
Novel Transcriptional and Translational Biomarkers of Tularemia Vaccine Efficacy in a Mouse Inhalation Model: Proof of Concept


 , ,
, ,
Abstract
1. Introduction
2. Methods
2.1. Bacteria
2.2. Vaccination of Mice
2.3. Transcriptomics
2.4. Multiplex and ELISA Assays
3. Results
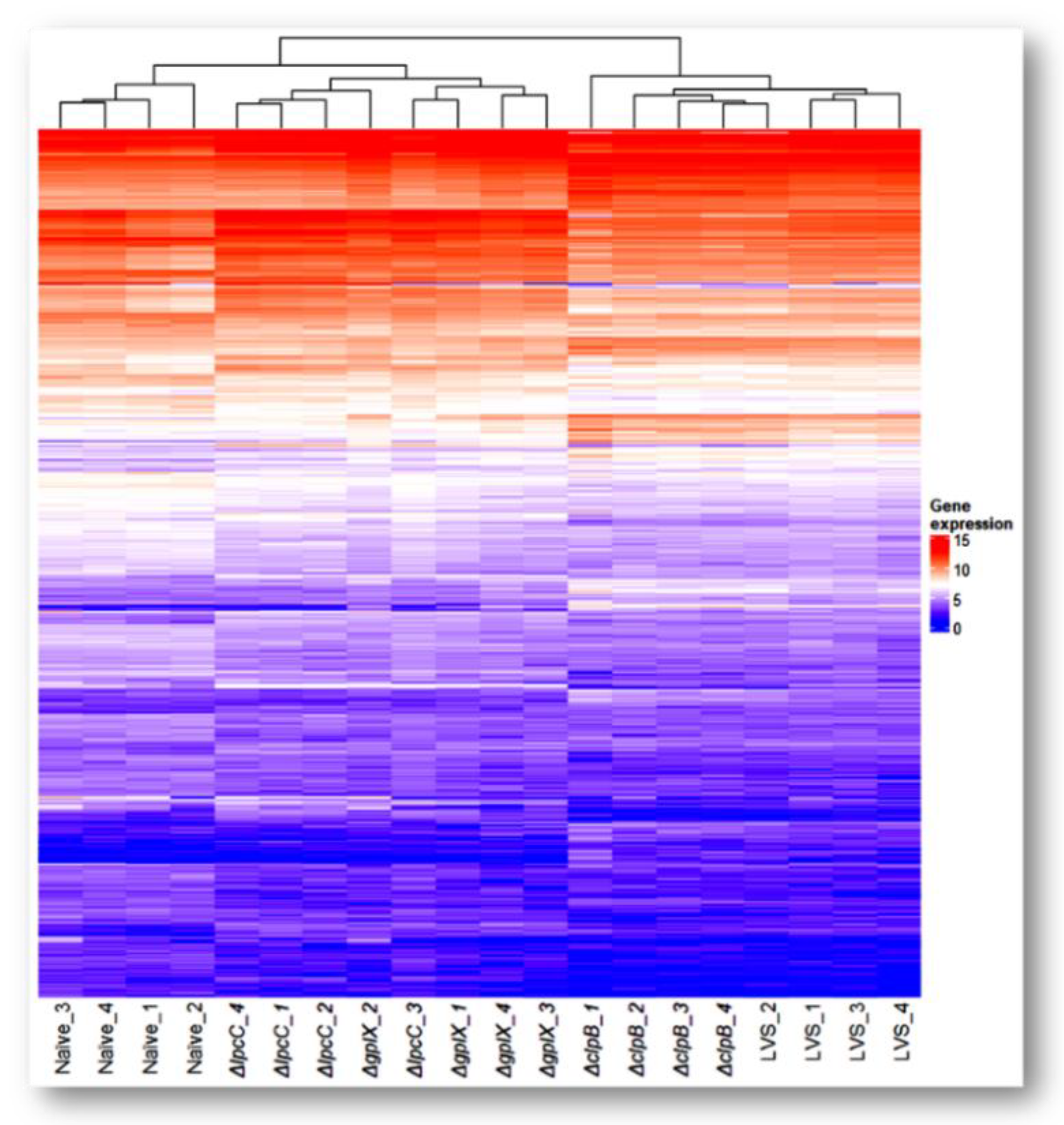
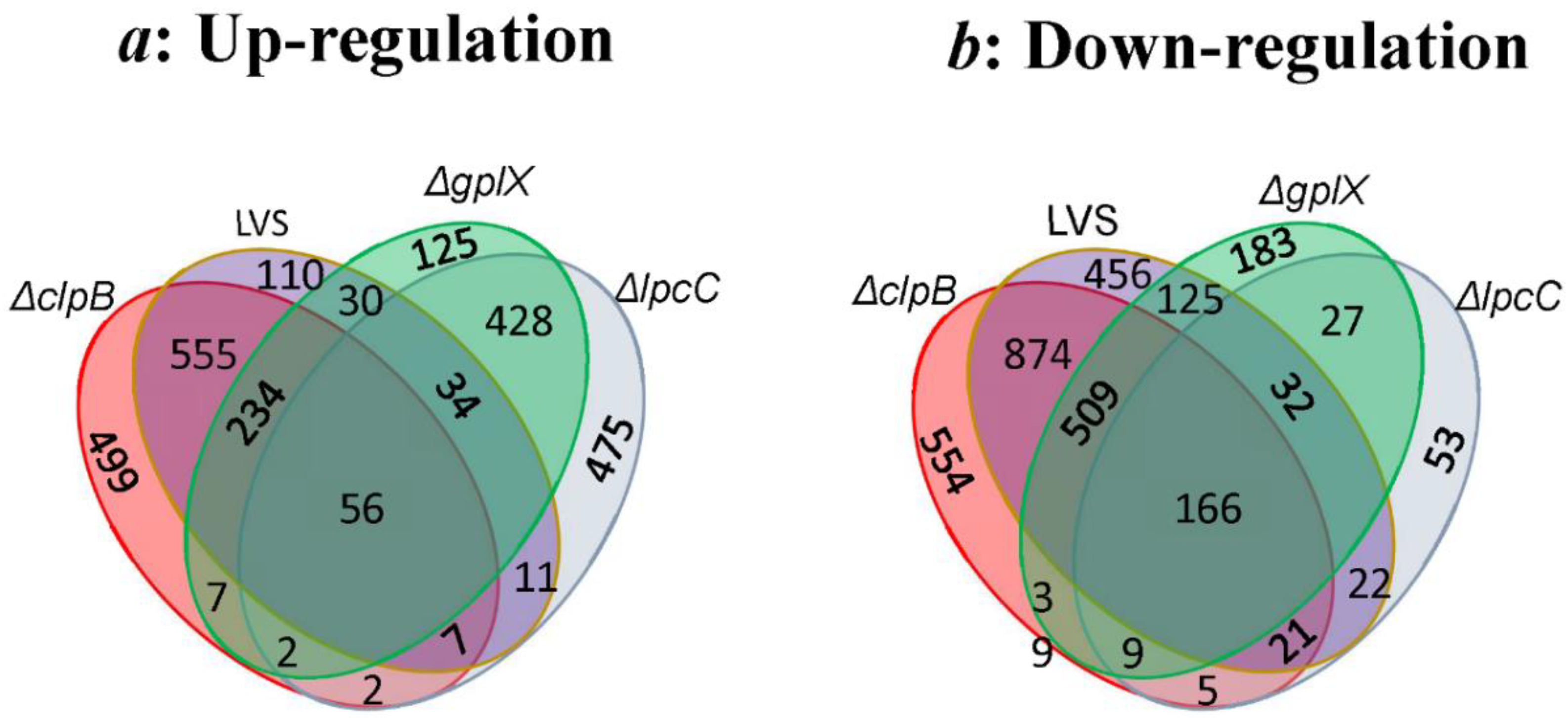
3.1. Transcriptomic Analysis
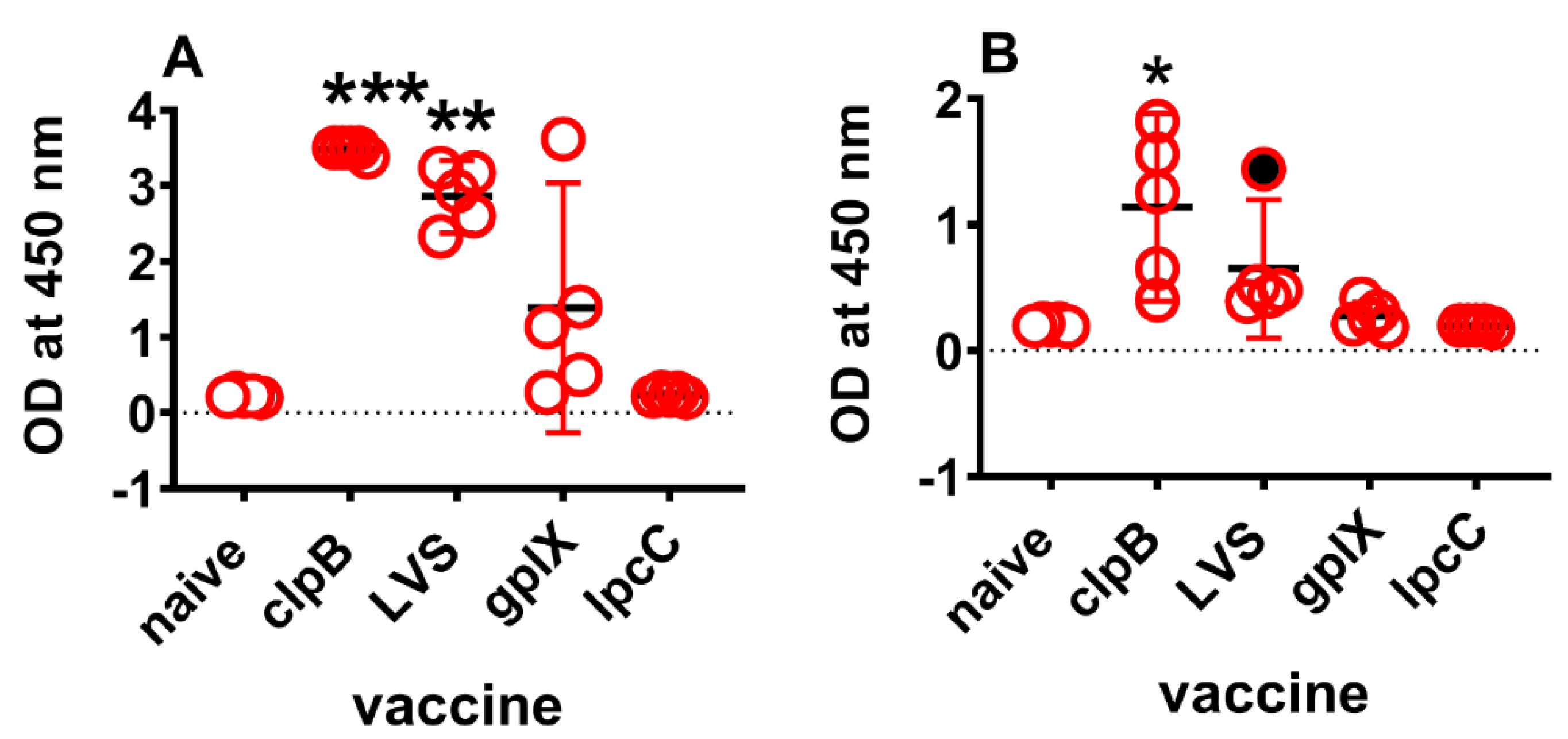
3.2. Proteomic Confirmation of Transcriptomics Findings
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sjostedt, A. Tularemia: History, epidemiology, pathogen physiology, and clinical manifestations. Ann. N. Y. Acad. Sci. 2007, 1105, 1–29. [Google Scholar] [CrossRef] [PubMed]
- McCruumb, F.R. Aerosol Infection of Man with Pasteurella Tularensis. Bacteriol. Rev. 1961, 25, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Oyston, P.C. Francisella tularensis: Unravelling the secrets of an intracellular pathogen. J. Med. Microbiol. 2008, 57 Pt 8, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Mares, C.A.; Li, Q.; Morris, E.G.; Teale, J.M. Features of sepsis caused by pulmonary infection with Francisella tularensis Type A strain. Microb. Pathog. 2011, 51, 39–47. [Google Scholar] [CrossRef]
- Bosio, C.M. The subversion of the immune system by francisella tularensis. Front. Microbiol. 2011, 2, 9. [Google Scholar] [CrossRef]
- Timofeev, V.; Titareva, G.; Bahtejeva, I.; Kombarova, T.; Kravchenko, T.; Mokrievich, A.; Dyatlov, I. The Comparative Virulence of Francisella tularensis Subsp. mediasiatica for Vaccinated Laboratory Animals. Microorganisms 2020, 8, 1403. [Google Scholar] [CrossRef]
- Conlan, J.W.; Chen, W.; Shen, H.; Webb, A.; KuoLee, R. Experimental tularemia in mice challenged by aerosol or intradermally with virulent strains of Francisella tularensis: Bacteriologic and histopathologic studies. Microb. Pathog. 2003, 34, 239–248. [Google Scholar] [CrossRef]
- Tarnvik, A.; Eriksson, M.; Sandstrom, G.; Sjostedt, A. Francisella tularensis—A model for studies of the immune response to intracellular bacteria in man. Immunology 1992, 76, 349–354. [Google Scholar]
- Schricker, R.L.; Eigelsbach, H.T.; Mitten, J.Q.; Hall, W.C. Pathogenesis of tularemia in monkeys aerogenically exposed to Francisella tularensis 425. Infect. Immun. 1972, 5, 734–744. [Google Scholar] [CrossRef]
- Meyer, C.G.; May, J. Germs employed as biological weapons. Anasthesiol. Intensivmed. Notf. Schmerzther. AINS 2002, 37, 538–546. [Google Scholar] [CrossRef]
- Kortepeter, M.G.; Parker, G.W. Potential biological weapons threats. Emerg. Infect. Dis. 1999, 5, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Oyston, P.C.; Sjostedt, A.; Titball, R.W. Tularaemia: Bioterrorism defence renews interest in Francisella tularensis. Nat. Rev. Microbiol. 2004, 2, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Dennis, D.T.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Friedlander, A.M.; Hauer, J.; Layton, M.; et al. Tularemia as a biological weapon: Medical and public health management. JAMA 2001, 285, 2763–2773. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M.; Skladal, P. Bacillus anthracis, Francisella tularensis and Yersinia pestis. The most important bacterial warfare agents-review. Folia Microbiol. 2009, 54, 263–272. [Google Scholar] [CrossRef]
- Eigelsbach, H.T.; Downs, C.M. Prophylactic effectiveness of live and killed tularemia vaccines. I. Production of vaccine and evaluation in the white mouse and guinea pig. J. Immunol. 1961, 87, 415–425. [Google Scholar]
- Saslaw, S.; Eigelsbach, H.T.; Wilson, H.E.; Prior, J.A.; Carhart, S. Tularemia vaccine study. I. Intracutaneous challenge. Arch. Intern. Med. 1961, 107, 689–701. [Google Scholar] [CrossRef]
- Saslaw, S.; Eigelsbach, H.T.; Prior, J.A.; Wilson, H.E.; Carhart, S. Tularemia vaccine study. II. Respiratory challenge. Arch. Intern. Med. 1961, 107, 702–714. [Google Scholar] [CrossRef]
- Hornick, R.B.; Dawkins, A.T.; Eigelsbach, H.T.; Tulis, J.J. Oral tularemia vaccine in man. Antimicrob. Agents Chemother. 1966, 6, 11–14. [Google Scholar]
- Cross, A.S.; Calia, F.M.; Edelman, R. From rabbits to humans: The contributions of Dr. Theodore, E. Woodward to tularemia research. Clin. Infect. Dis. 2007, 45 (Suppl. S1), S61–S67. [Google Scholar] [CrossRef]
- Burke, D.S. Immunization against tularemia: Analysis of the effectiveness of live Francisella tularensis vaccine in prevention of laboratory-acquired tularemia. J. Infect. Dis. 1977, 135, 55–60. [Google Scholar] [CrossRef]
- Hornick, R.B.; Eigelsbach, H.T. Aerogenic immunization of man with live Tularemia vaccine. Bacteriol. Rev. 1966, 30, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.R.; Watson, M.; Sell, T.K. Public Health Preparedness Funding: Key Programs and Trends From 2001 to 2017. Am. J. Public Health 2017, 107, S165–S167. [Google Scholar] [CrossRef]
- Twine, S.; Bystrom, M.; Chen, W.; Forsman, M.; Golovliov, I.; Johansson, A.; Kelly, J.; Lindgren, H.; Svensson, K.; Zingmark, C.; et al. A mutant of Francisella tularensis strain SCHU S4 lacking the ability to express a 58-kilodalton protein is attenuated for virulence and is an effective live vaccine. Infect. Immun. 2005, 73, 8345–8352. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Ryden, P.; Conlan, W. Molecular immune responses to aerosol challenge with Francisella tularensis in mice inoculated with live vaccine candidates of varying efficacy. PLoS ONE 2010, 5, e13349. [Google Scholar] [CrossRef] [PubMed]
- Conlan, J.W.; Shen, H.; Golovliov, I.; Zingmark, C.; Oyston, P.C.; Chen, W.; House, R.V.; Sjöstedt, A. Differential ability of novel attenuated targeted deletion mutants of Francisella tularensis subspecies tularensis strain SCHU S4 to protect mice against aerosol challenge with virulent bacteria: Effects of host background and route of immunization. Vaccine 2010, 28, 1824–1831. [Google Scholar] [CrossRef]
- Twine, S.; Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Ryden, P.; Conlan, W. BALB/c mice, but not C57BL/6 mice immunized with a ΔclpB mutant of Francisella tularensis subspecies tularensis are protected against respiratory challenge with wild-type bacteria: Association of protection with post-vaccination and post-challenge immune responses. Vaccine 2012, 30, 3634–3645. [Google Scholar]
- Ryden, P.; Twine, S.; Shen, H.; Harris, G.; Chen, W.; Sjostedt, A.; Conlan, W. Correlates of protection following vaccination of mice with gene deletion mutants of Francisella tularensis subspecies tularensis strain, SCHU S4 that elicit varying degrees of immunity to systemic and respiratory challenge with wild-type bacteria. Mol. Immunol. 2013, 54, 58–67. [Google Scholar] [CrossRef]
- Conlan, W.; Sjöstedt, A. Novel live vaccine candidates against airborne Francisella tularensis. In The Challenge of Highly Pathogenic Microorganisms; Springer: Dordrecht, The Netherlands, 2010; pp. 213–218. [Google Scholar]
- Golovliov, I.; Twine, S.M.; Shen, H.; Sjostedt, A.; Conlan, W. A ΔclpB Mutant of Francisella tularensis Subspecies holarctica Strain, FSC200, Is a More Effective Live Vaccine than F. tularensis LVS in a Mouse Respiratory Challenge Model of Tularemia. PLoS ONE 2013, 8, e78671. [Google Scholar] [CrossRef]
- Cowley, S.C.; Elkins, K.L. Immunity to francisella. Front. Microbiol. 2011, 2, 26. [Google Scholar] [CrossRef]
- Elkins, K.L.; Kurtz, S.L.; De Pascalis, R. Progress, challenges, and opportunities in Francisella vaccine development. Expert Rev. Vaccines 2016, 15, 1183–1196. [Google Scholar] [CrossRef]
- Allio, T. The FDA Animal Rule and its role in protecting human safety. Expert Opin. Drug Saf. 2018, 17, 971–973. [Google Scholar] [CrossRef]
- Beasley, D.W.C.; Brasel, T.L.; Comer, J.E. First vaccine approval under the FDA Animal Rule. NPJ Vaccines 2016, 1, 16013. [Google Scholar] [CrossRef]
- De Pascalis, R.; Chou, A.Y.; Bosio, C.M.; Huang, C.Y.; Follmann, D.A.; Elkins, K.L. Development of functional and molecular correlates of vaccine-induced protection for a model intracellular pathogen, F. tularensis LVS. PLoS Pathog. 2012, 8, e1002494. [Google Scholar] [CrossRef]
- De Pascalis, R.; Chou, A.Y.; Ryden, P.; Kennett, N.J.; Sjostedt, A.; Elkins, K.L. Models derived from in vitro analyses of spleen, liver, and lung leukocyte functions predict vaccine efficacy against the Francisella tularensis Live Vaccine Strain (LVS). mBio 2014, 5, e00936. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, R.; Mittereder, L.; Kennett, N.J.; Elkins, K.L. Activities of Murine Peripheral Blood Lymphocytes Provide Immune Correlates That Predict Francisella tularensis Vaccine Efficacy. Infect. Immun. 2016, 84, 1054–1061. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Pascalis, R.; Hahn, A.; Brook, H.M.; Ryden, P.; Donart, N.; Mittereder, L.; Frey, B.; Wu, T.H.; Elkins, K.L. A panel of correlates predicts vaccine-induced protection of rats against respiratory challenge with virulent Francisella tularensis. PLoS ONE 2018, 13, e0198140. [Google Scholar]
- Sjostedt, A.; Sandstrom, G.; Tarnvik, A.; Jaurin, B. Molecular cloning and expression of a T-cell stimulating membrane protein of Francisella tularensis. Microb. Pathog. 1989, 6, 403–414. [Google Scholar] [CrossRef]
- Golovliov, I.; Lindgren, H.; Eneslatt, K.; Conlan, W.; Mosnier, A.; Henry, T.; Sjöstedt, A. An In Vitro Co-culture Mouse Model Demonstrates Efficient Vaccine-Mediated Control of Francisella tularensis SCHU S4 and Identifies Nitric Oxide as a Predictor of Efficacy. Front. Cell. Infect. Microbiol. 2016, 6, 152. [Google Scholar] [CrossRef] [PubMed]
- Eneslatt, K.; Golovliov, I.; Ryden, P.; Sjostedt, A. Vaccine-Mediated Mechanisms Controlling Replication of Francisella tularensis in Human Peripheral Blood Mononuclear Cells Using a Co-culture System. Front. Cell. Infect. Microbiol. 2018, 8, 27. [Google Scholar] [CrossRef]
- Lindgren, H.; Eneslatt, K.; Golovliov, I.; Gelhaus, C.; Ryden, P.; Wu, T.; Sjöstedt, A. Vaccine-Mediated Mechanisms Controlling Francisella tularensis SCHU S4 Growth in a Rat Co-Culture System. Pathogens 2020, 9, 338. [Google Scholar] [CrossRef]
- Fuller, C.L.; Brittingham, K.C.; Hepburn, M.J.; Martin, J.W.; Petitt, P.L.; Pittman, P.R.; Bavari, S. Dominance of human innate immune responses in primary Francisella tularensis live vaccine strain vaccination. J. Allergy Clin. Immunol. 2006, 117, 1186–1188. [Google Scholar] [CrossRef] [PubMed]
- Fuller, C.L.; Brittingham, K.C.; Porter, M.W.; Hepburn, M.J.; Petitt, P.L.; Pittman, P.R.; Bavari, S. Transcriptome analysis of human immune responses following live vaccine strain (LVS) Francisella tularensis vaccination. Mol. Immunol. 2007, 44, 3173–3184. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goll, J.B.; Li, S.; Edwards, J.L.; Bosinger, S.E.; Jensen, T.L.; Wang, Y.; Hooper, W.F.; Gelber, C.E.; Sanders, K.L.; Anderson, E.J.; et al. Transcriptomic and Metabolic Responses to a Live-Attenuated Francisella tularensis Vaccine. Vaccines 2020, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Duong, D.M.; Goll, J.B.; Wood, D.C.; Jensen, T.L.; Yin, L.; Gelber, C.E.; Seyfried, N.T.; Anderson, E.; Natrajan, M.S.; et al. Proteomic Analysis of Human Immune Responses to Live-Attenuated Tularemia Vaccine. Vaccines 2020, 8, 413. [Google Scholar] [CrossRef] [PubMed]
- Twine, S.M.; Shen, H.; Kelly, J.F.; Chen, W.; Sjostedt, A.; Conlan, J.W. Virulence comparison in mice of distinct isolates of type A Francisella tularensis. Microb. Pathog. 2006, 40, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Hambleton, P.; Broster, M.; Dennis, P.; Henstridge, R.; Fitzgeorge, R.; Conlan, J. Survival of virulent Legionella pneumophila in aerosols. Epidemiol. Infect. 1983, 90, 451–460. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Tchagang, A.B.; Gawronski, A.; Berube, H.; Phan, S.; Famili, F.; Pan, Y. GOAL: A software tool for assessing biological significance of genes groups. BMC Bioinform. 2010, 11, 229. [Google Scholar] [CrossRef]
- Williams, M.S.; Baker, M.R.; Guina, T.; Hewitt, J.A.; Lanning, L.; Hill, H.; May, J.M.; Fogtman, B.; Pittman, P.R. Retrospective Analysis of Pneumonic Tularemia in Operation Whitecoat Human Subjects: Disease Progression and Tetracycline Efficacy. Front. Med. 2019, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Pittman, P.; Norris, S.; Coonan, K.; McKee, K. An assessment of Health Status among Medical Research Volunteers Who Served in the Project Whitecoat Program at Fort Detrick, Maryland. Mil. Med. 2005, 170, 183–187. [Google Scholar] [CrossRef]
- Saslaw, S.; Carhart, S. Studies with tularemia vaccines in volunteers. III. Serologic aspects following intracutaneous or respiratory challenge in both vaccinated and nonvaccinated volunteers. Am. J. Med. Sci. 1961, 241, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.M.; Powell, D.A.; Frelinger, J.A. Adaptive Immunity to Francisella tularensis and Considerations for Vaccine Development. Front. Cell. Infect. Microbiol. 2018, 8, 115. [Google Scholar] [CrossRef] [PubMed]
- Twine, S.M.; Petit, M.D.; Shen, H.; Mykytczuk, N.; Kelly, J.F.; Conlan, J.W. Immunoproteomic analysis of the murine antibody response to successful and failed immunization with live anti-Francisella vaccines. Biochem. Biophys. Res. Commun. 2006, 346, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Natrajan, M.S.; Rouphael, N.; Lai, L.; Kazmin, D.; Jensen, T.L.; Weiss, D.S.; Ibegbu, C.; Sztein, M.B.; Hooper, W.F.; Hill, H.; et al. Systems Vaccinology for a Live Attenuated Tularemia Vaccine Reveals Unique Transcriptional Signatures That Predict Humoral and Cellular Immune Responses. Vaccines 2019, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Eneslatt, K.; Rietz, C.; Ryden, P.; Stoven, S.; House, R.V.; Wolfraim, L.A.; Tärnvik, A.; Sjöstedt, A. Persistence of cell-mediated immunity three decades after vaccination with the live vaccine strain of Francisella tularensis. Eur. J. Immunol. 2011, 41, 974–980. [Google Scholar] [CrossRef]
- Pasetti, M.F.; Cuberos, L.; Horn, T.L.; Shearer, J.D.; Matthews, S.J.; House, R.V.; Sztein, M.B. An improved Francisella tularensis live vaccine strain (LVS) is well tolerated and highly immunogenic when administered to rabbits in escalating doses using various immunization routes. Vaccine 2008, 26, 1773–1785. [Google Scholar] [CrossRef]
- Tarnvik, A.; Sandstrom, G.; Lofgren, S. Time of lymphocyte response after onset of tularemia and after tularemia vaccination. J. Clin. Microbiol. 1979, 10, 854–860. [Google Scholar] [CrossRef]
- Conlan, J.W.; Sjostedt, A.; North, R.J. CD4+ and CD8+ T-cell-dependent and -independent host defense mechanisms can operate to control and resolve primary and secondary Francisella tularensis LVS infection in mice. Infect. Immun. 1994, 62, 5603–5607. [Google Scholar] [CrossRef]
- Cowley, S.C.; Elkins, K.L. Multiple T cell subsets control Francisella tularensis LVS intracellular growth without stimulation through macrophage interferon gamma receptors. J. Exp. Med. 2003, 198, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Elkins, K.L.; Cowley, S.C.; Conlan, J.W. Measurement of macrophage-mediated killing of intracellular bacteria, including Francisella and mycobacteria. Curr. Protoc. Immunol. 2011, 93, 14.25.1–14.25.13. [Google Scholar]
- Cole, L.E.; Santiago, A.; Barry, E.; Kang, T.J.; Shirey, K.A.; Roberts, Z.J.; Elkins, K.L.; Cross, A.S.; Vogel, S.N. Macrophage proinflammatory response to Francisella tularensis live vaccine strain requires coordination of multiple signaling pathways. J. Immunol. 2008, 180, 6885–6891. [Google Scholar] [CrossRef]
- Collazo, C.M.; Meierovics, A.I.; De Pascalis, R.; Wu, T.H.; Lyons, C.R.; Elkins, K.L. T cells from lungs and livers of Francisella tularensis-immune mice control the growth of intracellular bacteria. Infect. Immun. 2009, 77, 2010–2021. [Google Scholar] [CrossRef]
- Conlan, J.W.; Sjöstedt, A.; Gelhaus, H.C.; Fleming, P.; McRae, K.; Cobb, R.R.; De Pascalis, R.; Elkins, K.L. Modern Development and Production of a New Live Attenuated Bacterial Vaccine, SCHU S4 ΔclpB, to Prevent Tularemia. Pathogens 2021, 10, 795. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant | IN LD50 (CFU) | ID LD50 (CFU) | Survival against ID Challenge with SCHU S4 a | Survival against Respiratory Challenge with SCHU S4 b | Ref |
|---|---|---|---|---|---|
| LVS | 103 | >107 | 100% | 0–20% | [24,25] |
| ΔclpB | >104 <106 c | >107 | 100% | 60–100% | [24,27,47] |
| ΔgplX | NT | >107 | 80% | 0% | [27] |
| ΔlpcC | >103 <105 | >107 | 0% | 0% | [47] |
| ΔclpB/ Naïve | LVS/ Naïve | ΔgplX/ Naïve | ΔlpcC/ Naïve | ΔclpB/ LVS | ΔclpB/ ΔgplX | ΔclpB/ ΔlpcC | LVS/ ΔgplX | LVS/ ΔlpcC | ΔgplX/ ΔlpcC | |
|---|---|---|---|---|---|---|---|---|---|---|
| Up | 1362 | 1037 | 916 | 1015 | 108 | 961 | 1475 | 358 | 897 | 233 |
| Down | 2177 | 2205 | 1090 | 335 | 31 | 1219 | 1926 | 722 | 1546 | 164 |
| KEGG Pathway ID | KEGG Pathway Name | Total Known Genes | ΔclpB Alone | ΔclpB and LVS Shared | ΔclpB, LVS and ΔgplX Shared | Total Gene Counts a | |||
|---|---|---|---|---|---|---|---|---|---|
| padj | Matched Genes | padj | Matched Genes | padj | Matched Genes | ||||
| mmu04060 | Cytokine–cytokine receptor interaction | 292 | 3.98 × 10−8 | Ccr1, Il21, Cx3cl1, Cd70, Cxcr1, Il17f, Il23a, Osmr, Tnfrsf8, Tnfsf9, Tnfrsf1b, Ccl11, Ccl22, Ccl9, Ccl6, Fasl, Ccl3, Il23r, Ccr8, Fas, Gdf15, Ltbr, Cxcl16 | 1.15 × 10−14 | Il10, Il11, Il33, Il13, Il12b, Il2ra, Ccl17, Il15ra, Tnf, Tnfsf15, Bmp10, Csf2rb2, Ccl8, Tnfrsf1a, Csf2rb, Tnfrsf11b, Ccl4, Il1f9, Cxcl1, Ccr5, Cxcl2, Cxcl5, Il12rb2, Lif, Tnfsf10, Il17a, Ccl24, Il1a, Tnfrsf9, Il1b, Il1r2, Xcl1, Inhba, Tnfrsf12a, Inhbb, Csf2, Csf3 | 5.75 × 10−6 | Cxcl11, Il12rb1, Il22, Cxcl10, Il27, Il6, Ccl12, Cxcl9, Ccl7, Ifng, Ccl2, Il1rn, Cxcl3 | 73 |
| mmu04621 | NOD-like receptor signaling pathway | 211 | 3.29 × 10−5 | Jun, Pycard, Nlrp3, Nod2, Nod1, Mefv, Nlrp1b, Myd88, Tnf, Ifi207, Ifi206, Ifi204, Ripk3, Cybb, Il1b, Cxcl1, Cxcl2, Tlr4 | 1.16 × 10−14 | Txn1, Gbp5, Mapk13, Gbp7, Nampt, Nlrp1a, Oas1g, Il6, Ccl12, Gbp2b, Oas2, Oas1a, Irf7, Ccl2, Stat2, Stat1, Gbp2, Cxcl3, Gbp4, Gbp3 | 38 | ||
| mmu04062 | Chemokine signaling pathway | 192 | 7.03 × 10−4 | Ccr1, Hck, Ccl11, Ccl22, Ccl9, Cx3cl1, Ccl6, Cxcr1, Ccl3, Ccr8, Cxcl16, Gng12 | 9.03 × 10−3 | Ccl24, Ccl8, Ccl4, Gnb4, Stat3, Pik3r6, Xcl1, Cxcl1, Ccl17, Ccr5, Cxcl2, Cxcl5 | 1.19 × 10−4 | Cxcl11, Ccl12, Cxcl10, Cxcl9, Ccl7, Ccl2, Stat2, Stat1, Cxcl3 | 33 |
| mmu04612 | Antigen processing and presentation | 90 | 1.06 × 10−4 | H2-T24, Psme2b, Ctss, H2-K1, H2-Q4, Lgmn, H2-T3, H2-Q1, Hsp90aa1 | 2.23 × 10−7 | H2-T23, H2-T10, H2-Q6, H2-Q7, Hspa1b, Tap2, Hspa1a, H2-Q2, Psme1, Tnf, Hspa8, Gm11127, Tapbp, B2m | 0.01 | H2-T22, Ifng, Tap1, Psme2 | 27 |
| mmu04514 | Cell adhesion molecules | 174 | 7.15 × 10−5 | H2-T24, Sdc4, H2-K1, Ctla4, H2-Q4, Pdcd1, H2-T3, Tigit, H2-Q1, Nectin2, Selp, Mag, Cldn1 | 1.44 × 10−3 | H2-T23, H2-T10, H2-Q6, Sdc3, H2-Q7, H2-Q2, Nrcam, Pdcd1lg2, Itgam, Vcan, Ocln, Gm11127, Icam1 | 26 | ||
| mmu04610 | Complement and coagulation cascades | 93 | 3.32 × 10−3 | C1rb, C6, F13a1, Plat, Bdkrb1, Plaur, C2 | 6.68 × 10−9 | C1qb, Procr, C1ra, F10, Serping1, C5ar1, C3, F7, C1s2, Itgam, C1s1, Plau, C3ar1, Serpine1, A2m, Cfb | 23 | ||
| mmu04940 | Type I diabetes mellitus | 70 | 1.40 × 10−5 | H2-T24, Fasl, Ptprn, H2-K1, H2-Q4, Fas, Prf1, H2-T3, H2-Q1 | 4.11 × 10−6 | H2-T23, Il1a, H2-T10, Gm11127, H2-Q6, Il1b, H2-Q7, Il12b, H2-Q2, Tnf, Hspd1 | 0.03 | H2-T22, Ifng, Gzmb | 23 |
| mmu04630 | JAK-STAT signaling pathway | 168 | 2.37 × 10−5 | Il10, Il11, Socs3, Il12rb2, Il13, Il12b, Il2ra, Lif, Il15ra, Csf2rb2, Csf2rb, Myc, Stat3, Cdkn1a, Csf2, Csf3 | 1.41 × 10−3 | Il12rb1, Socs1, Il22, Il6, Ifng, Stat2, Stat1 | 23 | ||
| mmu04210 | Apoptosis | 136 | 1.29 × 10−4 | Ctss, Ctsd, Gadd45g, Fasl, Ctsz, Gadd45b, Fas, Prf1, Casp12, Bcl2a1b, Bcl2a1a | 1.76 × 10−3 | Ctsc, Csf2rb2, Jun, Tnfrsf1a, Csf2rb, Cycs, Fos, Daxx, Tnf, Tnfsf10, Tuba8 | 22 | ||
| mmu04620 | Toll-like receptor signaling pathway | 100 | 4.97 × 10−3 | Ccl3, Tlr8, Tlr7, Cd14, Ikbke, Tlr3, Tlr2 | 7.77 × 10−3 | Jun, Il1b, Ccl4, Il12b, Fos, Myd88, Tlr4, Tnf | 5.88 × 10−5 | Cxcl11, Mapk13, Il6, Cxcl10, Cxcl9, Irf7, Stat1 | 22 |
| mmu05332 | Graft-versus-host disease | 63 | 4.67 × 10−5 | H2-T24, Fasl, H2-K1, H2-Q4, Fas, Prf1, H2-T3, H2-Q1 | 6.84 × 10−5 | H2-T23, Il1a, H2-T10, Gm11127, H2-Q6, Il1b, H2-Q7, H2-Q2, Tnf | 3.54 × 10−3 | H2-T22, Il6, Ifng, Gzmb | 21 |
| mmu05330 | Allograft rejection | 63 | 4.67 × 10−5 | H2-T24, Fasl, H2-K1, H2-Q4, Fas, Prf1, H2-T3, H2-Q1 | 6.84 × 10−5 | Il10, H2-T23, H2-T10, Gm11127, H2-Q6, H2-Q7, Il12b, H2-Q2, Tnf | 0.02 | H2-T22, Ifng, Gzmb | 20 |
| mmu04650 | Natural killer cell mediated cytotoxicity | 121 | 9.74 × 10−4 | Raet1e, Klra7, Fasl, Sh2d1b2, H2-K1, Fas, Prf1, Ulbp1, Lcp2 | 0.02 | H2-T23, Raet1d, Klrk1, Fcer1g, Icam1, Csf2, Tnf, Tnfsf10 | 17 | ||
| mmu04640 | Hematopoietic cell lineage | 94 | 0.05 | Cd1d2, Sco1, Cd38, Cd14, Cd44 | 7.16 × 10−5 | Il11, Il1a, Itgam, Il1b, Il1r2, Il2ra, Anpep, Itga5, Csf2, Csf3, Tnf | 16 | ||
| mmu05320 | Autoimmune thyroid disease | 79 | 3.77 × 10−5 | H2-T24, Fasl, H2-K1, Ctla4, H2-Q4, Fas, Prf1, H2-T3, H2-Q1 | 7.28 × 10−3 | Il10, H2-T23, H2-T10, Gm11127, H2-Q6, H2-Q7, H2-Q2 | 16 | ||
| mmu05416 | Viral myocarditis | 88 | 0.01 | H2-T24, H2-K1, H2-Q4, Prf1, H2-T3, H2-Q1 | 3.59 × 10−3 | H2-T23, H2-T10, Gm11127, Cycs, H2-Q6, H2-Q7, Icam1, H2-Q2 | 14 | ||
| mmu00010 | Glycolysis/Gluconeogenesis | 67 | 2.64 × 10−6 | Aldh1b1, Pkm, Tpi1, Ldha, Eno1b, Pgk1, Pgam1, Eno1, Gapdh, Hk2, Pfkp | 11 | ||||
| mmu04623 | Cytosolic DNA-sensing pathway | 63 | 0.03 | Il33, Pycard, Ripk3, Il1b, Ccl4 | 3.60 × 10−5 | Il6, Cxcl10, Zbp1, Cgas, Irf7, Ifi202b | 11 | ||
| mmu03050 | Proteasome | 47 | 0.01 | Pomp, Psmb2, Psme1, Psma8, Psmb8 | 9.78 × 10−5 | Psma5, Ifng, Psme2, Psmb10, Psmb9 | 10 | ||
| mmu05133 | Pertussis | 77 | 0.02 | C1rb, Casp1, Cd14, Il23a, C2 | 1.11 × 10−16 | C1qb, Il10, C1ra, Jun, Pycard, Serping1, Nos2, Il12b, Itga5, Nlrp3, Fos, Nod1, Myd88, Tnf, C3, Il1a, C1s2, Itgam, C1s1, Il1b, Tlr4, Cxcl5 | 7.23 × 10−3 | Casp7, Mapk13, Il6, Irf1 | 31 |
| mmu05134 | Legionellosis | 61 | 0.04 | Bnip3, Casp1, Cd14, Tlr2 | 1.02 × 10−10 | Pycard, Cycs, Hspa1b, Il12b, Hspa1a, Myd88, Tnf, C3, Hspa8, Itgam, Il1b, Cxcl1, Cxcl2, Tlr4, Hspd1 | 0.02 | Casp7, Il6, Cxcl3 | 22 |
| mmu05321 | Inflammatory bowel disease | 62 | 9.51 × 10−3 | Il21, Il23r, Il17f, Il23a, Tlr2 | 1.41 × 10−7 | Il10, Il1a, Jun, Il12rb2, Il1b, Il13, Il12b, Stat3, Nod2, Tlr4, Tnf, Il17a | 3.67 × 10−4 | Il12rb1, Il22, Il6, Ifng, Stat1 | 22 |
| mmu04657 | IL-17 signaling pathway | 91 | 6.22 × 10−4 | Ccl11, Il17f, Ikbke, Tnfaip3, S100a9, S100a8, Hsp90aa1, Lcn2 | 2.57 × 10−7 | Jun, Il13, Mmp3, Fos, Ccl17, Cebpb, Tnf, Il17a, Il1b, Cxcl1, Csf2, Cxcl2, Csf3, Cxcl5 | 1.95 × 10−8 | Mapk13, Il6, Mmp13, Ccl12, Cxcl10, Ccl7, Ifng, Ccl2, Cxcl3, Ptgs2 | 32 |
| mmu04061 | Viral protein interaction with cytokine and cytokine receptor | 95 | 4.35 × 10−6 | Ccr1, Tnfrsf1b, Ccl11, Ccl22, Ccl9, Cx3cl1, Ccl6, Cxcr1, Ccl3, Ccr8, Ltbr | 4.45 × 10−7 | Il10, Il2ra, Ccl17, Tnf, Tnfsf10, Ccl24, Ccl8, Tnfrsf1a, Ccl4, Xcl1, Cxcl1, Ccr5, Cxcl2, Cxcl5 | 4.30 × 10−6 | Cxcl11, Il6, Ccl12, Cxcl10, Cxcl9, Ccl7, Ccl2, Cxcl3 | 33 |
| mmu04668 | TNF signaling pathway | 113 | 9.56 × 10−3 | Tnfrsf1b, Cx3cl1, Mmp14, Creb3l3, Fas, Creb3l1, Tnfaip3 | 2.68 × 10−9 | Jun, Socs3, Mmp3, Lif, Nod2, Fos, Creb5, Cebpb, Tnf, Tnfrsf1a, Ripk3, Il1b, Bcl3, Icam1, Cxcl1, Csf2, Cxcl2, Cxcl5 | 9.97 × 10−10 | Casp7, Mapk13, Il6, Ccl12, Cxcl10, Gm5431, Mlkl, Irf1, Ccl2, Ifi47, Cxcl3, Ptgs2 | 37 |
| mmu05146 | Amoebiasis | 107 | 3.97 × 10−4 | Cd1d2, Col3a1, Arg2, Col1a1, Gnal, Col4a2, Cd14, Serpinb6b, Tlr2 | 5.31 × 10−8 | Il10, Arg1, Nos2, Col4a1, Prdx1, Il12b, Tnf, Itgam, Il1b, Serpinb9, Il1r2, Cxcl1, Lamc2, Csf2, Cxcl2, Tlr4 | 4.24 × 10−3 | Serpinb9b, Il6, Ifng, Ctsg, Cxcl3 | 30 |
| mmu05323 | Rheumatoid arthritis | 87 | 0.04 | Atp6v1a, Ccl3, Ctla4, Il23a, Tlr2 | 1.44 × 10−7 | Il11, Jun, Mmp3, Fos, Tnf, Il17a, Il1a, Il1b, Icam1, Cxcl1, Csf2, Cxcl2, Tlr4, Cxcl5 | 1.72 × 10−3 | Il6, Ccl12, Ifng, Ccl2, Cxcl3 | 24 |
| mmu05140 | Leishmaniasis | 70 | 7.96 × 10−9 | Il10, Jun, Fcgr3, Nos2, Il12b, Fos, Myd88, Tnf, C3, Il1a, Itgam, Cybb, Il1b, Tlr4 | 6.45 × 10−4 | Fcgr1, Mapk13, Ifng, Stat1, Ptgs2 | 19 | ||
| mmu05144 | Malaria | 57 | 4.88 × 10−7 | Il10, Klrb1b, Klrk1, Hgf, Il1b, Icam1, Csf3, Myd88, Tlr4, Tnf, Thbs4 | 2.46 × 10−3 | Il6, Ccl12, Ifng, Ccl2 | 15 | ||
| mmu05143 | African trypanosomiasis | 39 | 8.73 × 10−3 | Fasl, Fas, Ido2, Ido1 | 7.65 × 10−4 | Il10, Il1b, Icam1, Il12b, Myd88, Tnf | 10 | ||
| mmu04625 | C-type lectin receptor signaling pathway | 112 | 0.03 | Ccl22, Egr2, Card9, Casp1, Il23a, Ikbke | 8.09 × 10−5 | Il10, Clec4d, Jun, Pycard, Fcer1g, Il1b, Bcl3, Clec7a, Il12b, Nlrp3, Ccl17, Tnf | 1.21 × 10−4 | Mapk13, Il6, Irf1, Stat2, Stat1, Clec4e, Ptgs2 | 25 |
| mmu05167 | Kaposi sarcoma-associated herpesvirus infection | 225 | 2.73 × 10−3 | Ccr1, H2-T24, Hck, H2-K1, H2-Q4, Ccr8, Fas, H2-T3, Ikbke, Gng12, H2-Q1, Tlr3 | 2.35 × 10−8 | H2-T23, Jun, H2-T10, Cycs, H2-Q6, H2-Q7, H2-Q2, Fos, Cd200r4, C3, Hif1a, Rcn1, Tnfrsf1a, Gm11127, Myc, Gnb4, Icam1, Stat3, Pik3r6, Cxcl1, Ccr5, Cdkn1a, Csf2, Cxcl2 | 7.64 × 10−5 | Mapk13, H2-T22, Il6, Eif2ak2, Irf7, Stat2, Bak1, Stat1, Cxcl3, Ptgs2 | 46 |
| mmu04145 | Phagosome | 182 | 4.35 × 10−4 | H2-T24, C1rb, Ctss, Lox, Atp6v1a, Lamp2, H2-K1, H2-Q4, Cd14, H2-T3, H2-Q1, Tlr2 | 1.01 × 10−6 | H2-T23, C1ra, Fcgr3, H2-T10, Tubb6, H2-Q6, Clec7a, H2-Q7, Tap2, H2-Q2, Itga5, Tuba8, C3, Itgam, Gm11127, Cybb, Olr1, Tlr4, Thbs4 | 9.58 × 10−3 | Fcgr1, Msr1, H2-T22, Tubb3, Tap1, Mpo | 37 |
| mmu05171 | Coronavirus disease—COVID-19 | 247 | 2.06 × 10−3 | C1rb, Sting1, F13a1, Casp1, Ikbke, Hbegf, C2, Selp, C6, Tlr8, Tlr7, Tlr3, Tlr2 | 3.40 × 10−8 | C1qb, Mx2, C5ar1, Mx1, Il12b, Tnf, C3, Tnfrsf1a, C3ar1, Ifih1, C1ra, Jun, Mmp3, Nlrp3, Fos, Myd88, C1s2, C1s1, Cybb, Il1b, Stat3, Csf2, Csf3, Tlr4, Cfb | 5.61 × 10−6 | Oas1g, Mapk13, Il6, Ccl12, Cxcl10, Oas2, Cgas, Oas1a, Eif2ak2, Ccl2, Stat2, Stat1 | 50 |
| mmu00220 | Arginine biosynthesis | 20 | 2.22 × 10−3 | Arg1, Got1, Nos2, Ass1 | 4 | ||||
| mmu00770 | Pantothenate and CoA biosynthesis | 20 | 2.22 × 10−3 | Aldh1b1, Vnn3, Dpys, Bcat1 | 4 | ||||
| mmu00524 | Neomycin, kanamycin and gentamicin biosynthesis | 5 | 0.05 | Hk3 | 1 | ||||
| mmu05417 | Lipid and atherosclerosis | 216 | 5.74 × 10−3 | Selp, Ero1a, Lox, Fasl, Ccl3, Fas, Casp1, Cd14, Ikbke, Hsp90aa1, Tlr2 | 1.05 × 10−8 | Jun, Pycard, Cycs, Mmp3, Hspa1b, Il12b, Hspa1a, Nlrp3, Fos, Myd88, Sod2, Tnf, Tnfsf10, Hspa8, Tnfrsf1a, Cybb, Il1b, Icam1, Stat3, Olr1, Cxcl1, Cxcl2, Tlr4, Hspd1 | 5.72 × 10−3 | Casp7, Mapk13, Il6, Ccl12, Irf7, Ccl2, Cxcl3 | 42 |
| mmu05169 | Epstein-Barr virus infection | 231 | 3.38 × 10−3 | H2-T24, Gadd45g, H2-K1, Gadd45b, H2-Q4, Fas, H2-T3, Ikbke, Tnfaip3, H2-Q1, Cd44, Tlr2 | 1.07 × 10−4 | H2-T23, Jun, H2-T10, Cycs, H2-Q6, H2-Q7, Tap2, H2-Q2, Myd88, Tnf, Gm11127, Myc, Icam1, Stat3, Vim, Cdkn1a, Tapbp, B2m | 4.16 × 10−7 | Mapk13, H2-T22, Cxcl10, Eif2ak2, Tap1, Oas1g, Il6, Oas2, Oas1a, Irf7, Stat2, Bak1, Stat1 | 43 |
| mmu05145 | Toxoplasmosis | 110 | 4.83 × 10−7 | Il10, Nos2, Cycs, Hspa1b, Il12b, Hspa1a, Myd88, Tnf, Hspa8, Tnfrsf1a, Stat3, Pik3r6, Lamc2, Ccr5, Tlr4 | 4.77 × 10−3 | Socs1, Mapk13, Ifng, Irgm1, Stat1 | 20 | ||
| mmu05142 | Chagas disease | 103 | 6.84 × 10−6 | C1qb, Il10, Jun, Nos2, Il12b, Fos, Myd88, Tnf, C3, Tnfrsf1a, Il1b, Serpine1, Tlr4 | 3.61 × 10−3 | Mapk13, Il6, Ccl12, Ifng, Ccl2 | 18 | ||
| mmu05164 | Influenza A | 173 | 8.65 × 10−6 | Ifih1, Il33, Pycard, Socs3, Cycs, Mx2, Mx1, Il12b, Nlrp3, Myd88, Tnf, Tnfsf10, Il1a, Tnfrsf1a, Il1b, Icam1, Tlr4 | 1.37 × 10−8 | Cxcl10, Eif2ak2, Oas1g, Il6, Ccl12, Ifng, Oas2, Oas1a, Irf7, Ccl2, Stat2, Bak1, Stat1 | 30 | ||
| mmu05163 | Human cytomegalovirus infection | 256 | 9.67 × 10−4 | Ccr1, H2-T24, Cx3cl1, Sting1, H2-K1, Creb3l3, H2-Q4, Creb3l1, H2-T3, H2-Q1, Fasl, Ccl3, Fas, Gng12 | 1.31 × 10−5 | H2-T23, H2-T10, Cycs, H2-Q6, H2-Q7, Tap2, H2-Q2, Creb5, Tnf, Tnfrsf1a, Gm11127, Il1b, Myc, Ccl4, Gnb4, Stat3, Cdkn2a, Ccr5, Cdkn1a, Tapbp, B2m | 9.89 × 10−4 | Mapk13, H2-T22, Il6, Ccl12, Cgas, Ccl2, Tap1, Bak1, Ptgs2 | 44 |
| mmu05162 | Measles | 146 | 3.92 × 10−6 | Ifih1, Jun, Cycs, Mx2, Mx1, Hspa1b, Il12b, Il2ra, Hspa1a, Fos, Myd88, Hspa8, Il1a, Il1b, Stat3, Tlr4 | 1.38 × 10−5 | Oas1g, Il6, Oas2, Oas1a, Eif2ak2, Irf7, Stat2, Bak1, Stat1 | 25 | ||
| mmu04064 | NF-kappa B signaling pathway | 105 | 6.47 × 10−3 | Gadd45g, Gadd45b, Cd14, Ltbr, Tnfaip3, Bcl2a1b, Bcl2a1a | 8.17 × 10−4 | Tnfrsf1a, Plau, Il1b, Ccl4, Icam1, Cxcl1, Cxcl2, Myd88, Tlr4, Tnf | 17 | ||
| mmu05310 | Asthma | 25 | 5.17 × 10−3 | Il10, Fcer1g, Il13, Tnf | 4 | ||||
| mmu04217 | Necroptosis | 174 | 0.03 | Fasl, Fth1, Chmp4b, Fas, Casp1, Tnfaip3, Tlr3, Hsp90aa1 | 1.44 × 10−3 | Il33, Pycard, Nlrp3, Pla2g4a, Tnf, Tnfsf10, Il1a, Tnfrsf1a, Ripk3, Cybb, Il1b, Stat3, Tlr4 | 7.76 × 10−3 | Zbp1, Ifng, Mlkl, Eif2ak2, Stat2, Stat1 | 27 |
| mmu05170 | Human immunodeficiency virus 1 infection | 240 | 0.01 | H2-T24, Tnfrsf1b, Fasl, Sting1, H2-K1, H2-Q4, Fas, H2-T3, Gng12, H2-Q1, Tlr2 | 1.68 × 10−5 | H2-T23, Jun, H2-T10, Cycs, H2-Q6, H2-Q7, Tap2, H2-Q2, Fos, Myd88, Tnf, Bst2, Tnfrsf1a, Gm11127, Gnb4, Samhd1, Ccr5, Tapbp, B2m, Tlr4 | 0.03 | Mapk13, H2-T22, Cgas, Trim30d, Tap1, Bak1 | 37 |
| mmu05161 | Hepatitis B | 163 | 7.29 × 10−3 | Egr2, Fasl, Creb3l3, Fas, Creb3l1, Ikbke, Casp12, Tlr3, Tlr2 | 7.05 × 10−3 | Ifih1, Jun, Cycs, Myc, Stat3, Fos, Cdkn1a, Creb5, Myd88, Tlr4, Tnf | 0.02 | Mapk13, Il6, Irf7, Stat2, Stat1 | 25 |
| mmu01230 | Biosynthesis of amino acids | 79 | 2.16 × 10−6 | Pkm, Tpi1, Arg1, Got1, Eno1b, Pgk1, Pgam1, Eno1, Gapdh, Bcat1, Pfkp, Ass1 | 12 | ||||
| mmu05160 | Hepatitis C | 165 | 8.80 × 10−4 | Socs3, Cycs, Ifit1bl1, Mx2, Mx1, Nr1h3, Tnf, Ywhag, Ocln, Tnfrsf1a, Myc, Stat3, Cdkn1a | 7.48 × 10−8 | Oas1g, Cxcl10, Ifng, Oas2, Oas1a, Ifit1bl2, Eif2ak2, Irf7, Stat2, Bak1, Stat1, Ifit1 | 25 | ||
| mmu05235 | PD-L1 expression and PD-1 checkpoint pathway in cancer | 88 | 3.59 × 10−3 | Hif1a, Jun, Stat3, Fos, Batf3, Myd88, Tlr4, Batf | 1.81 × 10−3 | Cd274, Mapk13, Ifng, Stat1, Batf2 | 13 | ||
| mmu04380 | Osteoclast differentiation | 128 | 2.90 × 10−4 | Il1a, Lilrb4a, Jun, Sirpb1a, Socs3, Fcgr3, Tnfrsf1a, Tnfrsf11b, Il1b, Fos, Tnf, Fosl2 | 1.71 × 10−3 | Socs1, Fcgr1, Mapk13, Ifng, Stat2, Stat1 | 18 | ||
| mmu05152 | Tuberculosis | 180 | 8.51 × 10−7 | Il10, Fcgr3, Nos2, Vdr, Cycs, Clec7a, Il12b, Nod2, Cebpb, Myd88, Tnf, C3, Il1a, Itgam, Tnfrsf1a, Fcer1g, Il1b, Tlr4, Hspd1 | 9.10 × 10−3 | Fcgr1, Mapk13, Il6, Ifng, Stat1, Clec4e | 25 | ||
| mmu04933 | AGE-RAGE signaling pathway in diabetic complications | 101 | 2.35 × 10−3 | Il1a, Jun, Col4a1, Cybb, Il1b, Icam1, Stat3, Serpine1, Tnf | 3.31 × 10−3 | Mapk13, Il6, Ccl12, Ccl2, Stat1 | 14 | ||
| mmu04066 | HIF-1 signaling pathway | 114 | 7.73 × 10−7 | Nos2, Eno1b, Timp1, Eno1, Gapdh, Hk2, Hif1a, Ldha, Cybb, Pgk1, Stat3, Serpine1, Cdkn1a, Tlr4, Pfkp | 15 | ||||
| mmu04659 | Th17 cell differentiation | 104 | 0.03 | Hif1a, Jun, Il1b, Il2ra, Stat3, Fos, Il17a | 5.80 × 10−4 | Il12rb1, Il22, Mapk13, Il6, Ifng, Stat1 | 13 | ||
| mmu05150 | Staphylococcus aureus infection | 124 | 2.29 × 10−6 | C1qb, Il10, C1ra, Fcgr3, Krt14, C5ar1, Fpr1, C3, C1s2, Itgam, C1s1, Ptafr, C3ar1, Icam1, Cfb | 15 | ||||
| mmu05230 | Central carbon metabolism in cancer | 69 | 7.45 × 10−4 | Hif1a, Pkm, Ldha, Myc, Pgam1, Slc16a3, Hk2, Pfkp | 8 | ||||
| mmu04658 | Th1 and Th2 cell differentiation | 88 | 0.04 | Jun, Il12rb2, Il13, Il12b, Il2ra, Fos | 0.01 | Il12rb1, Mapk13, Ifng, Stat1 | 10 | ||
| mmu05203 | Viral carcinogenesis | 229 | 8.77 × 10−3 | H2-T24, Egr2, Scin, H2-K1, Creb3l3, H2-Q4, Ccr8, Creb3l1, H2-T3, Ltbr, H2-Q1 | 2.36 × 10−3 | H2-T23, Jun, H2-T10, H2-Q6, H2-Q7, H2-Q2, Creb5, Ywhag, C3, Pkm, Gm11127, Stat3, Cdkn2a, Ccr5, Cdkn1a | 26 | ||
| mmu05165 | Human papillomavirus infection | 362 | 0.02 | H2-T24, Col1a1, Atp6v1a, Col4a2, H2-K1, Creb3l3, H2-Q4, Creb3l1, H2-T3, Ikbke, H2-Q1, Fasl, Fas, Tlr3 | 7.78 × 10−3 | H2-T23, H2-T10, Col4a1, Fzd7, H2-Q6, Mx2, H2-Q7, Mx1, Tnc, H2-Q2, Itga5, Creb5, Tnf, Pkm, Tnfrsf1a, Gm11127, Lamc2, Cdkn1a, Thbs4 | 0.03 | Oasl1, H2-T22, Irf1, Eif2ak2, Stat2, Bak1, Stat1, Ptgs2 | 41 |
| mmu05135 | Yersinia infection | 134 | 1.56 × 10−3 | Il10, Jun, Pycard, Il1b, Itga5, Nlrp3, Fos, Mefv, Myd88, Tlr4, Tnf | 0.04 | Mapk13, Il6, Ccl12, Ccl2 | 15 | ||
| mmu00330 | Arginine and proline metabolism | 54 | 4.24 × 10−3 | Aldh1b1, Gatm, Arg1, Got1, Nos2, Cndp2 | 6 | ||||
| mmu04930 | Type II diabetes mellitus | 48 | 0.01 | Socs3, Pkm, Hpca, Tnf, Hk2 | 5 | ||||
| mmu05205 | Proteoglycans in cancer | 205 | 0.03 | Col1a1, Fasl, Sdc4, Fas, Plaur, Met, Cd44, Hbegf, Tlr2 | 0.01 | Hif1a, Plau, Fzd7, Hgf, Myc, Hpse, Il12b, Stat3, Itga5, Cdkn1a, Tlr4, Tnf | 21 | ||
| mmu05418 | Fluid shear stress and atherosclerosis | 148 | 9.91 × 10−3 | Il1a, Jun, Tnfrsf1a, Il1b, Icam1, Il1r2, Fos, Mgst1, Tnf, Ass1 | 0.02 | Txn1, Mapk13, Ccl12, Ifng, Ccl2 | 15 | ||
| mmu04216 | Ferroptosis | 40 | 9.54 × 10−3 | Fth1, Slc39a14, Slc39a1, Cp | 4 | ||||
| mmu05132 | Salmonella infection | 253 | 9.31 × 10−4 | Jun, Pycard, Tubb6, Cycs, Nlrp3, Fos, Nod1, Gapdh, Myd88, Tnf, Tnfsf10, Tuba8, Tnfrsf1a, Ripk3, Il1b, Myc, Tlr4 | 0.01 | Txn1, Casp7, Mapk13, Il6, Mlkl, Tubb3, Bak1 | 24 | ||
| mmu05166 | Human T-cell leukemia virus 1 infection | 247 | 7.53 × 10−6 | H2-T23, Jun, H2-T10, H2-Q6, H2-Q7, Il2ra, H2-Q2, Fos, Creb5, Il15ra, Tnf, Tnfrsf1a, Gm11127, Myc, Icam1, Il1r2, Tspo, Cdkn2a, Cdkn1a, Csf2, B2m | 21 | ||||
| mmu01200 | Carbon metabolism | 122 | 2.55 × 10−3 | Pkm, Tpi1, Got1, Eno1b, Pgk1, Pgam1, Eno1, Gapdh, Hk2, Pfkp | 10 | ||||
| mmu05210 | Colorectal cancer | 88 | 0.01 | Jun, Cycs, Myc, Mcub, Fos, Cdkn1a, Ralgds | 7 | ||||
| mmu05168 | Herpes simplex virus 1 infection | 458 | 0.02 | H2-T23, Ifih1, Socs3, H2-T10, Cycs, H2-Q6, H2-Q7, Il12b, Tap2, H2-Q2, Itga5, Myd88, Tnf, Bst2, C3, Tnfrsf1a, Gm11127, Il1b, Tapbp, B2m, Daxx | 4.15 × 10−5 | H2-T22, Eif2ak2, Tap1, Oas1g, Il6, Ccl12, Ifng, Oas2, Cgas, Oas1a, Irf7, Ccl2, Stat2, Bak1, Stat1 | 36 | ||
| mmu04664 | Fc epsilon RI signaling pathway | 66 | 0.04 | Fcer1g, Il13, Pla2g4a, Csf2, Tnf | 5 | ||||
| mmu01524 | Platinum drug resistance | 80 | 0.03 | Slc31a1, Cycs, Cdkn2a, Mgst1, Cdkn1a, Atp7a | 6 | ||||
| mmu04978 | Mineral absorption | 54 | 0.03 | Slc6a19, Fth1, Slc39a1, Steap2 | 4 | ||||
| mmu04931 | Insulin resistance | 110 | 0.01 | Ptpn1, Socs3, Tnfrsf1a, Nr1h3, Stat3, Ppargc1b, Creb5, Tnf | 8 | ||||
| mmu05231 | Choline metabolism in cancer | 98 | 0.02 | Hif1a, Jun, Slc44a5, Pdgfc, Pla2g4a, Fos, Ralgds | 7 | ||||
| mmu05221 | Acute myeloid leukemia | 70 | 0.05 | Itgam, Myc, Stat3, Cebpe, Csf2 | 5 | ||||
| mmu04146 | Peroxisome | 86 | 0.04 | Prdx5, Nos2, Prdx1, Hp, Sod2, Xdh | 6 | ||||
| mmu04932 | Non-alcoholic fatty liver disease | 151 | 0.01 | Il1a, Jun, Socs3, Tnfrsf1a, Cycs, Il1b, Nr1h3, Cox4i2, Fos, Tnf | 10 | ||||
| mmu05222 | Small cell lung cancer | 93 | 0.05 | Nos2, Col4a1, Cycs, Myc, Lamc2, Cdkn1a | 6 | ||||
| mmu00052 | Galactose metabolism | 32 | 0.04 | Hk3, Mgam | 2 | ||||
| mmu04215 | Apoptosis—multiple species | 32 | 0.04 | Casp7, Bak1 | 2 | ||||
| mmu04010 | MAPK signaling pathway | 294 | 1.91 × 10−3 | Jun, Cacna1f, Hpca, Hgf, Pdgfc, Mcub, Hspa1b, Hspa1a, Pla2g4a, Fos, Myd88, Tnf, Hspa8, Il1a, Tnfrsf1a, Il1b, Myc, Daxx | 18 | ||||
| mmu05322 | Systemic lupus erythematosus | 148 | 0.03 | C1qb, C3, Il10, C1ra, C1s2, C1s1, Tnf, Elane, Trim21 | 9 | ||||
| mmu04916 | Melanogenesis | 100 | 0.02 | AC117663.3, Sco1, Creb3l3, Creb3l1, Mitf, AC110211.1 | 6 | ||||
| mmu04218 | Cellular senescence | 184 | 0.02 | H2-T23, Il1a, H2-T10, Gm11127, H2-Q6, Myc, H2-Q7, H2-Q2, Serpine1, Cdkn2a, Cdkn1a | 11 | ||||
| mmu05020 | Prion disease | 268 | 4.31 × 10−3 | C1qb, Cacna1f, Tubb6, Cycs, Psmb2, Hspa1b, Hspa1a, Creb5, Tnf, Tuba8, Hspa8, Il1a, Cybb, Il1b, Cox4i2, Psma8 | 16 | ||||
| mmu00500 | Starch and sucrose metabolism | 34 | 0.05 | Hk3, Mgam | 2 | ||||
| mmu04917 | Prolactin signaling pathway | 74 | 6.29 × 10−3 | Socs1, Mapk13, Irf1, Stat1 | 4 | ||||
| mmu04142 | Lysosome | 131 | 0.02 | Slc11a1, Ctss, Ctsd, Npc2, Lamp2, Ctsz, Lgmn | 7 | ||||
| mmu04151 | PI3K-Akt signaling pathway | 359 | 0.03 | Col4a1, Hgf, Pdgfc, Mcub, Il2ra, Tnc, Itga5, Creb5, Ywhag, Myc, Gnb4, Pik3r6, Lamc2, Cdkn1a, Csf3, Tlr4, Thbs4 | 17 | ||||
| mmu04926 | Relaxin signaling pathway | 129 | 0.05 | Col3a1, Col1a1, Col4a2, Creb3l3, Creb3l1, Gng12 | 6 | ||||
| mmu04622 | RIG-I-like receptor signaling pathway | 70 | 0.03 | Mapk13, Cxcl10, Irf7 | 3 | ||||
| mmu05200 | Pathways in cancer | 543 | 0.04 | Jun, Il12rb2, Nos2, Col4a1, Cycs, Fzd7, Hgf, Il13, Il12b, Il2ra, Fos, Il15ra, Ralgds, Csf2rb2, Hif1a, Csf2rb, Myc, Gnb4, Stat3, Cdkn2a, Lamc2, Mgst1, Cdkn1a | 23 | ||||
| mmu05202 | Transcriptional misregulation in cancer | 223 | 0.05 | Gadd45g, Gadd45b, Mitf, Plat, Cd14, Met, Nr4a3, Bcl2a1b, Bcl2a1a | 9 | ||||
| KEGG Pathway ID | KEGG Pathway Name | Total Known Genes | ΔclpB Alone | ΔclpB and LVS Shared | ΔclpB, LVS and ΔgplX Shared | Total Gene Counts a | |||
|---|---|---|---|---|---|---|---|---|---|
| padj | Matched Genes | padj | Matched Genes | padj | Matched Genes | ||||
| mmu04080 | Neuroactive ligand–receptor interaction | 358 | 2.69 × 10−5 | Gh, Ghrhr, Chrne, Gabrr2, Npff, Cnr1, Prss2, Lhcgr, Cort, Tshr, Gabra4, Vipr2, Gria2, Gal, Gria1, Sstr2, Chrna6, Grm6, P2rx2, Glp1r, Grik3, Tac2, S1pr5, Htr5b | 1.08 × 10−3 | Grin2a, Npb, Adra1b, Oxtr, Adra2b, Edn3, Ednrb, Npy1r, Aplnr, Gpr156, Vipr1, Ptgfr, Grm4, Gabbr1, Rxfp1, Lpar3 | 40 | ||
| mmu04020 | Calcium signaling pathway | 240 | 6.53 × 10−4 | Cacna1g, Atp2b2, Egf, Atp2a3, Fgf18, Plce1, Casq2, Fgfr4, Lhcgr, Plcd3, Camk2b, Pln, P2rx2, Adcy2, Mylk3, Htr5b | 1.59 × 10−4 | Grin2a, Ryr2, Adra1b, Oxtr, Ednrb, Cacna1b, Fgfr3, Fgfr2, Prkcg, Ntrk2, Ptgfr, Camk2a, Ntrk3, Cacna1i | 30 | ||
| mmu05414 | Dilated cardiomyopathy | 94 | 8.52 × 10−4 | Tro, Tnnt2, Pln, Sgca, Atp2a3, Itga8, Adcy2, Sgcg, Ttn | 8.30 × 10−3 | Ryr2, Itga11, Tgfb2, Adcy5, Cacna2d4, Cacng3 | 15 | ||
| mmu04512 | ECM–receptor interaction | 88 | 8.81 × 10−3 | Frem2, Vtn, Reln, Tnxb, Col4a3, Col6a6, Itga8 | 1.27 × 10−3 | Itga11, Sv2b, Sv2a, Lama3, Col6a4, Fras1, Thbs3 | 14 | ||
| mmu05410 | Hypertrophic cardiomyopathy | 91 | 2.83 × 10−3 | Tro, Tnnt2, Sgca, Atp2a3, Prkag3, Itga8, Sgcg, Ttn | 7.12 × 10−3 | Ryr2, Itga11, Ace, Tgfb2, Cacna2d4, Cacng3 | 14 | ||
| mmu04916 | Melanogenesis | 100 | 0.02 | Camk2b, Fzd2, Gnao1, Hr, Adcy2, Wnt2, AC084822.1 | 0.01 | Prkcg, Ednrb, Camk2a, Fzd6, Wnt5b, Adcy5 | 13 | ||
| mmu05217 | Basal cell carcinoma | 63 | 0.03 | Fzd8, Gli1, Wnt10a, Wnt10b | 0.03 | Bmp4, Apc2, Fzd6, Wnt5b | 8 | ||
| mmu04950 | Maturity onset diabetes of the young | 27 | 6.39 × 10−4 | Bhlha15, Hnf1b, Nr5a2, Foxa2, Foxa3 | 5 | ||||
| mmu01100 | Metabolic pathways | 1573 | 9.01 × 10−4 | Rimkla, Aldh1a1, Pcx, Selenbp2, Gsta4, Ptdss2, Sec1, B3gnt3, Uros, Mgat3, Suox, Phospho1, Pcyt1b, Cers1, Dhtkd1, B4galnt3, Cox6a2, Hmbs, Mgst3, Hsd3b1, Hagh, Adcy1, Car8, Nags, Mgll, Nqo1, Car2, Gpx1, St3gal5, Pigq, Pik3c2b, Aspdh, Cel, Gck, Cox6b2, Cox8b, Fahd1, Hyal3, Pipox, Urod, Mboat2, Pnpo, Sgpp2, Pip5k1b, Acmsd, Trak2 | 46 | ||||
| mmu04260 | Cardiac muscle contraction | 87 | 1.19 × 10−3 | Myl4, Actc1, Cox6b2, Cox8b, Cox6a2, Cacng4, Trdn | 7 | ||||
| mmu00260 | Glycine, serine and threonine metabolism | 40 | 3.94 × 10−3 | Gamt, Alas2, Cbs, Gcat, Gnmt | 5 | ||||
| mmu04310 | Wnt signaling pathway | 168 | 4.27 × 10−3 | Apc2, Prkcg, Tle2, Camk2a, Fzd6, Sox17, Wnt5b, Cxxc4, Dkk2 | 9 | ||||
| mmu04514 | Cell adhesion molecules | 174 | 5.37 × 10−3 | Cldn13, Cadm3, Cd4, Cdh4, Cldn9, H2-M2, Cd8b1, Nrxn2, Vtcn1 | 9 | ||||
| mmu04722 | Neurotrophin signaling pathway | 121 | 7.58 × 10−3 | Mapk12, Ntrk2, Camk2a, Ntrk3, Ntf3, Mapk11, Matk | 7 | ||||
| mmu05412 | Arrhythmogenic right ventricular cardiomyopathy | 77 | 0.02 | Actn2, Cdh2, Sgca, Atp2a3, Itga8, Sgcg | 6 | ||||
| mmu00860 | Porphyrin and chlorophyll metabolism | 43 | 0.05 | Hmbs, Uros, Urod | 3 | ||||
| mmu04360 | Axon guidance | 181 | 0.05 | Rac3, Syp, Efna4, Plxnb1, Prkcz, Sema4f, Myl9 | 0.05 | Epha4, Ablim2, Ephb6, Camk2b, Epha8, Hr, Ephb1, Bmp7, Sema4g | 6.92 × 10−3 | Efnb2, Camk2a, Ablim3, Sema6c, Wnt5b, Ntn4, Lrrc4c, L1cam, Rgma | 25 |
| mmu04024 | cAMP signaling pathway | 215 | 1.58×10−5 | Atp2b2, Npr1, Hhip, Atp2a3, Plce1, Lhcgr, Cnga2, Tshr, Cnga1, Vipr2, Ppp1r1b, Camk2b, Gria2, Gria1, Pln, Sstr2, Glp1r, Adcy2 | 0.02 | Grin2a, Ryr2, Oxtr, Edn3, Gabbr1, Camk2a, Fxyd1, Npy1r, Adcy5 | 27 | ||
| mmu04972 | Pancreatic secretion | 114 | 2.77 × 10−4 | Car2, Cckar, Cela3a, Prss1, Slc12a2, Adcy1, Cel, Slc26a3, Cpa1 | 0.01 | Atp2b2, Atp2a3, Kcnq1, Adcy2, Amy1, Ctrl, Prss2, Cpa2 | 17 | ||
| mmu04713 | Circadian entrainment | 98 | 0.05 | Cacna1g, Camk2b, Gria2, Gria1, Gnao1, Adcy2 | 8.67 × 10−5 | Grin2a, Ryr2, Gucy1a1, Prkcg, Camk2a, Gng8, Kcnj9, Adcy5, Cacna1i | 15 | ||
| mmu04974 | Protein digestion and absorption | 108 | 0.02 | Cela3a, Prss1, Col14a1, Col4a6, Col8a2, Cpa1 | 5.78×10−4 | Col11a1, Col4a3, Col13a1, Eln, Col6a6, Kcnq1, Ctrl, Col19a1, Prss2, Cpa2 | 16 | ||
| mmu04727 | GABAergic synapse | 89 | 2.46×10−3 | Gabra4, Gls2, Slc12a5, Gabrr2, Gnao1, Slc38a3, Abat, Adcy2 | 6.40 × 10−3 | Prkcg, Gabbr1, Gad1, Cacna1b, Gng8, Adcy5 | 14 | ||
| mmu04724 | Glutamatergic synapse | 113 | 8.26 × 10−4 | Shank1, Gria2, Gria1, Homer2, Gls2, Grm6, Gnao1, Slc38a3, Adcy2, Grik3 | 0.02 | Grin2a, Pla2g4f, Prkcg, Grm4, Gng8, Adcy5 | 16 | ||
| mmu04971 | Gastric acid secretion | 75 | 0.01 | Atp4a, Camk2b, Sstr2, Kcnq1, Adcy2, Mylk3 | 2.75 × 10−3 | Prkcg, Kcnj1, Camk2a, Kcnj16, Adcy5, Kcnf1 | 12 | ||
| mmu04911 | Insulin secretion | 86 | 0.02 | Cckar, Kcnn2, Adcy1, Gcg, Gck | 5.42 × 10−3 | Ryr2, Prkcg, Syt3, Camk2a, Ffar1, Adcy5 | 11 | ||
| mmu05200 | Pathways in cancer | 543 | 0.05 | Nqo1, Fbxo24, Gsta4, Ctnna3, Fzd8, Flt3l, Col4a6, Gli1, Mgst3, Notch3, Gnb3, Rac3, Adcy1, Wnt10a, Hes5, Wnt10b | 2.78 × 10−3 | Apc2, Hlf, Ednrb, Fzd6, Runx1t1, Fgfr3, Fgfr2, Bmp4, Prkcg, Heyl, Tgfb2, Hey2, Camk2a, Lama3, Hey1, Wnt5b, Gng8, Rxrg, Lpar3, Adcy5 | 36 | ||
| mmu04925 | Aldosterone synthesis and secretion | 102 | 0.05 | Cacna1g, Camk2b, Atp2b2, Npr1, Star, Adcy2 | 2.98 × 10−3 | Hsd3b6, Kcnk3, Prkcg, Cyp21a1, Camk2a, Adcy5, Cacna1i | 13 | ||
| mmu04261 | Adrenergic signaling in cardiomyocytes | 152 | 0.05 | Tro, Tnnt2, Camk2b, Pln, Atp2b2, Atp2a3, Kcnq1, Adcy2 | 7.70 × 10−3 | Mapk12, Ryr2, Adra1b, Camk2a, Adcy5, Cacna2d4, Cacng3, Mapk11 | 16 | ||
| mmu04725 | Cholinergic synapse | 112 | 0.03 | Ache, Camk2b, Chrna6, Gnao1, Kcnq1, Hr, Adcy2 | 0.02 | Prkcg, Camk2a, Cacna1b, Gng8, Adcy5, Kcnf1 | 13 | ||
| mmu05031 | Amphetamine addiction | 69 | 0.04 | Ppp1r1b, Camk2b, Ddc, Gria2, Gria1 | 0.04 | Grin2a, Prkcg, Camk2a, Adcy5 | 9 | ||
| mmu05231 | Choline metabolism in cancer | 98 | 0.04 | Pcyt1b, Slc22a2, Wasf3, Rac3, Pip5k1b | 0.05 | Gpcpd1, Egf, Slc22a4, Dgkb, Hr, Chkb | 11 | ||
| mmu04350 | TGF-beta signaling pathway | 95 | 1.0 × 10−45 | Bmp4, Tgfb2, 4930516B21Rik, Nog, Id4, Smad9, Id3, Fmod, Rgma, Thsd4 | 10 | ||||
| mmu04550 | Signaling pathways regulating pluripotency of stem cells | 140 | 1.15 × 10−5 | Bmp4, Mapk12, Apc2, 4930516B21Rik, Fzd6, Wnt5b, Id4, Smad9, Id3, Fgfr3, Fgfr2, Mapk11 | 12 | ||||
| mmu04921 | Oxytocin signaling pathway | 153 | 1.35 × 10−4 | Ryr2, Gucy1a1, Pla2g4f, Prkcg, Oxtr, Camk2a, Kcnj9, Adcy5, Kcnf1, Cacna2d4, Cacng3 | 11 | ||||
| mmu04976 | Bile secretion | 100 | 5.54 × 10−4 | Car2, Kcnn2, Aqp9, Ephx1, Aqp8, Adcy1, Slc22a7, Aqp1 | 8 | ||||
| mmu04728 | Dopaminergic synapse | 135 | 9.55 × 10−4 | Grin2a, Mapk12, Prkcg, Camk2a, Cacna1b, Gng8, Kcnj9, Adcy5, Mapk11 | 9 | ||||
| mmu04640 | Hematopoietic cell lineage | 94 | 1.87 × 10−3 | Cd24a, Cd4, Cd59b, Tfrc, Cd8b1, Dntt, Flt3l | 7 | ||||
| mmu04934 | Cushing syndrome | 162 | 3.36 × 10−3 | Apc2, Hsd3b6, Kcnk3, Cyp21a1, Camk2a, Fzd6, Wnt5b, Adcy5, Cacna1i | 9 | ||||
| mmu04010 | MAPK signaling pathway | 294 | 3.48 × 10−3 | Mapk12, Pla2g4f, Cacna1b, Fgfr3, Cacng3, Fgfr2, Prkcg, Ntrk2, Tgfb2, Ntf3, Cacna1i, Cacna2d4, Mapk11 | 13 | ||||
| mmu05144 | Malaria | 57 | 3.89 × 10−3 | Gypa, Hbb-bh2, Hba-a1, Hbb-bt, Hbb-bs, Ackr1 | 6 | ||||
| mmu05033 | Nicotine addiction | 40 | 3.94 × 10−3 | Gabra4, Gria2, Gria1, Chrna6, Gabrr2 | 5 | ||||
| mmu00350 | Tyrosine metabolism | 40 | 6.45 × 10−3 | Aoc3, Adh1, Aox4, Aox3 | 4 | ||||
| mmu04723 | Retrograde endocannabinoid signaling | 148 | 6.59 × 10−3 | Mapk12, Prkcg, Faah, Cacna1b, Gng8, Kcnj9, Adcy5, Mapk11 | 8 | ||||
| mmu05032 | Morphine addiction | 91 | 7.12 × 10−3 | Prkcg, Gabbr1, Cacna1b, Gng8, Kcnj9, Adcy5 | 6 | ||||
| mmu00514 | Other types of O-glycan biosynthesis | 43 | 8.33 × 10−3 | St6gal1, Colgalt2, Gxylt2, Galnt16 | 4 | ||||
| mmu00410 | beta-Alanine metabolism | 31 | 8.76 × 10−3 | Upb1, Aldh3a1, Aldh3b2, Abat | 4 | ||||
| mmu00750 | Vitamin B6 metabolism | 9 | 0.01 | Aox4, Aox3 | 2 | ||||
| mmu04924 | Renin secretion | 76 | 0.01 | Gucy1a1, Ace, Edn3, Adcy5, Kcnf1 | 5 | ||||
| mmu04927 | Cortisol synthesis and secretion | 72 | 0.01 | Hsd3b6, Kcnk3, Cyp21a1, Adcy5, Cacna1i | 5 | ||||
| mmu00360 | Phenylalanine metabolism | 23 | 0.02 | Ddc, Aldh3a1, Aldh3b2 | 3 | ||||
| mmu00920 | Sulfur metabolism | 11 | 0.02 | Selenbp2, Suox | 2 | ||||
| mmu04977 | Vitamin digestion and absorption | 24 | 0.02 | Slc23a1, Apoa4, Plb1 | 3 | ||||
| mmu05143 | African trypanosomiasis | 39 | 0.02 | Hbb-bh2, Hba-a1, Hbb-bt, Hbb-bs | 4 | ||||
| mmu04015 | Rap1 signaling pathway | 214 | 0.02 | Grin2a, Mapk12, Prkcg, Magi2, Fgfr3, Lpar3, Adcy5, Fgfr2, Mapk11 | 9 | ||||
| mmu04072 | Phospholipase D signaling pathway | 149 | 0.02 | Pla2g4f, Ptgfr, Grm4, Dgka, Lpar3, Adcy5, Dnm1 | 7 | ||||
| mmu04270 | Vascular smooth muscle contraction | 143 | 0.02 | Gucy1a1, Pla2g2d, Pla2g4f, Prkcg, Adra1b, Edn3, Adcy5 | 7 | ||||
| mmu04370 | VEGF signaling pathway | 58 | 0.02 | Mapk12, Pla2g4f, Prkcg, Mapk11 | 4 | ||||
| mmu04970 | Salivary secretion | 86 | 0.02 | Gucy1a1, Prkcg, Adra1b, Trpv6, Adcy5 | 5 | ||||
| mmu05152 | Tuberculosis | 180 | 0.02 | Cd209g, Mapk12, Cd209f, Tgfb2, Camk2a, Cd209a, Atp6v0a4, Mapk11 | 8 | ||||
| mmu05418 | Fluid shear stress and atherosclerosis | 148 | 0.02 | Bmp4, Mapk12, Thbd, 4930516B21Rik, Klf2, Mapk11, Nox1 | 7 | ||||
| mmu04150 | mTOR signaling pathway | 156 | 0.03 | Rps6ka6, Stradb, Deptor, Fbxo24, Fzd8, Wnt10a, Wnt10b | 7 | ||||
| mmu04014 | Ras signaling pathway | 232 | 0.03 | Grin2a, Pla2g2d, Pla2g4f, Prkcg, Ntrk2, Ntf3, Gng8, Fgfr3, Fgfr2 | 9 | ||||
| mmu04330 | Notch signaling pathway | 60 | 0.03 | Heyl, Tle2, Hey2, Hey1 | 4 | ||||
| mmu04390 | Hippo signaling pathway | 157 | 0.03 | Bmp4, Apc2, Tgfb2, Rassf6, Fzd6, Wnt5b, Ajuba | 7 | ||||
| mmu04750 | Inflammatory mediator regulation of TRP channels | 127 | 0.03 | Mapk12, Pla2g4f, Prkcg, Camk2a, Adcy5, Mapk11 | 6 | ||||
| mmu04912 | GnRH signaling pathway | 90 | 0.03 | Mapk12, Pla2g4f, Camk2a, Adcy5, Mapk11 | 5 | ||||
| mmu04913 | Ovarian steroidogenesis | 63 | 0.03 | Hsd3b6, Pla2g4f, Cyp1a1, Adcy5 | 4 | ||||
| mmu04926 | Relaxin signaling pathway | 129 | 0.03 | Mapk12, Ednrb, Gng8, Rxfp1, Adcy5, Mapk11 | 6 | ||||
| mmu04929 | GnRH secretion | 63 | 0.03 | Prkcg, Gabbr1, Kcnj9, Cacna1i | 4 | ||||
| mmu04960 | Aldosterone-regulated sodium reabsorption | 38 | 0.03 | Prkcg, Kcnj1, Sfn | 3 | ||||
| mmu00910 | Nitrogen metabolism | 17 | 0.04 | Car2, Car8 | 2 | ||||
| mmu04710 | Circadian rhythm | 30 | 0.04 | Npas2, Prkag3, Rorc | 3 | ||||
| mmu05135 | Yersinia infection | 134 | 0.04 | Cd4, Rps6ka6, Fbxo24, Cd8b1, Rac3, Pip5k1b | 6 | ||||
| mmu05218 | Melanoma | 72 | 0.04 | Egf, Fgf18, E2f2, Gadd45a, Hr | 5 | ||||
| mmu00250 | Alanine, aspartate and glutamate metabolism | 39 | 0.04 | Gad1, Aldh5a1, Ddo | 3 | ||||
| mmu00760 | Nicotinate and nicotinamide metabolism | 41 | 0.04 | Aox4, Aox3, Nmnat2 | 3 | ||||
| mmu00830 | Retinol metabolism | 97 | 0.04 | Adh1, Aox4, Aox3, Cyp1a1, Lrat | 5 | ||||
| mmu00982 | Drug metabolism—cytochrome P450 | 71 | 0.04 | Adh1, Aox4, Aox3, Fmo2 | 4 | ||||
| mmu04726 | Serotonergic synapse | 131 | 0.04 | Pla2g4f, Prkcg, Cacna1b, Gng8, Kcnj9, Adcy5 | 6 | ||||
| mmu04933 | AGE-RAGE signaling pathway in diabetic complications | 101 | 0.04 | Mapk12, Thbd, Tgfb2, Mapk11, Nox1 | 5 | ||||
| mmu05205 | Proteoglycans in cancer | 205 | 0.04 | Mapk12, Prkcg, Tgfb2, Camk2a, Fzd6, Wnt5b, Gpc3, Mapk11 | 8 | ||||
| mmu00380 | Tryptophan metabolism | 52 | 0.05 | Afmid, Ddc, Haao, Inmt | 4 | ||||
| mmu04918 | Thyroid hormone synthesis | 74 | 0.05 | Ttr, Adcy2, Duox2, Slc5a5, Tshr | 5 | ||||
| mmu05214 | Glioma | 74 | 0.05 | Camk2b, Egf, E2f2, Gadd45a, Hr | 5 | ||||
| mmu05225 | Hepatocellular carcinoma | 174 | 0.05 | Nqo1, Gsta4, Mgst3, Dpf3, Fzd8, Wnt10a, Wnt10b | 7 | ||||
| Gene Name | Gene Description | Mean | Fold Change | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Naive | ΔclpB | LVS | ΔgplX | ΔlpcC | ΔclpB/ Naïve | ΔclpB/ LVS | ΔclpB/ ΔgplX | ΔclpB/ ΔlpcC | ||
| Acod1 | aconitate decarboxylase 1 | 29 | 3971 | 1364 | 253 | 31 | 137.8 | 2.9 | 15.7 | 125.7 |
| Saa3 | serum amyloid A 3 | 47 | 5752 | 1000 | 45 | 21 | 122.2 | 5.8 | 128.1 | 282.4 |
| Ccl2 | chemokine (C-C motif) ligand 2 | 36 | 1606 | 685 | 109 | 65 | 44.7 | 2.3 | 14.9 | 24.8 |
| Clec4e | C-type lectin domain family 4, member e | 83 | 3073 | 978 | 208 | 105 | 37.1 | 3.1 | 14.8 | 29.2 |
| Timp1 | tissue inhibitor of metalloproteinase 1 | 90 | 3142 | 1181 | 174 | 95 | 35.0 | 2.7 | 18.1 | 33.2 |
| Serpine1 | serine (or cysteine) peptidase inhibitor, clade E, member 1 | 46 | 1472 | 447 | 70 | 60 | 31.8 | 3.3 | 21.1 | 24.7 |
| Mmp3 | matrix metallopeptidase 3 | 63 | 1862 | 371 | 111 | 59 | 29.8 | 5.0 | 16.8 | 31.7 |
| Inhba | inhibin beta-A | 34 | 858 | 273 | 36 | 24 | 25.2 | 3.1 | 24.0 | 35.1 |
| Cxcl2 | chemokine (C-X-C motif) ligand 2 | 13 | 314 | 63 | 12 | 13 | 24.5 | 5.0 | 25.4 | 23.9 |
| Il1rn | interleukin 1 receptor antagonist | 175 | 4269 | 1744 | 389 | 168 | 24.4 | 2.4 | 11.0 | 25.4 |
| Cxcl1 | chemokine (C-X-C motif) ligand 1 | 29 | 600 | 221 | 45 | 26 | 20.8 | 2.7 | 13.6 | 22.8 |
| Vcan | versican | 45 | 893 | 267 | 95 | 35 | 19.9 | 3.3 | 9.5 | 25.6 |
| Adamts4 | a disintegrin-like and metallopeptidase (reprolysin type) with thrombospondin type 1 motif, 4 | 23 | 415 | 130 | 33 | 25 | 18.2 | 3.2 | 12.6 | 16.9 |
| Cxcl5 | chemokine (C-X-C motif) ligand 5 | 54 | 961 | 465 | 96 | 70 | 17.8 | 2.1 | 10.0 | 13.9 |
| Il1r2 | interleukin 1 receptor, type II | 45 | 749 | 227 | 87 | 37 | 16.7 | 3.3 | 8.6 | 20.3 |
| Lipg | lipase, endothelial | 42 | 610 | 248 | 90 | 50 | 14.5 | 2.5 | 6.8 | 12.2 |
| Mmp8 | matrix metallopeptidase 8 | 172 | 2266 | 465 | 314 | 116 | 13.2 | 4.9 | 7.2 | 19.6 |
| Oas1g | 2′-5′ oligoadenylate synthetase 1G | 78 | 884 | 423 | 218 | 86 | 11.2 | 2.1 | 4.1 | 10.3 |
| Gzmb | granzyme B | 183 | 1943 | 803 | 497 | 291 | 10.6 | 2.4 | 3.9 | 6.7 |
| Chil1 | chitinase-like 1 | 96 | 1003 | 370 | 196 | 90 | 10.4 | 2.7 | 5.1 | 11.2 |
| Lox | lysyl oxidase | 95 | 905 | 189 | 47 | 62 | 9.5 | 4.8 | 19.2 | 14.6 |
| Il1a | interleukin 1 alpha | 145 | 1291 | 496 | 164 | 151 | 8.9 | 2.6 | 7.9 | 8.5 |
| Ccl3 | chemokine (C-C motif) ligand 3 | 75 | 478 | 127 | 83 | 93 | 6.3 | 3.7 | 5.7 | 5.2 |
| Ccl4 | chemokine (C-C motif) ligand 4 | 102 | 585 | 208 | 130 | 174 | 5.7 | 2.8 | 4.5 | 3.4 |
| Hp | haptoglobin | 654 | 3737 | 1587 | 1119 | 580 | 5.7 | 2.4 | 3.3 | 6.4 |
| Il1b | interleukin 1 beta | 1089 | 6125 | 3006 | 1833 | 1115 | 5.6 | 2.0 | 3.3 | 5.5 |
| Il1f9 | interleukin 1 family, member 9 | 108 | 559 | 227 | 211 | 86 | 5.2 | 2.5 | 2.7 | 6.5 |
| Aqp1 | aquaporin 1 | 13520 | 3451 | 9079 | 30537 | 46957 | −3.9 | −2.6 | −8.9 | −13.6 |
| Sptb | spectrin beta, erythrocytic | 8903 | 1854 | 5303 | 21539 | 33398 | −4.8 | −2.9 | −11.6 | −18.0 |
| Art4 | ADP-ribosyltransferase 4 | 406 | 84 | 185 | 610 | 1184 | −4.8 | −2.2 | −7.3 | −14.1 |
| Slc6a9 | solute carrier family 6 (neurotransmitter transporter, glycine), member 9 | 648 | 87 | 210 | 1234 | 2130 | −7.5 | −2.4 | −14.2 | −24.6 |
| 1300017J02Rik | RIKEN cDNA 1300017J02 gene | 720 | 59 | 170 | 1114 | 2409 | −12.2 | −2.9 | −18.9 | −40.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.Y.; Leclerc, S.; Pan, Y.; Liu, Z.; Stark, F.; Conlan, J.W. Novel Transcriptional and Translational Biomarkers of Tularemia Vaccine Efficacy in a Mouse Inhalation Model: Proof of Concept. Microorganisms 2022, 10, 36. https://doi.org/10.3390/microorganisms10010036
Liu QY, Leclerc S, Pan Y, Liu Z, Stark F, Conlan JW. Novel Transcriptional and Translational Biomarkers of Tularemia Vaccine Efficacy in a Mouse Inhalation Model: Proof of Concept. Microorganisms. 2022; 10(1):36. https://doi.org/10.3390/microorganisms10010036
Chicago/Turabian StyleLiu, Qing Yan, Sonia Leclerc, Youlian Pan, Ziying Liu, Felicity Stark, and Joseph Wayne Conlan. 2022. "Novel Transcriptional and Translational Biomarkers of Tularemia Vaccine Efficacy in a Mouse Inhalation Model: Proof of Concept" Microorganisms 10, no. 1: 36. https://doi.org/10.3390/microorganisms10010036
APA StyleLiu, Q. Y., Leclerc, S., Pan, Y., Liu, Z., Stark, F., & Conlan, J. W. (2022). Novel Transcriptional and Translational Biomarkers of Tularemia Vaccine Efficacy in a Mouse Inhalation Model: Proof of Concept. Microorganisms, 10(1), 36. https://doi.org/10.3390/microorganisms10010036