
Dietary Fiber Intake and Gut Microbiota in Human Health


Abstract
1. Introduction
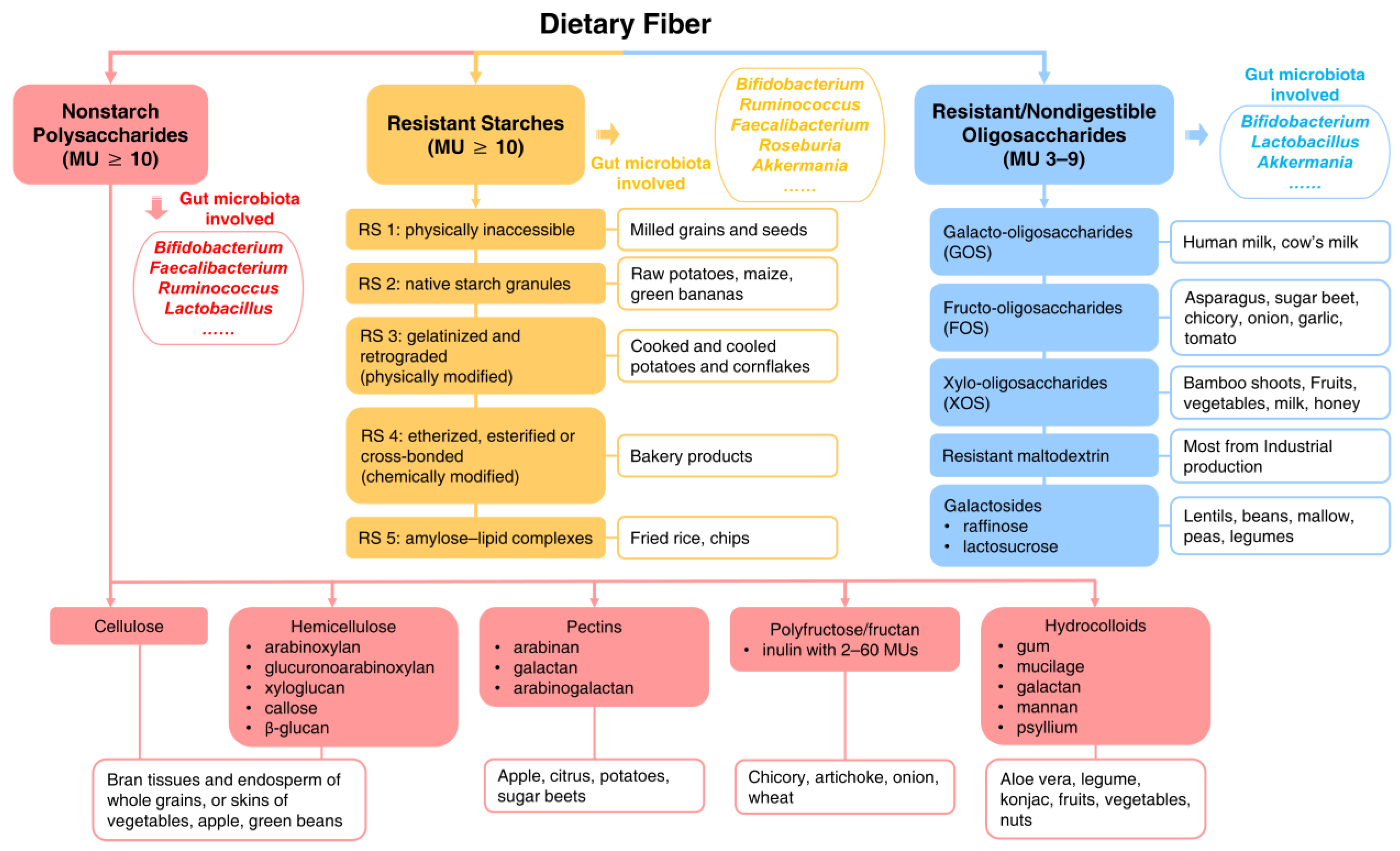
2. Dietary Fiber and Its Main Types
3. Average Levels and Recommended Amounts of Dietary Fiber Intake
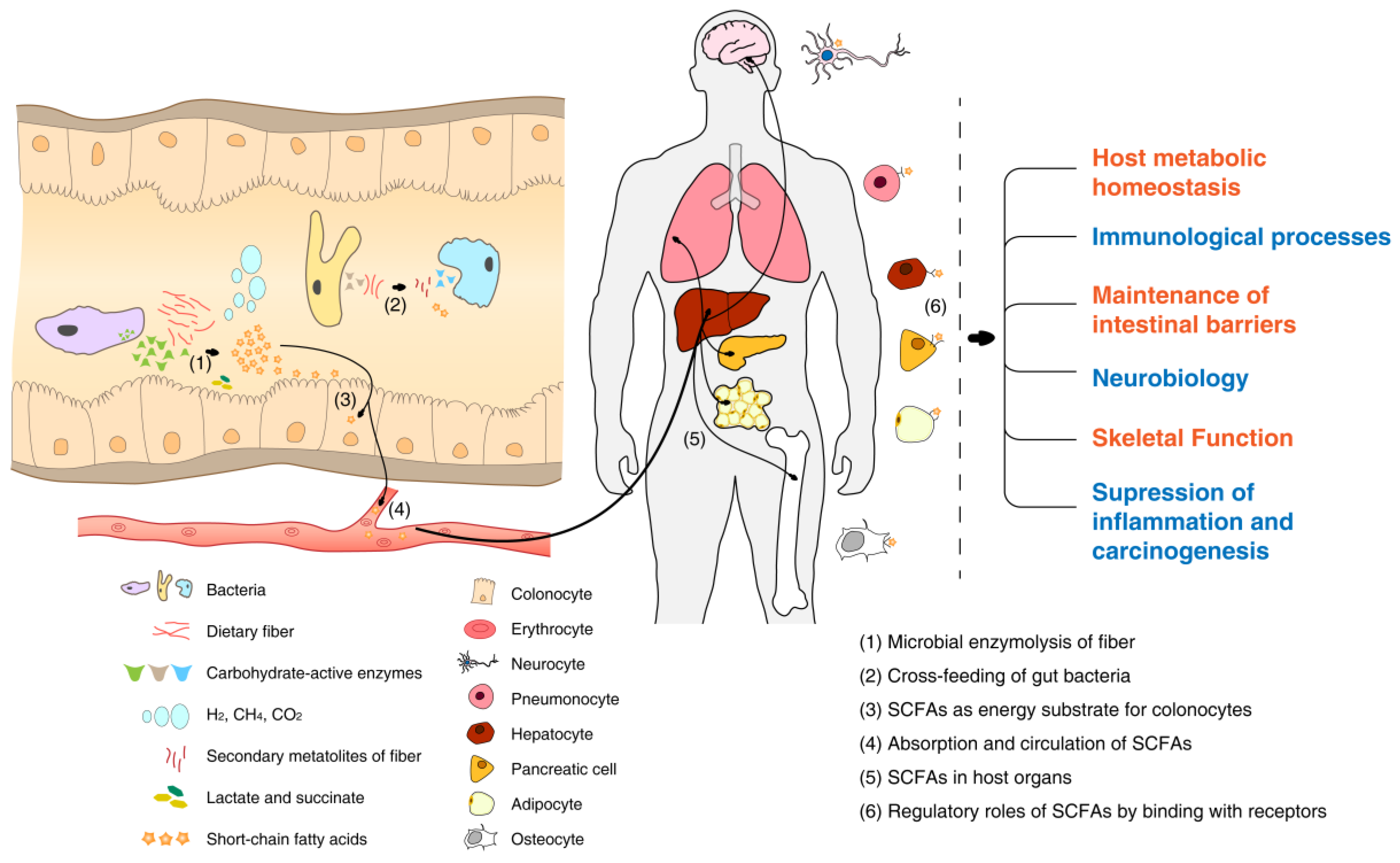
4. Fermentation of Dietary Fiber by the Gut Microbiota
5. Impacts of Dietary Fiber on Gut Microbial Community Structure and Diversity
6. Influences of Dietary Fiber on Different Gut Microbes
6.1. Bifidobacterium
6.2. Faecalibacterium
6.3. Ruminococcus
6.4. Lactobacillus
6.5. Prevotella
7. Cutoff Threshold of Dietary Fiber Intake on Gut Microbiota
8. Modulation of Dietary Fiber on Gut Microbiota in Diabetes Patients
9. Conclusions and Future Prospects
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrews, P.; Johnson, R.J. Evolutionary basis for the human diet: Consequences for human health. J. Intern. Med. 2020, 287, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Eaton, S.B.; Eaton, S.B., 3rd; Konner, M.J. Paleolithic nutrition revisited: A twelve-year retrospective on its nature and implications. Eur. J. Clin. Nutr. 1997, 51, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R.; Jeyakumar, T.; Poinar, H.; Gros, P. Shifting Climates, Foods, and Diseases: The Human Microbiome through Evolution. BioEssays News Rev. Mol. Cell. Dev. Biol. 2019, 41, e1900034. [Google Scholar] [CrossRef] [PubMed]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, L.; Yang, L.; Chu, H. The critical role of gut microbiota in obesity. Front. Endocrinol. 2022, 13, 1025706. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef]
- Papadopoulos, P.D.; Tsigalou, C.; Valsamaki, P.N.; Konstantinidis, T.G.; Voidarou, C.; Bezirtzoglou, E. The Emerging Role of the Gut Microbiome in Cardiovascular Disease: Current Knowledge and Perspectives. Biomedicines 2022, 10, 948. [Google Scholar] [CrossRef]
- Ye, J.; Wu, Z.; Zhao, Y.; Zhang, S.; Liu, W.; Su, Y. Role of gut microbiota in the pathogenesis and treatment of diabetes mullites: Advanced research-based review. Front. Microbiol. 2022, 13, 1029890. [Google Scholar] [CrossRef]
- Joint FAO/WHO Food Standards Programme. Secretariat of the CODEX Alimentarius Commission: CODEX Alimentarius (CODEX) Guidelines on Nutrition Labeling CAC/GL 2-1985 as Last Amended 2010; FAO: Rome, Italy, 2010. [Google Scholar]
- de Menezes, E.W.; Giuntini, E.B.; Dan, M.C.T.; Sardá, F.A.H.; Lajolo, F.M. Codex dietary fibre definition–Justification for inclusion of carbohydrates from 3 to 9 degrees of polymerisation. Food Chem. 2013, 140, 581–585. [Google Scholar] [CrossRef]
- Dai, F.J.; Chau, C.F. Classification and regulatory perspectives of dietary fiber. J. Food Drug Anal. 2017, 25, 37–42. [Google Scholar] [CrossRef]
- Stephen, A.M.; Champ, M.M.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary fibre in Europe: Current state of knowledge on definitions, sources, recommendations, intakes and relationships to health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific Opinion on Dietary Reference Values for carbohydrates and dietary fibre. EFSA J. 2010, 8, 1462. [Google Scholar]
- Rezende, E.S.V.; Lima, G.C.; Naves, M.M.V. Dietary fibers as beneficial microbiota modulators: A proposed classification by prebiotic categories. Nutrition 2021, 89, 111217. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.; de Boeck, G.; Becker, K. Dietary roles of non-starch polysaccharides in human nutrition: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 899–935. [Google Scholar] [CrossRef] [PubMed]
- Tungland, B.C.; Meyer, D. Nondigestible Oligo- and Polysaccharides (Dietary Fiber): Their Physiology and Role in Human Health and Food. Compr. Rev. Food Sci. Food Saf. 2002, 1, 90–109. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Inulin-type fructans: Functional food ingredients. J. Nutr. 2007, 137 (Suppl. 11), 2493s–2502s. [Google Scholar] [CrossRef]
- Raigond, P.; Ezekiel, R.; Raigond, B. Resistant starch in food: A review. J. Sci. Food Agric. 2015, 95, 1968–1978. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Miller, K.B. Review of whole grain and dietary fiber recommendations and intake levels in different countries. Nutr. Rev. 2020, 78 (Suppl. 1), 29–36. [Google Scholar] [CrossRef]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the ‘fiber gap’. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef]
- Agnes, N.P.; Christensen, T.; Matthiessen, J.; Knudsen, V.K.; Rosenlund-Sørensen, M.; Biltoft-Jensen, A.; Hinsch, H.; Ygil, K.H.; Kørup, K.; Saxholt, E.; et al. Dietary Habits in Denmark 2011–2013: Main Results. 2015. Available online: https://www.food.dtu.dk/publikationer/ernaering-og-kostvaner/de_nationale_kostundersoegelser (accessed on 25 October 2022).
- Totland, T.H.; Melnæs, B.K.; Lundberg-Hallén, N.; Helland-Kigen, K.M.; Lund-Blix, N.A.; Myhre, J.B.; Johansen, A.M.W.; Løken, E.B.; Andersen, L.F. Norkost 3–En Landsomfattende Kostholdsundersøkelse Blant Menn og Kvinner i Norge i Alderen 18–70 år 2010–11. 2012. Available online: https://www.helsedirektoratet.no/search?searchquery=Norkost%203 (accessed on 25 October 2022).
- Max Rubner-Institut. Die Nationale Verzehrsstudie II Abschlussbericht Teil 2 (Diet-History-Interviews); Max Rubner-Institut: Karlsruhe, Germany, 2008. [Google Scholar]
- Australian Bureau of Statistics Australian Health Survey: Nutrition First Results-Foods and Nutrients. Available online: https://www.abs.gov.au/statistics/health/health-conditions-and-risks/australian-health-survey-nutrition-first-results-foods-and-nutrients/2011-12/Table%201%20Mean%20daily%20energy%20and%20nutrient%20intake.xls (accessed on 25 October 2022).
- Frølich, W.; Aman, P.; Tetens, I. Whole grain foods and health–A Scandinavian perspective. Food Nutr. Res. 2013, 57. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Wang, Z.H.; Zhang, J.G.; Du, W.W.; Su, C.; Zhang, J.; Zhai, F.Y.; Zhang, B. Trends in dietary fiber intake in Chinese aged 45 years and above, 1991–2011. Eur. J. Clin. Nutr. 2014, 68, 619–622. [Google Scholar] [CrossRef] [PubMed]
- National Health and Family Planning Commission. Chinese Dietary Reference Intakes— Part 1: Macronutrient. 2017. Available online: http://www.nhc.gov.cn/wjw/yingyang/201710/fdade20feb8144ba921b412944ffb779.shtml (accessed on 25 October 2022).
- Ministry of Health, Labour and Welfare. The National Health and Nutrition Survey in Japan, 2019. 2019. Available online: https://www.mhlw.go.jp/stf/seisakunitsuite/bunya/kenkou_iryou/kenkou/eiyou/r1-houkoku_00002.html (accessed on 25 October 2022).
- Ministry of Health, Labour and Welfare. Dietary Reference Intakes for Japanese, (2020 Edition). 2020. Available online: https://www.mhlw.go.jp/stf/newpage_08517.html (accessed on 25 October 2022).
- U.S. Department of Agriculture, Agricultural Research Service. Nutrient Intakes from Food and Beverages: Mean Amounts Consumed per Individual, by Gender and Age. 2022. Available online: https://www.ars.usda.gov/northeast-area/beltsville-md-bhnrc/beltsville-human-nutrition-research-center/food-surveys-research-group/docs/wweia-data-tables (accessed on 25 October 2022).
- U.S. Department of Agriculture; U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; 2020. Available online: https://www.dietaryguidelines.gov (accessed on 25 October 2022).
- Ahmed, M.; Praneet Ng, A.; L’Abbe, M.R. Nutrient intakes of Canadian adults: Results from the Canadian Community Health Survey (CCHS)-2015 Public Use Microdata File. Am. J. Clin. Nutr. 2021, 114, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Canada’s Dietary Guidelines: For Health Professionals and Policy Makers. 2019. Available online: https://food-guide.canada.ca/en/guidelines (accessed on 25 October 2022).
- National Health and Medical Research Council, Department of Health and Aged Care. Australian Dietary Guidelines-Providing the Scientific Evidence for Healthier Australian Diets. 2013. Available online: https://www.eatforhealth.gov.au/guidelines (accessed on 25 October 2022).
- Public Health England. NDNS: Results from Years 9 to 11 (Combined)–Statistical Summary. 2020. Available online: https://www.gov.uk/government/statistics/ndns-results-from-years-9-to-11-2016-to-2017-and-2018-to-2019/ndns-results-from-years-9-to-11-combined-statistical-summary (accessed on 25 October 2022).
- Public Health England. Government Dietary Recommendations. 2016. Available online: https://www.gov.uk/government/publications/the-eatwell-guide (accessed on 25 October 2022).
- INCA 3: Evolution des Habitudes et Modes de Consommation, de Nouveaux Enjeux en Matière de Sécurité Sanitaire et de Nutrition. Available online: https://www.anses.fr/fr/content/inca-3-evolution-des-habitudes-et-modes-de-consommation-de-nouveaux-enjeux-en-mati%C3%A8re-de (accessed on 25 October 2022).
- Agence Nationale de Sécurité Sanitaire. Actualisation des Repères du PNNS: Élaboration des Références Nutritionnelles. 2016. Available online: https://www.anses.fr/en/system/files/NUT2012SA0103Ra-2.pdf (accessed on 25 October 2022).
- Nordic Nutrition Recommendations 2012: Integrating Nutrition and Physical Activity, 5th ed.; Nordisk Ministerråd: Copenhagen, Denmark, 2014; p. 627.
- Ballaststoffe (Nahrungsfasern): Richtwerte für die Zufuhr. Available online: https://www.dge.de/wissenschaft/referenzwerte/ballaststoffe/ (accessed on 25 October 2022).
- Baturin, A.K.; Martinchik, A.N.; Kambarov, A.O. The transit of Russian nation nutrition at the turn of the 20th and 21st centuries. Vopr. Pitan. 2020, 89, 60–70. [Google Scholar]
- Federal Public Health Institution, Centre for Hygiene Education. Nutrients/Fiber. Available online: http://cgon.rospotrebnadzor.ru/content/sostav-pitaniya/pishevaya-kletchatka (accessed on 25 October 2022).
- Grasgruber, P.; Hrazdíra, E. Nutritional and socio-economic predictors of adult height in 152 world populations. Econ. Hum. Biol. 2020, 37, 100848. [Google Scholar] [CrossRef]
- Ihira, H.; Sawada, N.; Iwasaki, M.; Yamaji, T.; Goto, A.; Noda, M.; Iso, H.; Tsugane, S. Adult height and all-cause and cause-specific mortality in the Japan Public Health Center-based Prospective Study (JPHC). PLoS ONE 2018, 13, e0197164. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Sawada, N.; Tomata, Y.; Zhang, S.; Goto, A.; Yamaji, T.; Iwasaki, M.; Inoue, M.; Tsuji, I.; Tsugane, S. Association between adherence to the Japanese diet and all-cause and cause-specific mortality: The Japan Public Health Center-based Prospective Study. Eur. J. Nutr. 2021, 60, 1327–1336. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef]
- Tuncil, Y.E.; Thakkar, R.D.; Arioglu-Tuncil, S.; Hamaker, B.R.; Lindemann, S.R. Subtle Variations in Dietary-Fiber Fine Structure Differentially Influence the Composition and Metabolic Function of Gut Microbiota. mSphere 2020, 5, e00180-20. [Google Scholar] [CrossRef]
- Ndeh, D.; Gilbert, H.J. Biochemistry of complex glycan depolymerisation by the human gut microbiota. FEMS Microbiol. Rev. 2018, 42, 146–164. [Google Scholar] [CrossRef]
- Falony, G.; De Vuyst, L. Ecological Interactions of Bacteria in the Human Gut. In Prebiotics and Probiotics Science and Technology; Charalampopoulos, D., Rastall, R.A., Eds.; Springer: New York, NY, USA, 2009; pp. 639–679. [Google Scholar]
- Turroni, F.; Milani, C.; Duranti, S.; Mahony, J.; van Sinderen, D.; Ventura, M. Glycan Utilization and Cross-Feeding Activities by Bifidobacteria. Trends Microbiol. 2018, 26, 339–350. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Kumar, J.; Rani, K.; Datt, C. Molecular link between dietary fibre, gut microbiota and health. Mol. Biol. Rep. 2020, 47, 6229–6237. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Mandaliya, D.K.; Seshadri, S. Short Chain Fatty Acids, pancreatic dysfunction and type 2 diabetes. Pancreatol. Off. J. Int. Assoc. Pancreatol. (IAP) 2019, 19, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science (New York N.Y.) 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- De Filippo, C.; Di Paola, M.; Ramazzotti, M.; Albanese, D.; Pieraccini, G.; Banci, E.; Miglietta, F.; Cavalieri, D.; Lionetti, P. Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front. Microbiol. 2017, 8, 1979. [Google Scholar] [CrossRef]
- Merra, G.; Noce, A.; Marrone, G.; Cintoni, M.; Tarsitano, M.G.; Capacci, A.; De Lorenzo, A. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients 2020, 13, 19–25. [Google Scholar] [CrossRef]
- Ruengsomwong, S.; La-Ongkham, O.; Jiang, J.; Wannissorn, B.; Nakayama, J.; Nitisinprasert, S. Microbial Community of Healthy Thai Vegetarians and Non-Vegetarians, Their Core Gut Microbiota, and Pathogen Risk. J. Microbiol. Biotechnol. 2016, 26, 1723–1735. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Lu, M.; Cai, J.; Lu, B.; Luo, C.; Dai, M. Habitual Diet Pattern Associations with Gut Microbiome Diversity and Composition: Results from a Chinese Adult Cohort. Nutrients 2022, 14, 2639. [Google Scholar] [CrossRef]
- An, R.; Wilms, E.; Smolinska, A.; Hermes, G.D.A.; Masclee, A.A.M.; de Vos, P.; Schols, H.A.; van Schooten, F.J.; Smidt, H.; Jonkers, D.; et al. Sugar Beet Pectin Supplementation Did Not Alter Profiles of Fecal Microbiota and Exhaled Breath in Healthy Young Adults and Healthy Elderly. Nutrients 2019, 11, 2193. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual dietary fibre intake influences gut microbiota response to an inulin-type fructan prebiotic: A randomised, double-blind, placebo-controlled, cross-over, human intervention study. Br. J. Nutr. 2018, 119, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.T.; Brooks, S.P.J.; Inglis, G.D.; Yanke, L.J.; Green, J.; Petronella, N.; Ramdath, D.D.; Bercik, P.; Green-Johnson, J.M.; Kalmokoff, M. Impact of β2-1 fructan on faecal community change: Results from a placebo-controlled, randomised, double-blinded, cross-over study in healthy adults. Br. J. Nutr. 2017, 118, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Reider, S.J.; Moosmang, S.; Tragust, J.; Trgovec-Greif, L.; Tragust, S.; Perschy, L.; Przysiecki, N.; Sturm, S.; Tilg, H.; Stuppner, H.; et al. Prebiotic Effects of Partially Hydrolyzed Guar Gum on the Composition and Function of the Human Microbiota-Results from the PAGODA Trial. Nutrients 2020, 12, 1257. [Google Scholar] [CrossRef]
- Wilms, E.; An, R.; Smolinska, A.; Stevens, Y.; Weseler, A.R.; Elizalde, M.; Drittij, M.J.; Ioannou, A.; van Schooten, F.J.; Smidt, H.; et al. Galacto-oligosaccharides supplementation in prefrail older and healthy adults increased faecal bifidobacteria, but did not impact immune function and oxidative stress. Clin. Nutr. (Edinb. Scotl.) 2021, 40, 3019–3031. [Google Scholar] [CrossRef]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ (Clin. Res. Ed.) 2018, 361, k2179. [Google Scholar] [CrossRef]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Martínez, I.; Stegen, J.C.; Maldonado-Gómez, M.X.; Eren, A.M.; Siba, P.M.; Greenhill, A.R.; Walter, J. The gut microbiota of rural papua new guineans: Composition, diversity patterns, and ecological processes. Cell Rep. 2015, 11, 527–538. [Google Scholar] [CrossRef]
- Simpson, H.L.; Campbell, B.J. Review article: Dietary fibre-microbiota interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef]
- Segata, N. Gut Microbiome: Westernization and the Disappearance of Intestinal Diversity. Curr. Biol. CB 2015, 25, R611-3. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Peters, B.A.; Friedlander, C.; Freiman, H.J.; Goedert, J.J.; Sinha, R.; Miller, G.; Bernstein, M.A.; Hayes, R.B.; Ahn, J. Association of dietary fibre intake and gut microbiota in adults. Br. J. Nutr. 2018, 120, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Nguyen, S.M.; Yang, Y.; Xu, W.; Cai, H.; Wu, J.; Cai, Q.; Long, J.; Zheng, W.; Shu, X.O. Long-term diet quality is associated with gut microbiome diversity and composition among urban Chinese adults. Am. J. Clin. Nutr. 2021, 113, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Björkman, A.; Cai, K.; Liu, G.; Wang, C.; Li, Y.; Xia, H.; Sun, L.; Kristiansen, K.; Wang, J.; et al. Impact of a 3-Months Vegetarian Diet on the Gut Microbiota and Immune Repertoire. Front. Immunol. 2018, 9, 908. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Hermes, G.D.A.; Emanuel, E.C.; Holst, J.J.; Zoetendal, E.G.; Smidt, H.; Troost, F.; Schaap, F.G.; Damink, S.O.; Jocken, J.W.E.; et al. Effect of wheat bran derived prebiotic supplementation on gastrointestinal transit, gut microbiota, and metabolic health: A randomized controlled trial in healthy adults with a slow gut transit. Gut Microbes 2020, 12, 1704141. [Google Scholar] [CrossRef]
- Chen, O.; Sudakaran, S.; Blonquist, T.; Mah, E.; Durkee, S.; Bellamine, A. Effect of arabinogalactan on the gut microbiome: A randomized, double-blind, placebo-controlled, crossover trial in healthy adults. Nutrition 2021, 90, 111273. [Google Scholar] [CrossRef]
- Hiel, S.; Bindels, L.B.; Pachikian, B.D.; Kalala, G.; Broers, V.; Zamariola, G.; Chang, B.P.I.; Kambashi, B.; Rodriguez, J.; Cani, P.D.; et al. Effects of a diet based on inulin-rich vegetables on gut health and nutritional behavior in healthy humans. Am. J. Clin. Nutr. 2019, 109, 1683–1695. [Google Scholar] [CrossRef]
- Gondalia, S.V.; Wymond, B.; Benassi-Evans, B.; Berbezy, P.; Bird, A.R.; Belobrajdic, D.P. Substitution of Refined Conventional Wheat Flour with Wheat High in Resistant Starch Modulates the Intestinal Microbiota and Fecal Metabolites in Healthy Adults: A Randomized, Controlled Trial. J. Nutr. 2022, 152, 1426–1437. [Google Scholar] [CrossRef]
- DeMartino, P.; Johnston, E.A.; Petersen, K.S.; Kris-Etherton, P.M.; Cockburn, D.W. Additional Resistant Starch from One Potato Side Dish per Day Alters the Gut Microbiota but Not Fecal Short-Chain Fatty Acid Concentrations. Nutrients 2022, 14, 721. [Google Scholar] [CrossRef]
- Hughes, R.L.; Horn, W.H.; Finnegan, P.; Newman, J.W.; Marco, M.L.; Keim, N.L.; Kable, M.E. Resistant Starch Type 2 from Wheat Reduces Postprandial Glycemic Response with Concurrent Alterations in Gut Microbiota Composition. Nutrients 2021, 13, 645. [Google Scholar] [CrossRef]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. mBio 2019, 10, e02566-18. [Google Scholar] [CrossRef]
- Zhang, L.; Ouyang, Y.; Li, H.; Shen, L.; Ni, Y.; Fang, Q.; Wu, G.; Qian, L.; Xiao, Y.; Zhang, J.; et al. Metabolic phenotypes and the gut microbiota in response to dietary resistant starch type 2 in normal-weight subjects: A randomized crossover trial. Sci. Rep. 2019, 9, 4736. [Google Scholar] [CrossRef]
- Rodriguez, J.; Neyrinck, A.M.; Zhang, Z.; Seethaler, B.; Nazare, J.A.; Robles Sánchez, C.; Roumain, M.; Muccioli, G.G.; Bindels, L.B.; Cani, P.D.; et al. Metabolite profiling reveals the interaction of chitin-glucan with the gut microbiota. Gut Microbes 2020, 12, 1810530. [Google Scholar] [CrossRef]
- Fu, J.; Xu, K.; Ni, X.; Li, X.; Zhu, X.; Xu, W. Habitual Dietary Fiber Intake, Fecal Microbiota, and Hemoglobin A1c Level in Chinese Patients with Type 2 Diabetes. Nutrients 2022, 14, 1003. [Google Scholar] [CrossRef]
- Li, T.; Lu, X.; Yang, X. Evaluation of clinical safety and beneficial effects of stachyose-enriched α-galacto-oligosaccharides on gut microbiota and bowel function in humans. Food Funct. 2017, 8, 262–269. [Google Scholar] [CrossRef]
- Kiewiet, M.B.G.; Elderman, M.E.; El Aidy, S.; Burgerhof, J.G.M.; Visser, H.; Vaughan, E.E.; Faas, M.M.; de Vos, P. Flexibility of Gut Microbiota in Ageing Individuals during Dietary Fiber Long-Chain Inulin Intake. Mol. Nutr. Food Res. 2021, 65, e2000390. [Google Scholar] [CrossRef]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long-chain inulin extracted from globe artichoke (Cynara scolymus) in healthy human subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, Z.; Inoue, R.; Ozeki, M.; Okubo, T.; Takagi, T.; Honda, A.; Naito, Y. Effect of Repeated Consumption of Partially Hydrolyzed Guar Gum on Fecal Characteristics and Gut Microbiota: A Randomized, Double-Blind, Placebo-Controlled, and Parallel-Group Clinical Trial. Nutrients 2019, 11, 2170. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant starches types 2 and 4 have differential effects on the composition of the fecal microbiota in human subjects. PLoS ONE 2010, 5, e15046. [Google Scholar] [CrossRef] [PubMed]
- Vulevic, J.; Juric, A.; Walton, G.E.; Claus, S.P.; Tzortzis, G.; Toward, R.E.; Gibson, G.R. Influence of galacto-oligosaccharide mixture (B-GOS) on gut microbiota, immune parameters and metabonomics in elderly persons. Br. J. Nutr. 2015, 114, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Cloetens, L.; Broekaert, W.F.; Delaedt, Y.; Ollevier, F.; Courtin, C.M.; Delcour, J.A.; Rutgeerts, P.; Verbeke, K. Tolerance of arabinoxylan-oligosaccharides and their prebiotic activity in healthy subjects: A randomised, placebo-controlled cross-over study. Br. J. Nutr. 2010, 103, 703–713. [Google Scholar] [CrossRef]
- Walton, G.E.; Lu, C.; Trogh, I.; Arnaut, F.; Gibson, G.R. A randomised, double-blind, placebo controlled cross-over study to determine the gastrointestinal effects of consumption of arabinoxylan-oligosaccharides enriched bread in healthy volunteers. Nutr. J. 2012, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Tanihiro, R.; Sakano, K.; Oba, S.; Nakamura, C.; Ohki, K.; Hirota, T.; Sugiyama, H.; Ebihara, S.; Nakamura, Y. Effects of Yeast Mannan Which Promotes Beneficial Bacteroides on the Intestinal Environment and Skin Condition: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2020, 12, 3673. [Google Scholar] [CrossRef] [PubMed]
- Fastinger, N.D.; Karr-Lilienthal, L.K.; Spears, J.K.; Swanson, K.S.; Zinn, K.E.; Nava, G.M.; Ohkuma, K.; Kanahori, S.; Gordon, D.T.; Fahey, G.C., Jr. A novel resistant maltodextrin alters gastrointestinal tolerance factors, fecal characteristics, and fecal microbiota in healthy adult humans. J. Am. Coll. Nutr. 2008, 27, 356–366. [Google Scholar] [CrossRef]
- Calame, W.; Weseler, A.R.; Viebke, C.; Flynn, C.; Siemensma, A.D. Gum arabic establishes prebiotic functionality in healthy human volunteers in a dose-dependent manner. Br. J. Nutr. 2008, 100, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Turroni, F.; Ferrario, C.; van Sinderen, D.; Ventura, M. Meta-analysis of the human gut microbiome from urbanized and pre-agricultural populations. Environ. Microbiol. 2017, 19, 1379–1390. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science (New York N.Y.) 2011, 334, 105–108. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Mielgo-Ayuso, J.; Martín-Rodríguez, A.; Ramos-Campo, D.J.; Redondo-Flórez, L.; Tornero-Aguilera, J.F. The Burden of Carbohydrates in Health and Disease. Nutrients 2022, 14, 3809. [Google Scholar] [CrossRef]
- Fragiadakis, G.K.; Wastyk, H.C.; Robinson, J.L.; Sonnenburg, E.D.; Sonnenburg, J.L.; Gardner, C.D. Long-term dietary intervention reveals resilience of the gut microbiota despite changes in diet and weight. Am. J. Clin. Nutr. 2020, 111, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, L.; Zhai, Q.; Zhao, J.; Zhang, H.; Lee, Y.K.; Lu, W.; Li, M.; Chen, W. Chinese gut microbiota and its associations with staple food type, ethnicity, and urbanization. NPJ Biofilms Microbiomes 2021, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, D.; Sasaki, K.; Ikuta, N.; Yasuda, T.; Fukuda, I.; Kondo, A.; Osawa, R. Low amounts of dietary fibre increase in vitro production of short-chain fatty acids without changing human colonic microbiota structure. Sci. Rep. 2018, 8, 435. [Google Scholar] [CrossRef] [PubMed]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [PubMed]
- Gaundal, L.; Myhrstad, M.C.W.; Rud, I.; Gjøvaag, T.; Byfuglien, M.G.; Retterstøl, K.; Holven, K.B.; Ulven, S.M.; Telle-Hansen, V.H. Gut microbiota is associated with dietary intake and metabolic markers in healthy individuals. Food Nutr. Res. 2022, 66. [Google Scholar] [CrossRef]
- Kolida, S.; Meyer, D.; Gibson, G.R. A double-blind placebo-controlled study to establish the bifidogenic dose of inulin in healthy humans. Eur. J. Clin. Nutr. 2007, 61, 1189–1195. [Google Scholar] [CrossRef]
- Reimer, R.A.; Soto-Vaca, A.; Nicolucci, A.C.; Mayengbam, S.; Park, H.; Madsen, K.L.; Menon, R.; Vaughan, E.E. Effect of chicory inulin-type fructan-containing snack bars on the human gut microbiota in low dietary fiber consumers in a randomized crossover trial. Am. J. Clin. Nutr. 2020, 111, 1286–1296. [Google Scholar] [CrossRef]
- Deehan, E.C.; Yang, C.; Perez-Muñoz, M.E.; Nguyen, N.K.; Cheng, C.C.; Triador, L.; Zhang, Z.; Bakal, J.A.; Walter, J. Precision Microbiome Modulation with Discrete Dietary Fiber Structures Directs Short-Chain Fatty Acid Production. Cell Host Microbe 2020, 27, 389–404.e6. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourié, B.; Bornet, F.; Rambaud, J.C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Raskine, L.; Simoneau, G.; Paineau, D.; Bornet, F. The capacity of short-chain fructo-oligosaccharides to stimulate faecal bifidobacteria: A dose-response relationship study in healthy humans. Nutr. J. 2006, 5, 8. [Google Scholar] [CrossRef]
- Tandon, D.; Haque, M.M.; Gote, M.; Jain, M.; Bhaduri, A.; Dubey, A.K.; Mande, S.S. A prospective randomized, double-blind, placebo-controlled, dose-response relationship study to investigate efficacy of fructo-oligosaccharides (FOS) on human gut microflora. Sci. Rep. 2019, 9, 5473. [Google Scholar] [CrossRef] [PubMed]
- Maki, K.C.; Gibson, G.R.; Dickmann, R.S.; Kendall, C.W.; Chen, C.Y.; Costabile, A.; Comelli, E.M.; McKay, D.L.; Almeida, N.G.; Jenkins, D.; et al. Digestive and physiologic effects of a wheat bran extract, arabino-xylan-oligosaccharide, in breakfast cereal. Nutrition 2012, 28, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- François, I.E.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal health parameters in healthy adult human volunteers: A double-blind, randomised, placebo-controlled, cross-over trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef] [PubMed]
- Lefranc-Millot, C.; Guérin-Deremaux, L.; Wils, D.; Neut, C.; Miller, L.E.; Saniez-Degrave, M.H. Impact of a resistant dextrin on intestinal ecology: How altering the digestive ecosystem with NUTRIOSE®, a soluble fibre with prebiotic properties, may be beneficial for health. J. Int. Med. Res. 2012, 40, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.M.; Solch, R.J.; Dennis-Wall, J.C.; Ukhanova, M.; Nieves, C., Jr.; Mai, V.; Christman, M.C.; Gordon, D.T.; Langkamp-Henken, B. In healthy adults, resistant maltodextrin produces a greater change in fecal bifidobacteria counts and increases stool wet weight: A double-blind, randomized, controlled crossover study. Nutr. Res. (New York N.Y.) 2018, 60, 33–42. [Google Scholar] [CrossRef]
- Mai, V.; Burns, A.M.; Solch, R.J.; Dennis-Wall, J.C.; Ukhanova, M.; Langkamp-Henken, B. Resistant Maltodextrin Consumption in a Double-Blind, Randomized, Crossover Clinical Trial Induces Specific Changes in Potentially Beneficial Gut Bacteria. Nutrients 2022, 14, 2192. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Yang, G.; Wei, J.; Liu, P.; Zhang, Q.; Tian, Y.; Hou, G.; Meng, L.; Xin, Y.; Jiang, X. Role of the gut microbiota in type 2 diabetes and related diseases. Metab. Clin. Exp. 2021, 117, 154712. [Google Scholar] [CrossRef]
- Pedersen, C.; Gallagher, E.; Horton, F.; Ellis, R.J.; Ijaz, U.Z.; Wu, H.; Jaiyeola, E.; Diribe, O.; Duparc, T.; Cani, P.D.; et al. Host-microbiome interactions in human type 2 diabetes following prebiotic fibre (galacto-oligosaccharide) intake. Br. J. Nutr. 2016, 116, 1869–1877. [Google Scholar] [CrossRef]
- Gonai, M.; Shigehisa, A.; Kigawa, I.; Kurasaki, K.; Chonan, O.; Matsuki, T.; Yoshida, Y.; Aida, M.; Hamano, K.; Terauchi, Y. Galacto-oligosaccharides ameliorate dysbiotic Bifidobacteriaceae decline in Japanese patients with type 2 diabetes. Benef. Microbes 2017, 8, 705–716. [Google Scholar] [CrossRef]
- Birkeland, E.; Gharagozlian, S.; Birkeland, K.I.; Valeur, J.; Måge, I.; Rud, I.; Aas, A.M. Prebiotic effect of inulin-type fructans on faecal microbiota and short-chain fatty acids in type 2 diabetes: A randomised controlled trial. Eur. J. Nutr. 2020, 59, 3325–3338. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.M.; Davy, B.M.; Ponder, M.A.; McMillan, R.P.; Hughes, M.D.; Hulver, M.W.; Neilson, A.P.; Davy, K.P. Prebiotic Inulin Supplementation and Peripheral Insulin Sensitivity in adults at Elevated Risk for Type 2 Diabetes: A Pilot Randomized Controlled Trial. Nutrients 2021, 13, 3235. [Google Scholar] [CrossRef] [PubMed]
- Reimer, R.A.; Wharton, S.; Green, T.J.; Manjoo, P.; Ramay, H.R.; Lyon, M.R.; Gahler, R.J.; Wood, S. Effect of a functional fibre supplement on glycemic control when added to a year-long medically supervised weight management program in adults with type 2 diabetes. Eur. J. Nutr. 2021, 60, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Gallego, R.; Moreno-Indias, I.; Bea, A.M.; Sánchez-Alcoholado, L.; Fumanal, A.J.; Quesada-Molina, M.; Prieto-Martín, A.; Gutiérrez-Repiso, C.; Civeira, F.; Tinahones, F.J. An alcohol-free beer enriched with isomaltulose and a resistant dextrin modulates gut microbiome in subjects with type 2 diabetes mellitus and overweight or obesity: A pilot study. Food Funct. 2021, 12, 3635–3646. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Hermes, G.D.A.; Müller, M.; Bastings, J.; Vaughan, E.E.; van Den Berg, M.A.; Holst, J.J.; Venema, K.; Zoetendal, E.G.; Blaak, E.E. Fiber mixture-specific effect on distal colonic fermentation and metabolic health in lean but not in prediabetic men. Gut Microbes 2022, 14, 2009297. [Google Scholar] [CrossRef]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.E.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of Diet With Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology 2017, 153, 87–97.e3. [Google Scholar] [CrossRef]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Feng, Q.Q.; Ojo, O.O.; Wang, X.H. The Role of Dietary Fibre in Modulating Gut Microbiota Dysbiosis in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2020, 12, 3239. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Region | Intake Level (g/day) | Reference | Recommendation (g/day) | Reference | ||||
|---|---|---|---|---|---|---|---|---|
| Men | Women | Overall | Men | Women | Overall | |||
| Asia | ||||||||
| China | 19.4 | 17.6 | -- | China Health and Nutrition Survey (2011) [27] | -- | -- | 25–30 | Chinese Dietary Reference Intake (2017) [28] |
| Japan | 19.9 | 18.0 | 18.8 | National Health and Nutrition Survey in Japan (2019) [29] | 21 | 18 | 10 g/kcal | Dietary Reference Intake for Japanese (2020 Edition) [30] |
| North America | ||||||||
| USA | 18.1 | 15.2 | 16.6 | What We Eat in America (2017–2020) [31] | 38 | 25 | 14 g/kcal | Dietary Guidelines for Americans (2020–2025) [32] |
| Canada | 18.4 | 16.2 | -- | Canadian Community Health Survey (2015) [33] | 38 | 25 | -- | Canada’s Dietary Guidelines (2019) [34] |
| Oceania | ||||||||
| Australia | 24.8 | 21.1 | -- | Australian Health Survey (2011–2012) [25] | 38 | 28 | -- | Australian Dietary Guidelines (2013) [35] |
| Europe | ||||||||
| European Union | -- | -- | 25 | Scientific Opinion on Dietary Reference Values (EFSA-2010) [13] | ||||
| UK | -- | -- | 19.7 | UK National Diet and Nutrition Survey (2016–2019) [36] | -- | -- | 30 | Eatwell Guide (2016) [37] |
| France | 21.6 | 17.7 | 19.6 | National Individual Food Consumption 3 (2014–2015) [38] | -- | -- | 30 | Guidelines: development of nutritional references (2016) [39] |
| Denmark | -- | -- | 22 | Dietary habits in Denmark (2011–2013) [22] | 30 | 25 | 30 g/10MJ | Nordic Nutrition Recommendations (2012) [40] |
| Norway | 26 | 22 | 24 | Norkost 3 (2010–2011) [23] | ||||
| Germany | 24.8 | 23.1 | -- | National Consumption Research II (2005–2007) [24] | -- | -- | 30 or 14.6 g/kcal | The D-A-CH reference for nutrient intake (2021) [41] |
| Russia | 15 | 12 | -- | Russia Longitudinal Monitoring Survey (2018) [42] | -- | -- | 30 | Center for hygienic education of the population [43] |
| Author (Year) Region | Participants N | F:M a Mean Age (Years) | RCT Design | Dietary Fiber | Type of Fiber | Administrated Dosage (g/day) | Duration of Intervention | Changes in Gut Microbiota b | Cutoff Threshold (g/day) |
|---|---|---|---|---|---|---|---|---|
| Kolida, S. [116] (2007) UK | 30 | 15:15 26.5 ± 3.1 | double-blind crossover | Inulin | NSP | 0 g 5 g 8 g | 2-week intervention 1-week washout | ↑: Bifidobacteria * Depending on initial abundance | NO |
| Reimer, R. A. [117] (2020) Canada | 50 | 28:22 around 31 | double-blind crossover | Inulin-type fructans (ITF) | NSP | 0 g or 7 g (n = 25) 0 g or 3 g (n = 25) | 4-week intervention 4-week washout | ↔: α-diversity (Shannon index) ↔: β-diversity (Bray–Curtis distance) ↑: Bifidobacterium, Cellulomonas, Nesterenkonia, Brevibacterium ↓: Lachnospira, Oscillospira ↔: Fecal SCFAs | 7 g for Bifidobacterium, Cellulomonas, Nesterenkonia, Brevibacterium, Lachnospira, Oscillospira |
| Calame, W. [105] (2008) Netherlands | 48 | -- 30.9 ± 12.8 | double-blind | Gum arabic | NSP | 0 g (placebo-control, n = 8) 5 g (n = 8) 10 g (n = 8) 20 g (n = 8) 40 g (n = 8) 10 g inulin (positive control, n = 8) | 4-week intervention | ↑: Bifidobacteria, lactobacilli, bacteroides | 10 g for bifidobacteria, lactobacilli, bacteroides |
| Deehan, E. C. [118] (2020) Canada | 40 | 20:20 28.4 ± 8.1 | double-blind | RS4 | RS | Placebo (n = 10) Maize RS4 (n = 10) Potato RS4 (n = 10) Tapioca RS4 (n = 10) All 10 g for 1 week →20 g for 1 week →35 g for 1 week →50 g for 1 week | 4-week intervention | In maize group: ↓: α-diversity (Pielou and Shannon index) ↕: β-diversity (Bray-Curtis) ↑: Bifidobacterium ↑: Eubacterium rectale, Oscillibacter, Anaeromassilibacillus, Ruminococcus ↓: Agathobaculum butyriciproducens, Adlercreutzia equolifaciens ↑: Fecal butyrate In tapioca group: ↓: α-diversity (Pielou and Shannon index) ↕: β-diversity (Bray-Curtis) ↑: Bifidobacterium ↑: Parabacteroides distasonis, Faecalibacterium, Eisenbergiella ↓: Eubacterium hallii and Clostridium viride ↑: Fecal propionate *No effect of potato RS4 detected | In maize group: 20 g for α-diversity, β-diversity, Eubacterium rectale, Oscillibacter, Anaeromassilibacillus, Ruminococcus; 35 g for fecal butyrate; In tapioca group: 35 g for α-diversity, β-diversity, fecal propionate |
| Bouhnik, Y. [119] (1999) France | 40 | 22:18 29.6 | Short-chain fructo-oligosaccharides (SC-FOS) | ROS | 0 g (n = 8) 2.5 g (n = 8) 5 g (n = 8) 10 g (n = 8) 20 g (n = 8) | 7-day intervention | ↑: Bifidobacteria | 5 g for bifidobacteria | |
| Bouhnik, Y. [120] (2006) France | 40 | 22:18 29 ± 1.3 | Short-chain fructo-oligosaccharides (SC-FOS) | ROS | 0 g (n = 8) 2.5 g (n = 8) 5 g (n = 8) 7.5 g (n = 8) 10 g (n = 8) | 7-day intervention | ↑: Bifidobacteria (linear correlation) ↑: Total anaerobes | 10 g for total anaerobes | |
| Tandon, D. [121] (2019) India | 69 | 35:34 around 30 | double-blind | Fructo-oligosaccharides (FOS) | ROS | 0 g (n = 17) 2.5 g (n = 16) 5 g (n = 18) 10 g (n = 18) | 90-day intervention | ↑: α-diversity (Chao) ↑: Bifidobacterium, Lactobacillus, Faecalibacterium, Ruminococcus, Sutterella, Oscillospira *Reversal impact of prebiotics postdiscontinuation *Inconsistent results of analyses performed on data at two distinct levels of taxonomy (OTUs or genus) | 10 g for α-diversity, Bifidobacterium, Lactobacillus, Faecalibacterium, Ruminococcus, Sutterella, Oscillospira |
| Maki, K. C. [122] (2012) The US | 55 | -- 53.1 ± 12.6 | double-blind crossover | Arabinoxylan-oligosaccharide (AXOS) | ROS | 0 g 2.2 g 4.8 g | 3-week intervention 2-week washout | ↑: Bifidobacterium spp. | 4.8 g for Bifidobacterium spp. |
| François, I. E. [123] (2012) Belgium | 57 | 27:30 42 ± 17 | double-blind crossover | Arabinoxylan oligosaccharides (AXOS) | ROS | 0 g 2.4 g 8 g | 3-week intervention 2-week washout | ↑: Bifidobacteria ↑: Fecal SCFAs (acetic acid, propionic acid) | 8 g for bifidobacteria, fecal SCFAs |
| Fastinger, N. D. [104] (2008) The US | 38 | 19:19 around 27 | double-blind | Resistant maltodextrin | ROS | 0 g (n = 12) 7.5 g (n = 13) 15 g (n = 13) | 3-week intervention | ↑: Bifidobacterium (nonsignificant) ↑: Proportion of propionic acid (linear) | NO |
| Lefranc-Millot, C. [124] (2012) France | 48 | 24:24 28 | double-blind | Resistant dextrin | ROS | 0 g(n = 12) 10 g (n = 13) 15 g (n = 12) 20 g (n = 11) | 2-week intervention | ↔: Bifidobacterium spp., Lactobacillus spp. ↑: Bacteroides ↓: Clostridium perfringens ↓: Colonic pH | 10 g for Bacteroides; 15 g for Clostridium perfringens; 20 g for colonic pH |
| Burns, A. M. [125] (2018) The US | 49 | 28:21 26.3 ± 6.8 | double-blind cross-over | Resistant maltodextrin | ROS | 0 g (n = 16) 15 g (n = 17) 25 g (n = 16) | 3-week intervention 2-week washout | ↑: Bifidobacteria | 25 g for bifidobacteria |
| Mai, V. [126] (2022) The US | 49 | 28:21 26.3 ± 6.8 | double-blind cross-over | Resistant maltodextrin | ROS | 0 g (n = 16) 15 g (n = 17) 25 g (n = 16) | 3-week intervention 2-week washout | ↑: Fusicatenibacter saccharivorans ↑: Akkermansia muciniphila, Faecalibacterium prausnitzii (in individuals with low baseline counts) | 25 g for Akkermansia muciniphila, Faecalibacterium prausnitzii |
| Author Year Region | Subjects | F:M a Mean Age (years) | RCT Design | Fiber Type or Sources | Amount of Fiber (g/day) | Duration of Intervention | Change in Gut Microbiota b | Metabolic Effects b | Microbiome and metabolic Indicators |
|---|---|---|---|---|---|---|---|---|---|
| Pedersen, C. [129] (2016) UK | 29 well-controlled men | All men around 57 | Double-blind | galacto-oligosaccharide (GOS) | 5.5 (n = 14) placebo (n = 15) | 12 weeks | ↑: α-diversity (Shannon and Simpson indices) ↑: Bifidobacterium (close to significance) | No significant effect on glucose, insulin, or C-peptide fasting concentrations | Bifidobacterium positively correlated with total AUC of glucose and IL-6 |
| Gonai, M. [130] (2017) Japan | 52 patients | -- 50 ± 10 | Double-blind | galacto-oligosaccharide (GOS) | 10 (n = 27) placebo (n = 25) | 4 weeks | ↓: α-diversity (Observed OTUs) ↑: Bifidobacteriaceae ↓: Lachnospiraceae, Ruminococcaceae, Peptostreptococcaceae, Erysipelotrichaceae, Porphyromonadaceae | No clinical parameters changed significantly | Negative correlations of A1c with Bifidobacteriaceae and Peptostreptococcaceae; FPG with Peptostreptococcaceae; TG with Uminococcaceae; positive correlations of AST and ALT with Lachnospiraceae |
| Zhao, L. [65] (2018) China | 43 patients | -- 35–70 | Open-label | fiber in diet | high-fiber diet (n = 27) usual care (n = 16) | 84 days | ↕: β-diversity (Bray-Curtis) ↑: Bifidobacterium spp. and other SCFA producers ↑: CAZyme-encoding genes for starch and inulin degradation ↑: fhs for acetate and but for butyrate formation pathway ↑: acetate and butyrate | ↓: A1c ↑: Postprandial insulin | Higher acetate and butyrate coincided with a significantly greater AUC of postprandial glucagon-like peptide-1 and a higher level of fasting peptide YY, which partly improve A1c level |
| Birkeland, E. [131] (2020) Norway | 25 patients | 10:15 63.1 | Double-blinded, crossover | inulin-type fructans | 16 placebo | 6-week intervention 4-week washout | ↔: α-diversity (observed OTUs) ↕: β-diversity ↑: Bifidobacterium adolescentis, Bacteroides ovatus, Faecalibacterium prausnitzii ↓: Ruminococcus ↑: SCFAs (acetic acid and propionic acid) | Bifidobacterium adolescentis negatively related to fecal butyric acid | |
| Mitchell, C. M. [132] (2021) US | 22 adults at risk for T2D | 14:8 54.4 ± 8.3 | Double-blind | Inulin | 10 (n = 13) placebo (n = 9) | 6 weeks | ↑: Bifidobacteria | ↓: Fasting insulin, HOMA-IR | No significant correlation between changes in bifidobacteria and any outcome variables. |
| Reimer, R. A. [133] (2021) Canada | 290 adult patients with overweight/obesity | 198:92 around 55 | Double-blind | soluble viscous fiber | 15–20 (n = 147) isocaloric placebo (n = 143) | 52 weeks | ↑: Collinsella, Parabacteroides, Roseburia ↓: Faecalibacterium, Lactobacillus, Oscillibacter | ↓: A1c, BMI, WC, LDL | |
| Mateo-Gallego, R. [134] (2021) Spain | 14 patients with overweight or obesity | -- 56.1 ± 6.27 | Double-blinded, crossover | isomaltulose + resistant dextrin | 16.5 + 5.28 placebo | 10-week intervention 6–8 weeks’ washout | ↔: α-diversity (Shannon, Pielou, Observed features) ↔: β-diversity (weighted Unifrac) ↑: Parabacteroides ↓: Bacteroides, Odoribacter, Butyricimonas, Oscillospira | ↓: BMI, Blood glucose, HOMA-IR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, J.; Zheng, Y.; Gao, Y.; Xu, W. Dietary Fiber Intake and Gut Microbiota in Human Health. Microorganisms 2022, 10, 2507. https://doi.org/10.3390/microorganisms10122507
Fu J, Zheng Y, Gao Y, Xu W. Dietary Fiber Intake and Gut Microbiota in Human Health. Microorganisms. 2022; 10(12):2507. https://doi.org/10.3390/microorganisms10122507
Chicago/Turabian StyleFu, Jiongxing, Yan Zheng, Ying Gao, and Wanghong Xu. 2022. "Dietary Fiber Intake and Gut Microbiota in Human Health" Microorganisms 10, no. 12: 2507. https://doi.org/10.3390/microorganisms10122507
APA StyleFu, J., Zheng, Y., Gao, Y., & Xu, W. (2022). Dietary Fiber Intake and Gut Microbiota in Human Health. Microorganisms, 10(12), 2507. https://doi.org/10.3390/microorganisms10122507