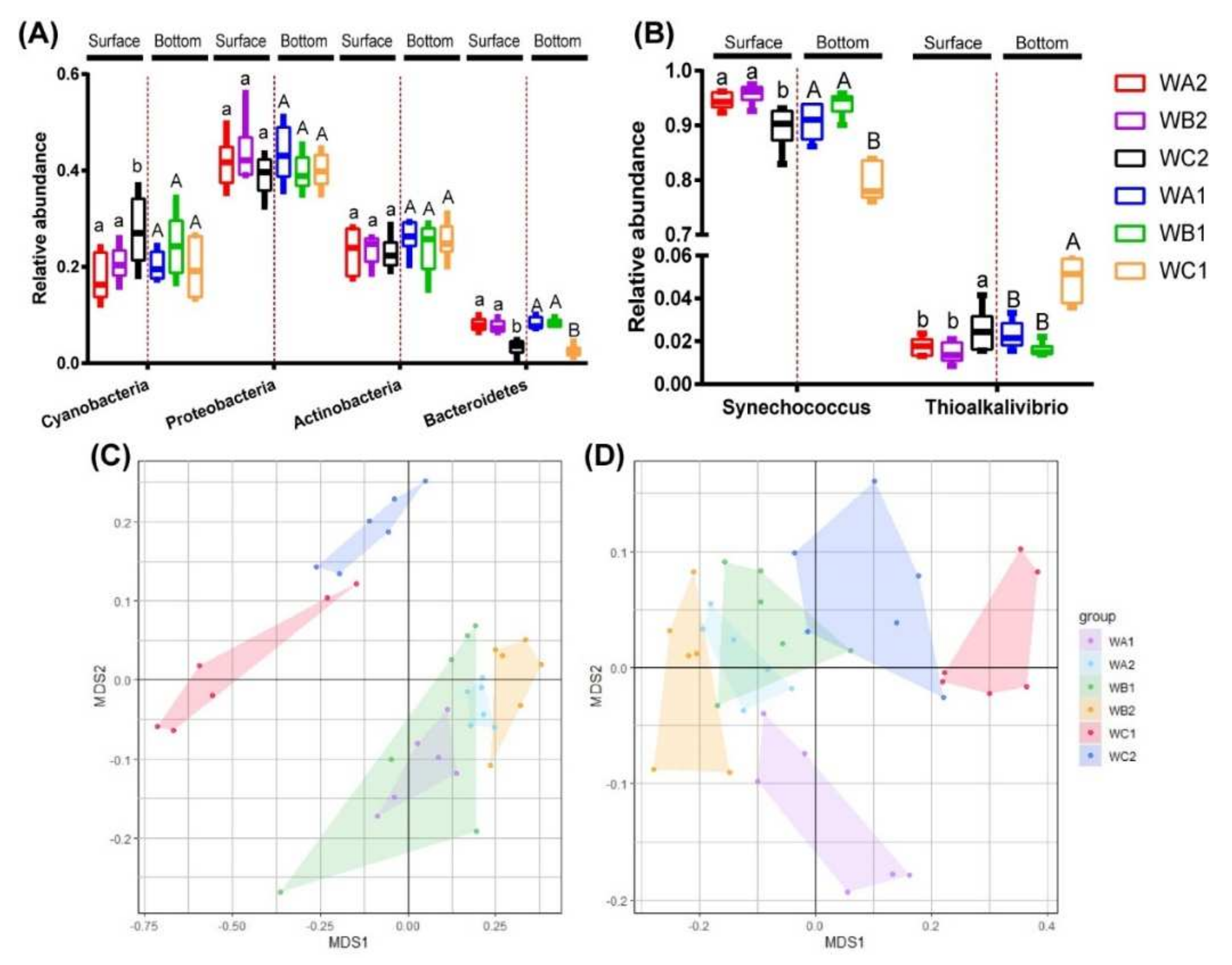
3.2. Composition of Bacterial and Carbon-Fixation Bacterial Community
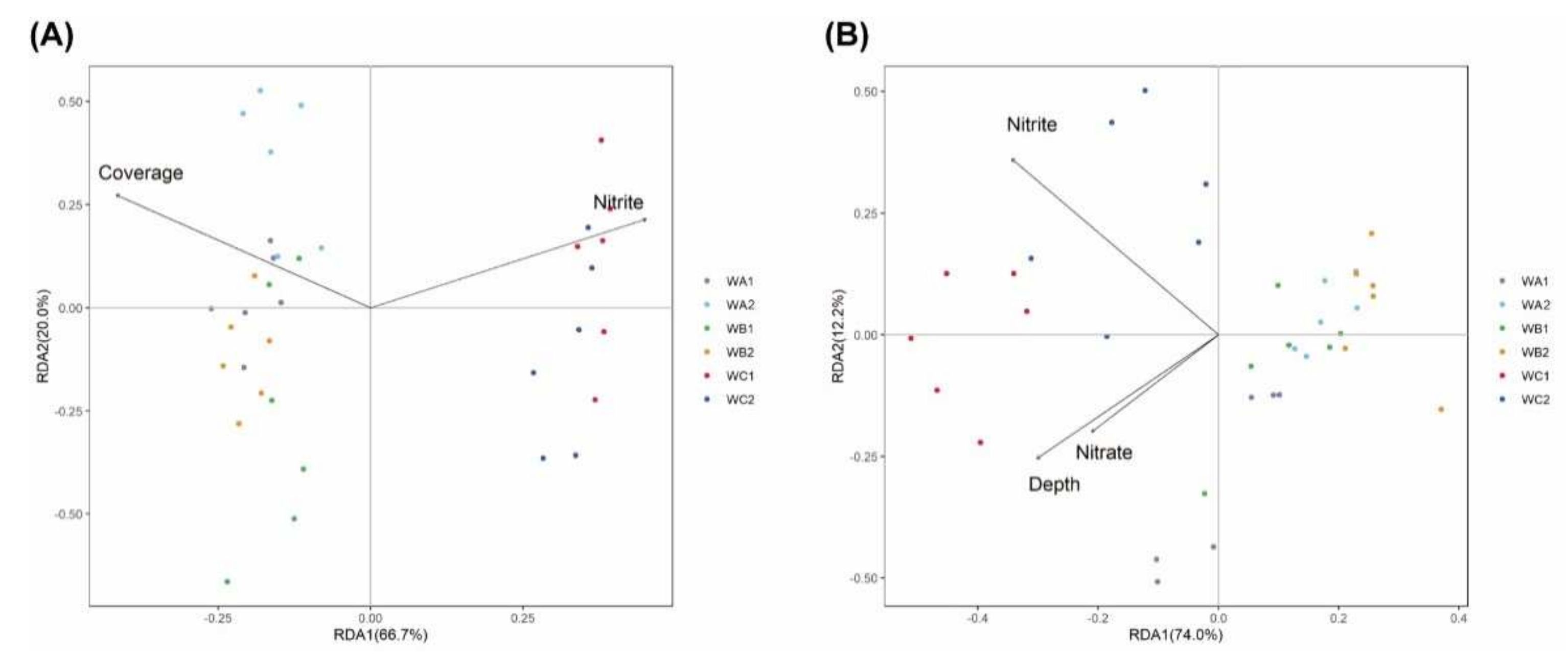
3.3. Relationship between Microbial Communities and Environment
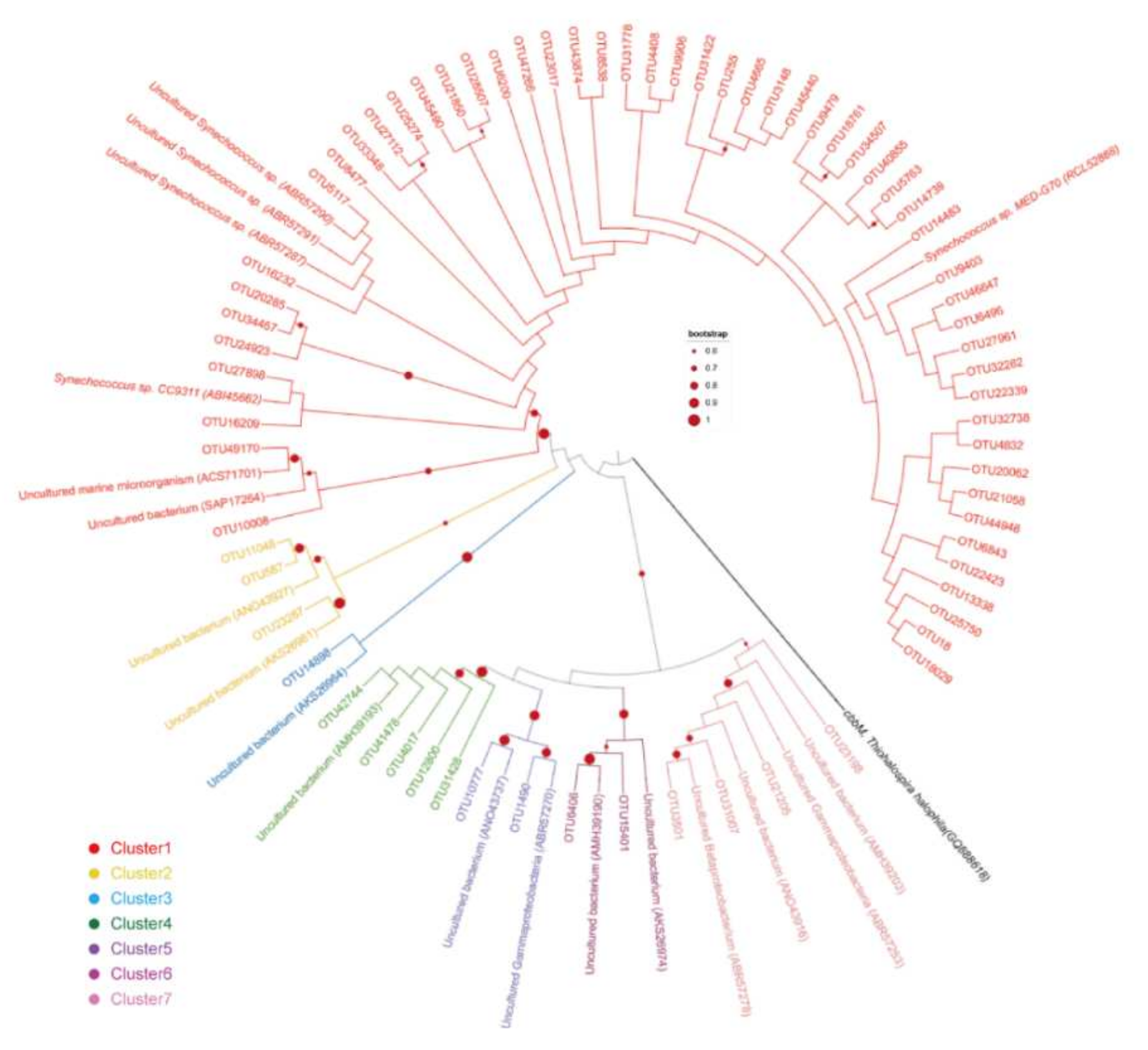
3.4. Phylogenetic Diversity of Carbon-Fixation Bacteria
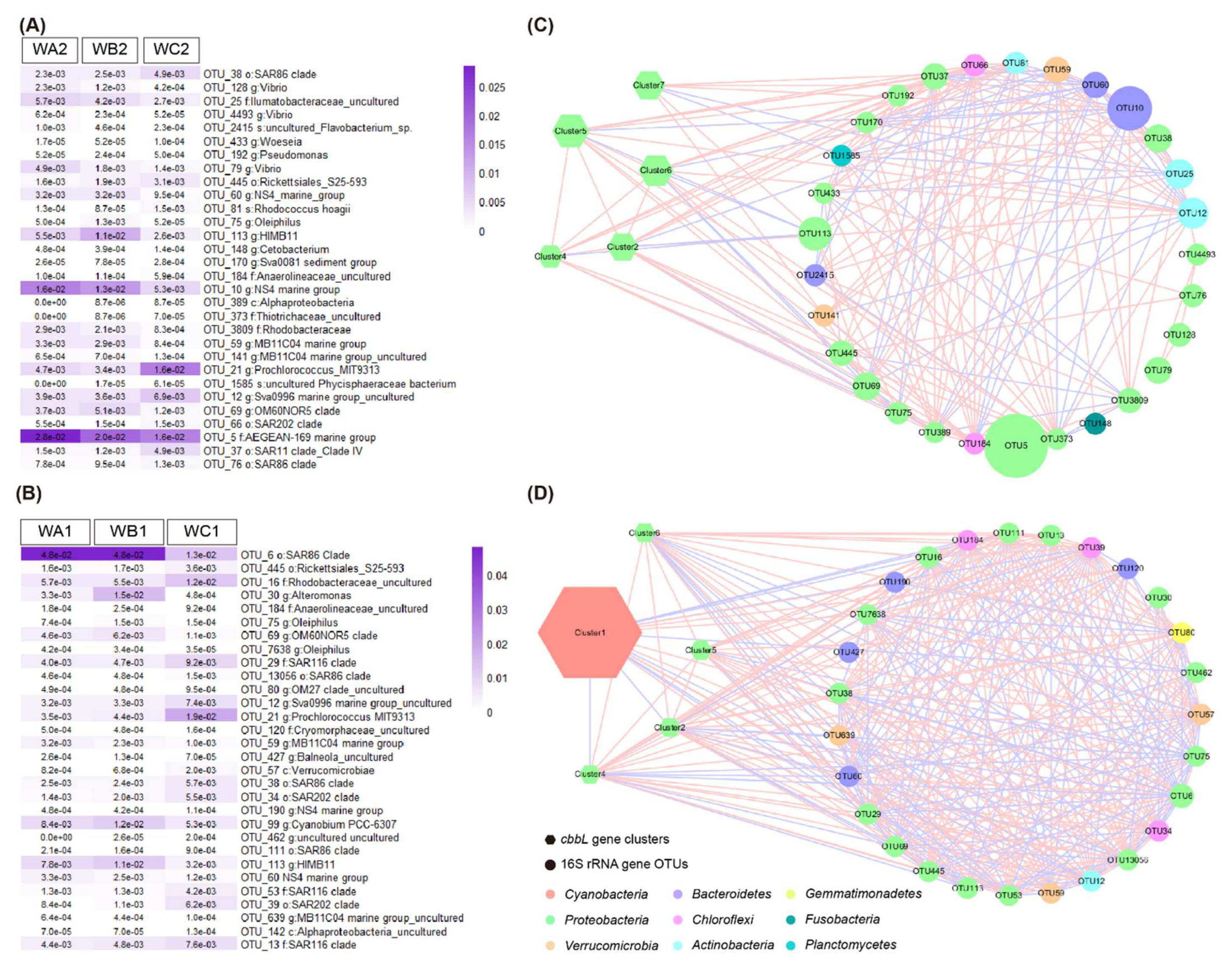
3.5. Interaction of Key Bacteria and Carbon-Fixers
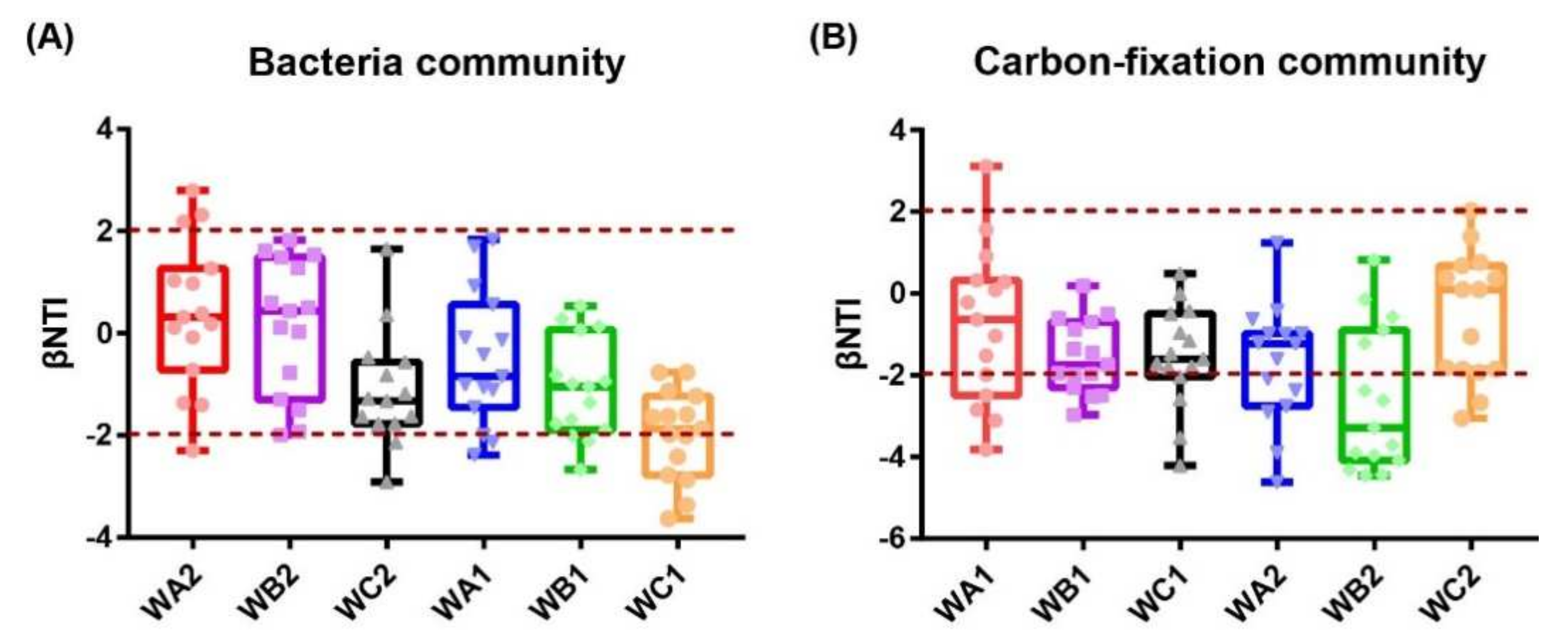
3.6. Assembly of Bacterial and Carbon-Fixation Bacterial Communities
4. Discussion
4.1. Environmental Drivers of the Bacterial Distribution in Coral-Associated Water Column
4.2. Interactions between Heterotrophic Bacteria and Carbon-Fixation Bacteria
4.3. Assembly of Environmental Bacterial and Carbon-Fixation Bacterial Community in Seawater of High-Latitude Coral Habitat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.M.; McClean, C.J.; Veron, J.E.; Hawkins, J.P.; Allen, G.R.; McAllister, D.E.; Mittermeier, C.G.; Schueler, F.W.; Spalding, M.; Wells, F.; et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 2002, 295, 1280–1284. [Google Scholar] [CrossRef] [Green Version]
- Van Hoytema, N.; Bednarz, V.N.; Cardini, U.; Naumann, M.S.; Al-Horani, F.A.; Wild, C. The influence of seasonality on benthic primary production in a red sea coral reef. Mar. Biol. 2016, 163, 52. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Carlson, C.A.; Carpenter, R.C. Sources of organic carbon to coral reef flats. Oceanography 2013, 26, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Cardini, U.; Bednarz, V.N.; van Hoytema, N.; Rovere, A.; Naumann, M.S.; Al-Rshaidat, M.; Wild, C. Budget of primary production and dinitrogen fixation in a highly seasonal red sea coral reef. Ecosystems 2016, 19, 771–785. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, B.G. Coral reef primary productivity: A beggar’s banquet. Trends Ecol. Evol. 1988, 3, 106–111. [Google Scholar] [CrossRef]
- Silveira, C.B.; Cavalcanti, G.S.; Walter, J.M.; Silva-Lima, A.W.; Dinsdale, E.A.; Bourne, D.G.; Thompson, C.C.; Thompson, F.L. Microbial processes driving coral reef organic carbon flow. FEMS Microbiol. Rev. 2017, 41, 575–595. [Google Scholar] [CrossRef] [Green Version]
- Garren, M.; Azam, F. New directions in coral reef microbial ecology. Environ. Microbiol. 2012, 14, 833–844. [Google Scholar] [CrossRef]
- Kegler, H.F.; Lukman, M.; Teichberg, M.; Plass-Johnson, J.; Hassenrück, C.; Wild, C.; Gärdes, A. Bacterial community composition and potential driving factors in different reef habitats of the spermonde archipelago, indonesia. Front. Microbiol. 2017, 8, 662. [Google Scholar] [CrossRef] [Green Version]
- Voolstra, C.R.; Berumen, M.L. Coral Reefs of the Red Sea; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Furby, K.A.; Apprill, A.; Cervino, J.M.; Ossolinski, J.E.; Hughen, K.A. Incidence of lesions on fungiidae corals in the eastern red sea is related to water temperature and coastal pollution. Mar. Environ. Res. 2014, 98, 29–38. [Google Scholar] [CrossRef]
- Locarnini, M.; Mishonov, A.; Baranova, O.; Boyer, T.; Zweng, M.; Garcia, H.; Seidov, D.; Weathers, K.; Paver, C.; Smolyar, I. World Ocean Atlas 2018. Volume 1: Temperature; U.S. Department of Commerce: Washington, DC, USA, 2018. [Google Scholar]
- Hu, J.; Wang, L.; Zhang, S.; Xi, X.; Le, Y.; Fu, X.; Tsang, Y.; Gao, M. Interactions between autotrophic and heterotrophic strains improve CO2 fixing efficiency of non-photosynthetic microbial communities. Appl. Biochem. Biotechnol. 2015, 176, 1459–1471. [Google Scholar] [CrossRef]
- Wang, J.; Shen, J.I.; Wu, Y.; Tu, C.; Soininen, J.; Stegen, J.C.; He, J.; Liu, X.; Zhang, L.; Zhang, E. Phylogenetic beta diversity in bacterial assemblages across ecosystems: Deterministic versus stochastic processes. ISME J. 2013, 7, 1310–1321. [Google Scholar] [CrossRef]
- Glasl, B.; Bourne, D.G.; Frade, P.R.; Thomas, T.; Schaffelke, B.; Webster, N.S. Microbial indicators of environmental perturbations in coral reef ecosystems. Microbiome 2019, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Schoettner, S.; Wild, C.; Hoffmann, F.; Boetius, A.; Ramette, A. Spatial scales of bacterial diversity in cold-water coral reef ecosystems. PLoS ONE 2012, 7, e32093. [Google Scholar] [CrossRef] [Green Version]
- Kinsey, D.; Hopley, D. The significance of coral reefs as global carbon sinks—Response to greenhouse. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1991, 89, 363–377. [Google Scholar] [CrossRef]
- Kayanne, H.; Suzuki, A.; Saito, H. Diurnal changes in the partial pressure of carbon dioxide in coral reef water. Science 1995, 269, 214–216. [Google Scholar] [CrossRef]
- Chisholm, J.R.; Barnes, D.J. Anomalies in coral reef community metabolism and their potential importance in the reef CO2 source-sink debate. Proc. Natl. Acad. Sci. USA 1998, 95, 6566–6569. [Google Scholar] [CrossRef] [Green Version]
- Precht, W.F.; Aronson, R.B. Climate flickers and range shifts of reef corals. Front. Ecol. Environ. 2004, 2, 307–314. [Google Scholar] [CrossRef]
- Greenstein, B.J.; Pandolfi, J.M. Escaping the heat: Range shifts of reef coral taxa in coastal western Australia. Glob. Change Biol. 2008, 14, 513–528. [Google Scholar] [CrossRef]
- Yamano, H.; Sugihara, K.; Nomura, K. Rapid poleward range expansion of tropical reef corals in response to rising sea surface temperatures. Geophys. Res. Lett. 2011, 38, L04601. [Google Scholar] [CrossRef]
- Beger, M.; Sommer, B.; Harrison, P.L.; Smith, S.D.; Pandolfi, J.M. Conserving potential coral reef refuges at high latitudes. Div. Distrib. 2014, 20, 245–257. [Google Scholar] [CrossRef]
- Li, J.; Long, L.; Zou, Y.; Zhang, S. Microbial community and transcriptional responses to increased temperatures in coral pocillopora damicornis holobiont. Environ. Microbiol. 2021, 23, 826–843. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, P.; Rui, J.; Cheng, L.; Wang, Q.; Liu, X.; Yuan, Q. Dark carbon fixation and chemolithotrophic microbial community in surface sediments of the cascade reservoirs, southwest china. Sci. Total Environ. 2020, 698, 134316. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than blast. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Uparse: Highly accurate otu sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. Vsearch: A versatile open source tool for metagenomics. Peerj 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Commun. Ecol. Package 2013, 2, 1–295. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Liaw, A.; Wiener, M. Classification and regression by randomforest. R News 2002, 2, 18–22. [Google Scholar]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Bruce, T.; Meirelles, P.M.; Garcia, G.; Paranhos, R.; Rezende, C.E.; de Moura, R.L.; Coni, E.O.; Vasconcelos, A.T.; Amado Filho, G.; Hatay, M. Abrolhos bank reef health evaluated by means of water quality, microbial diversity, benthic cover, and fish biomass data. PLoS ONE 2012, 7, e36687. [Google Scholar] [CrossRef]
- Frade, P.R.; Glasl, B.; Matthews, S.A.; Mellin, C.; Serrão, E.A.; Wolfe, K.; Mumby, P.J.; Webster, N.S.; Bourne, D.G. Spatial patterns of microbial communities across surface waters of the great barrier reef. Commun. Biol. 2020, 3, 442. [Google Scholar] [CrossRef]
- Luo, H.; Moran, M.A. How do divergent ecological strategies emerge among marine bacterioplankton lineages? Trends Microbiol. 2015, 23, 577–584. [Google Scholar] [CrossRef]
- Laas, P.; Ugarelli, K.; Absten, M.; Boyer, B.; Briceño, H.; Stingl, U. Composition of prokaryotic and eukaryotic microbial communities in waters around the florida reef tract. Microorganisms 2021, 9, 1120. [Google Scholar] [CrossRef]
- Neave, M.J.; Apprill, A.; Aeby, G.; Miyake, S.; Voolstra, C.R. Microbial communities of red sea coral reefs. Coral Reefs Red Sea 2019, 4, 53–68. [Google Scholar]
- Kelly, L.W.; Williams, G.J.; Barott, K.L.; Carlson, C.A.; Dinsdale, E.A.; Edwards, R.A.; Haas, A.F.; Haynes, M.; Lim, Y.W.; McDole, T. Local genomic adaptation of coral reef-associated microbiomes to gradients of natural variability and anthropogenic stressors. Proc. Natl. Acad. Sci. USA 2014, 111, 10227–10232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.E.; Alldredge, A.L.; McCliment, E.A.; Amaral-Zettler, L.A.; Carlson, C.A. Depleted dissolved organic carbon and distinct bacterial communities in the water column of a rapid-flushing coral reef ecosystem. ISME J. 2011, 5, 1374–1387. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.; González-Díaz, P.; Armenteros, M.; Ferrer, V.M.; Bretos, F.; Bartels, E.; Santoro, A.E.; Apprill, A. Microbial signatures of protected and impacted northern caribbean reefs: Changes from cuba to the florida keys. Environ. Microbiol. 2020, 22, 499–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Ding, Z.; Li, W.; Chen, X.; Yu, Y.; Zhao, X.; Lian, X.; Wang, Y. The effect of seawater environmental factors on the corals of wailingding island in the pearl river estuary. Cont. Shelf Res. 2020, 197, 104087. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Langlois, R.; Milne, A.; Achterberg, E.P.; La Roche, J.; Geider, R.J. Relative influence of nitrogen and phosphorous availability on phytoplankton physiology and productivity in the oligotrophic sub-tropical north Atlantic Ocean. Limnol. Oceanogr. 2008, 53, 291–305. [Google Scholar] [CrossRef]
- Moore, C.; Mills, M.; Arrigo, K.; Berman-Frank, I.; Bopp, L.; Boyd, P.; Galbraith, E.; Geider, R.; Guieu, C.; Jaccard, S. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Coles, S.L.; Strathmann, R. Observations on coral mucus “flocs” and their potential trophic significance 1. Limnol. Oceanogr. 1973, 18, 673–678. [Google Scholar] [CrossRef]
- Kurihara, H.; Ikeda, N.; Umezawa, Y. Diurnal and seasonal variation of particle and dissolved organic matter release by the coral acropora tenuis. Peerj 2018, 6, e5728. [Google Scholar] [CrossRef]
- Moynihan, M.A.; Goodkin, N.F.; Morgan, K.M.; Kho, P.Y.; Lopes dos Santos, A.; Lauro, F.M.; Baker, D.M.; Martin, P. Coral-associated nitrogen fixation rates and diazotrophic diversity on a nutrient-replete equatorial reef. ISME J. 2022, 16, 233–246. [Google Scholar] [CrossRef]
- Lesser, M.P.; Morrow, K.M.; Pankey, S.M.; Noonan, S.H. Diazotroph diversity and nitrogen fixation in the coral stylophora pistillata from the great barrier reef. ISME J. 2018, 12, 813–824. [Google Scholar] [CrossRef]
- El-Khaled, Y.C.; Roth, F.; Rädecker, N.; Tilstra, A.; Karcher, D.B.; Kürten, B.; Jones, B.H.; Voolstra, C.R.; Wild, C. Nitrogen fixation and denitrification activity differ between coral- and algae-dominated red sea reefs. Sci. Rep. 2021, 11, 11820. [Google Scholar] [CrossRef]
- Buchan, A.; LeCleir, G.R.; Gulvik, C.A.; González, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef]
- Zheng, Q.; Wang, Y.; Lu, J.; Lin, W.; Chen, F.; Jiao, N. Metagenomic and metaproteomic insights into photoautotrophic and heterotrophic interactions in a synechococcus culture. MBio 2020, 11, e03261-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, S.A.; Green, D.H.; Hart, M.C.; Küpper, F.C.; Sunda, W.G.; Carrano, C.J. Photolysis of iron–siderophore chelates promotes bacterial–algal mutualism. Proc. Natl. Acad. Sci. USA 2009, 106, 17071–17076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazamia, E.; Czesnick, H.; Nguyen, T.T.V.; Croft, M.T.; Sherwood, E.; Sasso, S.; Hodson, S.J.; Warren, M.J.; Smith, A.G. Mutualistic interactions between vitamin b12-dependent algae and heterotrophic bacteria exhibit regulation. Environ. Microbiol. 2012, 14, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.J.; Kirkegaard, R.; Szul, M.J.; Johnson, Z.I.; Zinser, E.R. Facilitation of robust growth of prochlorococcus colonies and dilute liquid cultures by “helper” heterotrophic bacteria. Appl. Environ. Microbiol. 2008, 74, 4530–4534. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.J.; Johnson, Z.I.; Szul, M.J.; Keller, M.; Zinser, E.R. Dependence of the cyanobacterium prochlorococcus on hydrogen peroxide scavenging microbes for growth at the ocean’s surface. PLoS ONE 2011, 6, e16805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labban, A.; Palacio, A.S.; García, F.C.; Hadaidi, G.; Ansari, M.I.; López-Urrutia, Á.; Alonso-Sáez, L.; Hong, P.-Y.; Morán, X.A.G. Temperature responses of heterotrophic bacteria in co-culture with a red sea synechococcus strain. Front. Microbiol. 2021, 12, 612732. [Google Scholar] [CrossRef]
- Arandia-Gorostidi, N.; Weber, P.K.; Alonso-Sáez, L.; Morán, X.A.G.; Mayali, X. Elevated temperature increases carbon and nitrogen fluxes between phytoplankton and heterotrophic bacteria through physical attachment. ISME J. 2017, 11, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Hornick, T.; Bach, L.T.; Crawfurd, K.J.; Spilling, K.; Achterberg, E.; Brussaard, C.; Riebesell, U.; Grossart, H. Ocean acidification indirectly alters trophic interaction of heterotrophic bacteria at low nutrient conditions. Biogeosci. Discuss. 2016, 10, 61. [Google Scholar]
- Zhang, R.; Jiang, T.; Tian, Y.; Xie, S.; Zhou, L.; Li, Q.; Jiao, N. Volcanic ash stimulates growth of marine autotrophic and heterotrophic microorganisms. Geology 2017, 45, 679–682. [Google Scholar] [CrossRef]
- Hoarfrost, A.; Nayfach, S.; Ladau, J.; Yooseph, S.; Arnosti, C.; Dupont, C.L.; Pollard, K.S. Global ecotypes in the ubiquitous marine clade sar86. ISME J. 2020, 14, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrshad, M.; Rodriguez-Valera, F.; Amoozegar, M.A.; López-García, P.; Ghai, R. The enigmatic sar202 cluster up close: Shedding light on a globally distributed dark ocean lineage involved in sulfur cycling. ISME J. 2018, 12, 655–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, S.J. Sar11 bacteria: The most abundant plankton in the oceans. Ann. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Pajares, S.; Varona-Cordero, F.; Hernández-Becerril, D.U. Spatial distribution patterns of bacterioplankton in the oxygen minimum zone of the tropical mexican pacific. Microb. Ecol. 2020, 80, 519–536. [Google Scholar] [CrossRef]
- Zheng, Q.; Wang, Y.; Xie, R.; Lang, A.S.; Liu, Y.; Lu, J.; Zhang, X.; Sun, J.; Suttle, C.A.; Jiao, N. Dynamics of heterotrophic bacterial assemblages within synechococcus cultures. Appl. Environ. Microbiol. 2018, 84, e01517. [Google Scholar] [CrossRef] [Green Version]
- Cruz, B.N.; Neuer, S. Heterotrophic bacteria enhance the aggregation of the marine picocyanobacteria prochlorococcus and synechococcus. Front. Microbiol. 2019, 10, 1864. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Nakamura, T. Carbon dynamics in coral reefs. In Blue Carbon in Shallow Coastal Ecosystems; Springer: Singapore, 2019; pp. 273–293. [Google Scholar]
- Stegen, J.C.; Lin, X.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Pearman, J.K.; Aylagas, E.; Voolstra, C.R.; Anlauf, H.; Villalobos, R.; Carvalho, S. Disentangling the complex microbial community of coral reefs using standardized autonomous reef monitoring structures (arms). Mol. Ecol. 2019, 28, 3496–3507. [Google Scholar] [CrossRef] [Green Version]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Li, J.; Lyu, Y.; Zou, Y.; Li, Q.; Yang, Q.; Tang, X.; Yuan, X.; Jiang, Z.; Zhang, S. Interaction and Assembly of Bacterial Communities in High-Latitude Coral Habitat Associated Seawater. Microorganisms 2022, 10, 558. https://doi.org/10.3390/microorganisms10030558
Chen Y, Li J, Lyu Y, Zou Y, Li Q, Yang Q, Tang X, Yuan X, Jiang Z, Zhang S. Interaction and Assembly of Bacterial Communities in High-Latitude Coral Habitat Associated Seawater. Microorganisms. 2022; 10(3):558. https://doi.org/10.3390/microorganisms10030558
Chicago/Turabian StyleChen, Yu, Jie Li, Yuanjiao Lyu, Yiyang Zou, Qiqi Li, Qingsong Yang, Xiaoyu Tang, Xiangcheng Yuan, Zhijian Jiang, and Si Zhang. 2022. "Interaction and Assembly of Bacterial Communities in High-Latitude Coral Habitat Associated Seawater" Microorganisms 10, no. 3: 558. https://doi.org/10.3390/microorganisms10030558
APA StyleChen, Y., Li, J., Lyu, Y., Zou, Y., Li, Q., Yang, Q., Tang, X., Yuan, X., Jiang, Z., & Zhang, S. (2022). Interaction and Assembly of Bacterial Communities in High-Latitude Coral Habitat Associated Seawater. Microorganisms, 10(3), 558. https://doi.org/10.3390/microorganisms10030558