
Biochemical Properties of Two Plasmodium malariae Cysteine Proteases, Malapain-2 and Malapain-4


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Cloning of Genes Encoding Malapains (MPs)
2.2. Expression, Purification, and Refolding of Recombinant MPs
2.3. Enzyme Activity Assay
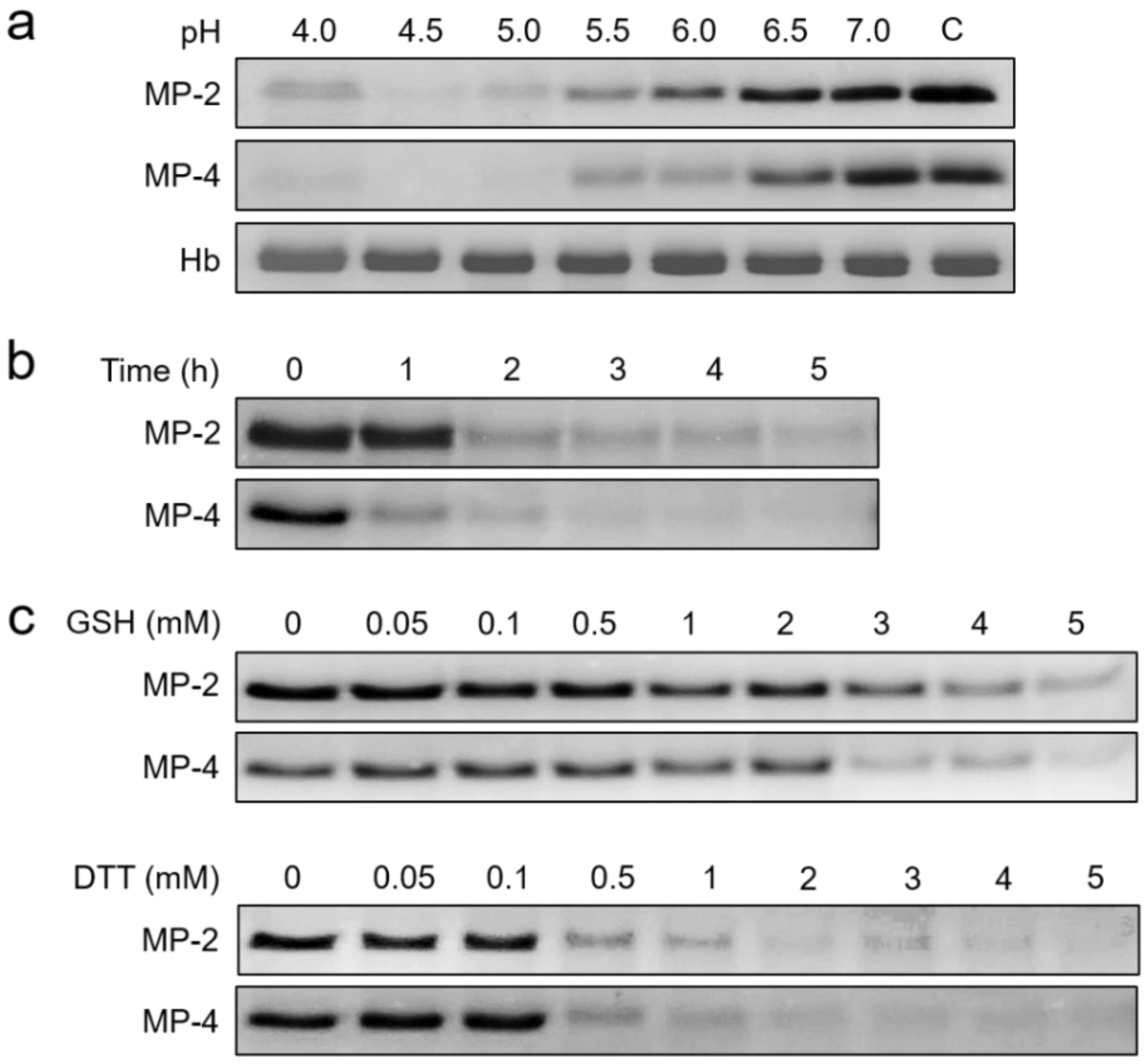
2.4. Biochemical Properties and Kinetics of MP-2 and MP-4
2.5. N-Terminal Amino Acid Sequencing
2.6. Hydrolysis of Human Hemoglobin
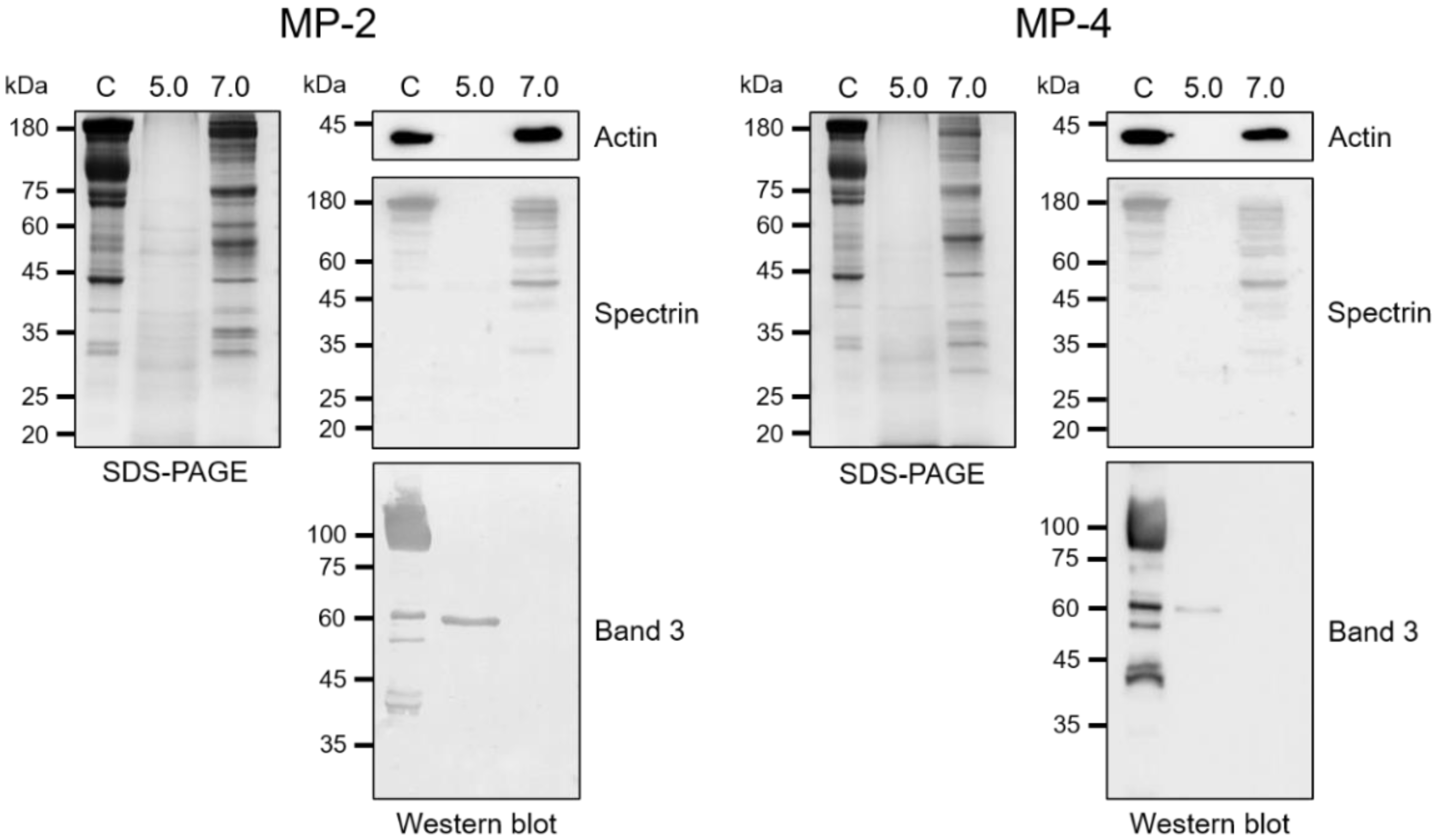
2.7. Hydrolysis of Erythrocyte Cytoskeletal Proteins
2.8. Statistical Analysis
3. Results
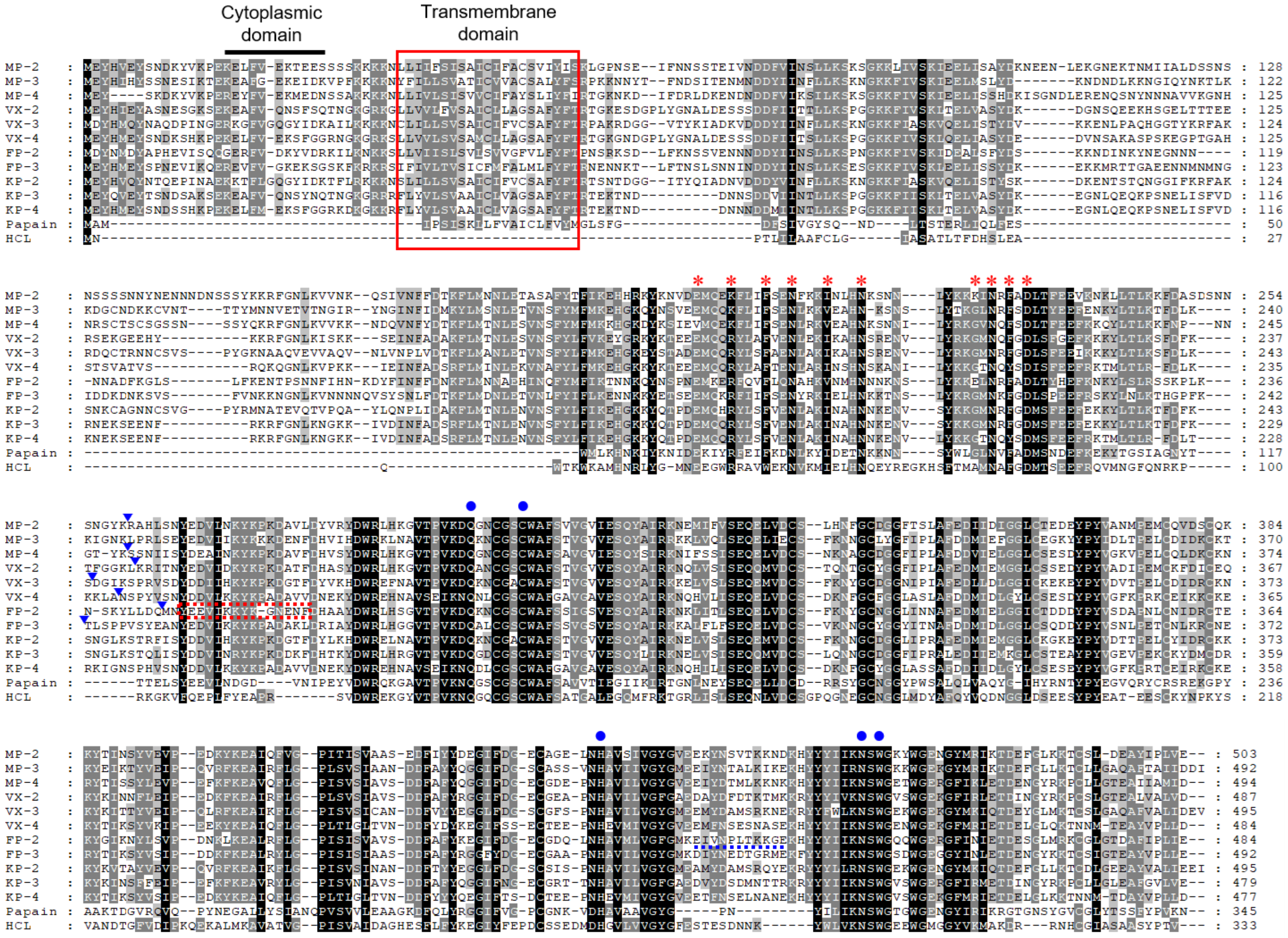
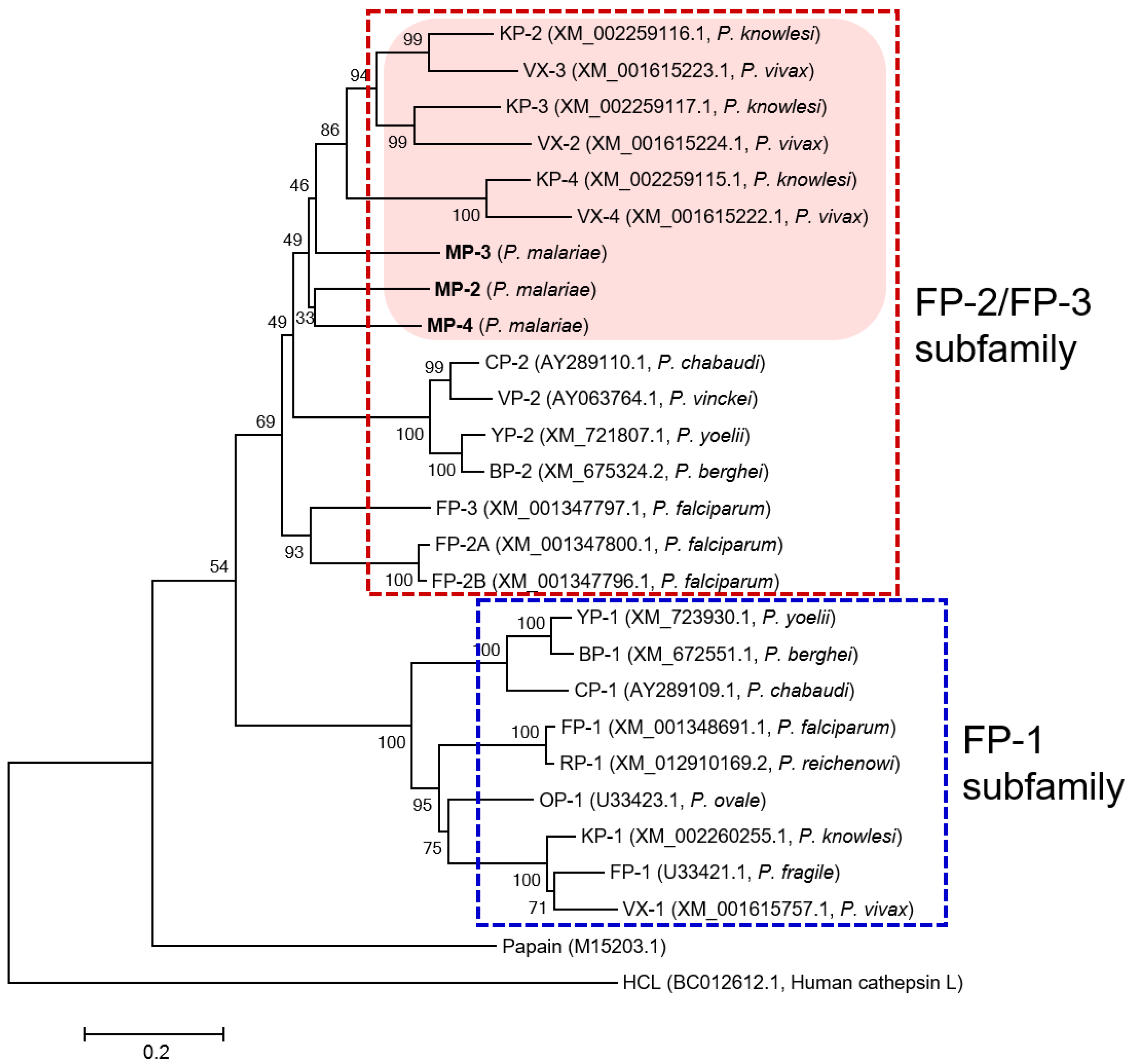
3.1. MPs Showed Close Relationship with VXs and KPs
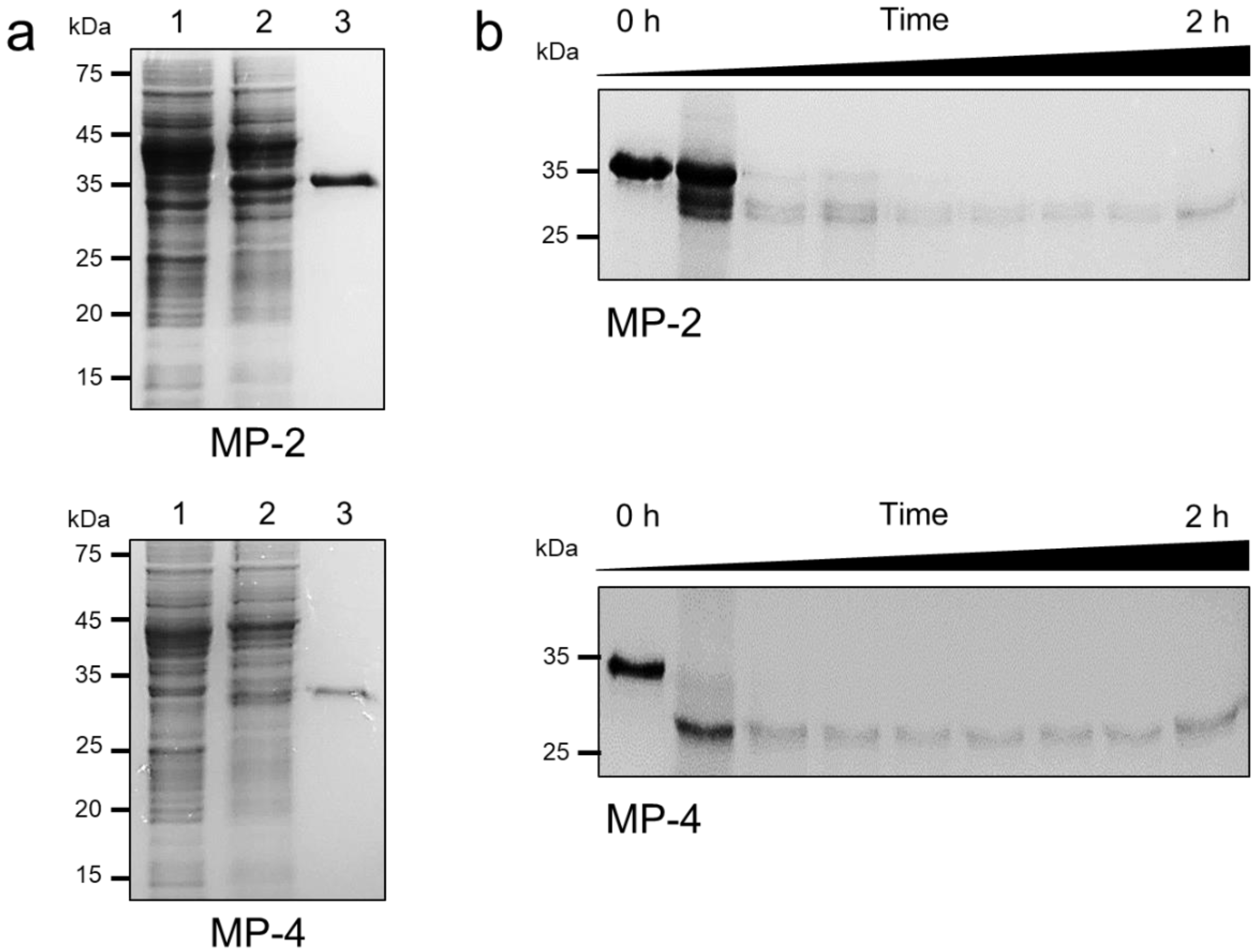
3.2. Recombinant MP-2 and MP-4 Were Successfully Produced by E. coli
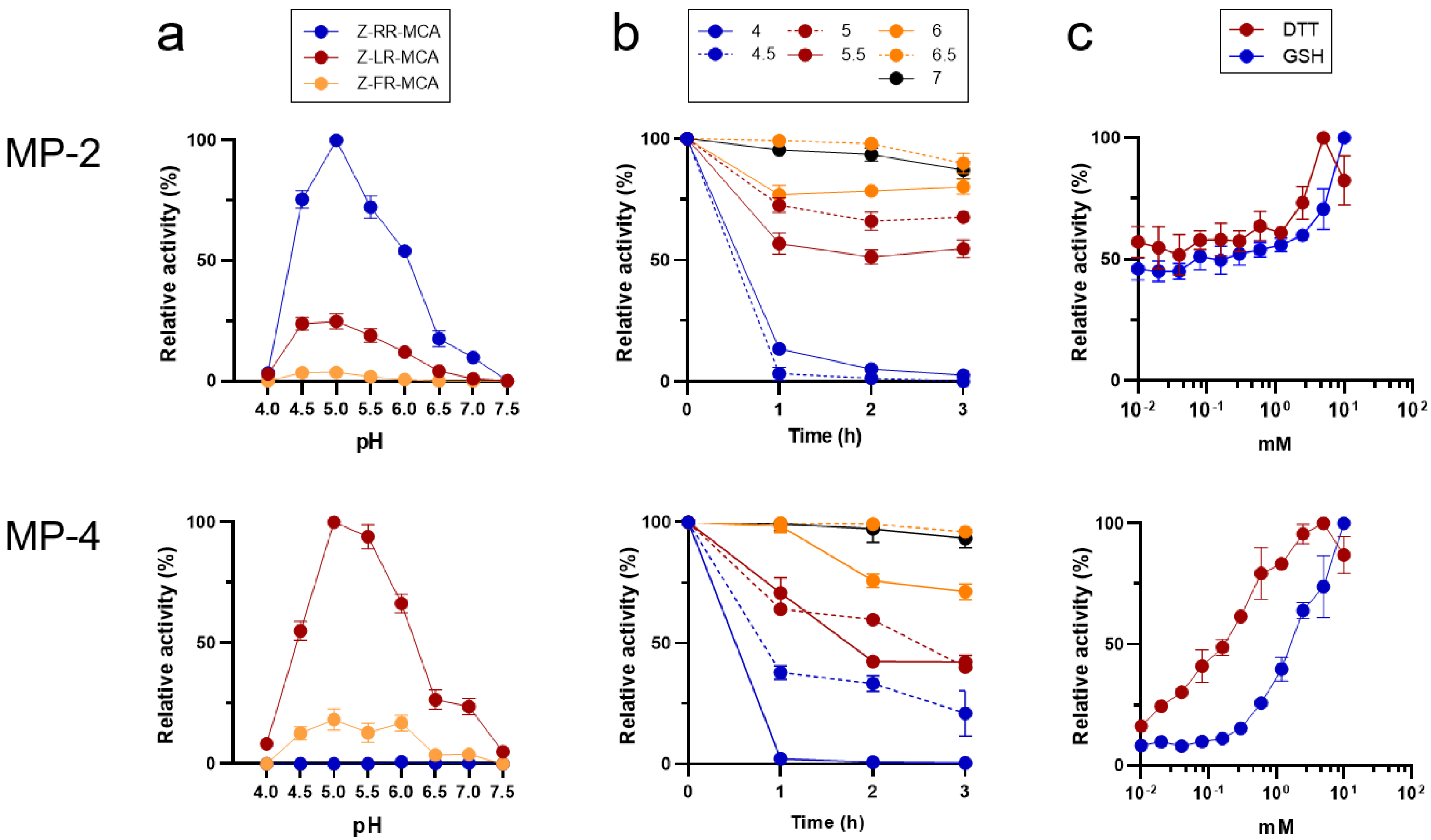
3.3. MP-2 and MP-4 Showed Different Preference for Peptide Substrates
3.4. MP-2 and MP-4 Hydrolyzed Hemoglobin and Erythrocyte Cytoskeletal Proteins
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rosenthal, P.J.; McKerrow, J.H.; Aikawa, M.; Nagasawa, H.; Leech, J.H. A malarial cysteine proteinase is necessary for hemoglobin degradation by Plasmodium falciparum. J. Clin. Investig. 1988, 82, 1560–1566. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.E.; Banerjee, R.; Goldberg, D.E. Biosynthesis and maturation of the malaria aspartic hemoglobinases plasmepsins I and II. J. Biol. Chem. 1997, 272, 14961–14968. [Google Scholar] [CrossRef] [PubMed]
- Sijwali, P.S.; Shenai, B.R.; Gut, J.; Singh, A.; Rosenthal, P.J. Expression and characterization of the Plasmodium falciparum haemoglobinase falcipain-3. Biochem. J. 2001, 360, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Shenai, B.R.; Sijwali, P.S.; Singh, A.; Rosenthal, P.J. Characterization of native and recombinant falcipain-2, a principal trophozoite cysteine protease and essential hemoglobinase of Plasmodium falciparum. J. Biol. Chem. 2000, 275, 29000–29010. [Google Scholar] [CrossRef] [PubMed]
- Salmon, B.L.; Oksman, A.; Goldberg, D.E. Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis. Proc. Natl. Acad. Sci. USA 2001, 98, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.C.; Baruch, A.; Grainger, M.; Bozdech, Z.; Medzihradszky, K.F.; Engel, J.; DeRisi, J.; Holder, A.A.; Bogyo, M. A role for the protease falcipain 1 in host cell invasion by the human malaria parasite. Science 2002, 298, 2002–2006. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, P.; Sijwali, P.; Singh, A.; Shenai, B. Cysteine Proteases of Malaria Parasites: Targets for Chemotherapy. Curr. Pharm. Des. 2005, 8, 1659–1672. [Google Scholar] [CrossRef] [PubMed]
- Wegscheid-Gerlach, C.; Gerber, H.-D.; Diederich, W. Proteases of Plasmodium falciparum as Potential Drug Targets and Inhibitors Thereof. Curr. Top. Med. Chem. 2010, 10, 346–367. [Google Scholar] [CrossRef]
- Bailly, E.; Jambou, R.; SaveL, J.; Jaureguiberry, G. Plasmodium falciparum: Differential Sensitivity In Vitro to E-64 (Cysteine Protease Inhibitor) and Pepstatin A (Aspartyl Protease Inhibitor). J. Protozool. 1992, 39, 593–599. [Google Scholar] [CrossRef]
- Rosenthal, P.J. Plasmodium falciparum: Effects of proteinase inhibitors on globin hydrolysis by cultured malaria parasites. Exp. Parasitol. 1995, 80, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Singh, V.; Singh, S. Structural insights into key Plasmodium proteases as therapeutic drug targets. Front. Microbiol. 2019, 10, 394. [Google Scholar] [CrossRef] [PubMed]
- Bekono, B.D.; Ntie-Kang, F.; Owono Owono, L.C.; Megnassan, E. Targeting Cysteine Proteases from Plasmodium falciparum: A General Overview, Rational Drug Design and Computational Approaches for Drug Discovery. Curr. Drug Targets 2018, 19, 501–526. [Google Scholar] [CrossRef] [PubMed]
- Perez, B.; Teixeira, C.; Gomes, J.; Gomes, P. Development of Plasmodium falciparum Protease Inhibitors in the Past Decade (2002–2012). Curr. Med. Chem. 2013, 20, 3049–3068. [Google Scholar] [CrossRef]
- Mayxay, M.; Pukrittayakamee, S.; Newton, P.N.; White, N.J. Mixed-species malaria infections in humans. Trends Parasitol. 2004, 20, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Kotepui, M.; Kotepui, K.U.; De Jesus Milanez, G.; Masangkay, F.R. Plasmodium spp. mixed infection leading to severe malaria: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 11068. [Google Scholar] [CrossRef] [PubMed]
- Na, B.K.; Shenai, B.R.; Sijwali, P.S.; Choe, Y.; Pandey, K.C.; Singh, A.; Craik, C.S.; Rosenthal, P.J. Identification and biochemical characterization of vivapains, cysteine proteases of the malaria parasite Plasmodium vivax. Biochem. J. 2004, 378, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Na, B.K.; Bae, Y.A.; Zo, Y.G.; Choe, Y.; Kim, S.H.; Desai, P.V.; Avery, M.A.; Craik, C.S.; Kim, T.S.; Rosenthal, P.J.; et al. Biochemical properties of a novel cysteine protease of Plasmodium vivax, vivapain-4. PLoS Negl. Trop. Dis. 2010, 4, 1–11. [Google Scholar] [CrossRef][Green Version]
- Prasad, R.; Atul; Soni, A.; Puri, S.K.; Sijwali, P.S. Expression, characterization, and cellular localization of knowpains, papain-like cysteine proteases of the Plasmodium knowlesi malaria parasite. PLoS ONE 2012, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vinetz, J.M.; Li, J.; McCutchan, T.F.; Kaslow, D.C. Plasmodium malariae Infection in an Asymptomatic 74-Year-Old Greek Woman with Splenomegaly. N. Engl. J. Med. 1998, 338, 367–371. [Google Scholar] [CrossRef]
- Douglas, N.M.; Lampah, D.A.; Kenangalem, E.; Simpson, J.A.; Poespoprodjo, J.R.; Sugiarto, P.; Anstey, N.M.; Price, R.N. Major Burden of Severe Anemia from Non-Falciparum Malaria Species in Southern Papua: A Hospital-Based Surveillance Study. PLoS Med. 2013, 10, e1001575. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.D.; Sumawinata, I.W.; Masbar, S.; Laksana, B.; Prodjodipuro, P.; Susanti, I.; Sismadi, P.; Mahmud, N.; Bangs, M.J.; Baird, J.K. Chloroquine-resistant Plasmodium malariae in south Sumatra, Indonesia. Lancet 2002, 360, 58–60. [Google Scholar] [CrossRef]
- Sijwali, P.S.; Brinen, L.S.; Rosenthal, P.J. Systematic optimization of expression and refolding of the Plasmodium falciparum cysteine protease falcipain-2. Protein Expr. Purif. 2001, 22, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J.; Kembhavi, A.A.; Brown, M.A.; Kirschke, H.; Knight, C.G.; Tamai, M.; Hanada, K. L-trans-Epoxysuccinyl-leucylamido(4-guanidino)butane (E-64) and its analogues as inhibitors of cysteine proteinases including cathepsins, B, H and L. Biochem. J. 1982, 201, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bennett, V.; Branton, D. Selective association of spectrin with the cytoplasmic surface of human erythrocyte plasma membranes. Quantitative determination with purified (32P)spectrin. J. Biol. Chem. 1977, 252, 2753–2763. [Google Scholar] [CrossRef]
- Rosenthal, P.J. Falcipains and other cysteine proteases of malaria parasites. Adv. Exp. Med. Biol. 2011, 712, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Shenai, B.R.; Choe, Y.; Gut, J.; Sijwali, P.S.; Craik, C.S.; Rosenthal, P.J. Critical role of amino acid 23 in mediating activity and specificity of vinckepain-2, a papain-family cysteine protease of rodent malaria parasites. Biochem. J. 2002, 368, 273–281. [Google Scholar] [CrossRef]
- Singh, A.; Walker, K.J.; Sijwali, P.S.; Lau, A.L.; Rosenthal, P.J. A chimeric cysteine protease of Plasmodium berghei engineered to resemble the Plasmodium falciparum protease falcipain-2. Protein Eng. Sel. 2007, 20, 171–177. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MP-3 | MP-4 | VX-2 | VX-3 | VX-4 | FP-2 | FP-3 | KP-2 | KP-3 | KP-4 | Papain | HCL | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MP-2 | 51.7 | 60.5 | 51.0 | 48.0 | 50.3 | 47.2 | 49.6 | 48.8 | 48.0 | 50.9 | 21.0 | 20.2 |
| MP-3 | – | 52.7 | 52.0 | 59.8 | 47.6 | 49.2 | 49.6 | 59.8 | 50.2 | 49.2 | 21.2 | 17.9 |
| MP-4 | – | – | 57.3 | 50.0 | 50.7 | 48.4 | 54.7 | 48.4 | 53.7 | 49.5 | 22.1 | 20.2 |
| VX-2 | – | – | – | 53.9 | 60.2 | 48.7 | 53.1 | 53.5 | 71.7 | 57.6 | 20.7 | 19.8 |
| VX-3 | – | – | – | – | 48.5 | 44.9 | 47.1 | 77.8 | 51.5 | 48.7 | 21.0 | 18.7 |
| VX-4 | – | – | – | – | – | 44.4 | 49.7 | 47.9 | 53.7 | 78.4 | 20.3 | 19.6 |
| FP-2 | – | – | – | – | – | – | 54.3 | 44.3 | 45.2 | 44.4 | 22.8 | 20.7 |
| FP-3 | – | – | – | – | – | – | – | 47.3 | 50.3 | 49.5 | 20.9 | 21.2 |
| KP-2 | – | – | – | – | – | – | – | – | 57.4 | 49.9 | 21.0 | 20.0 |
| KP-3 | – | – | – | – | – | – | – | – | – | 66.4 | 21.0 | 20.1 |
| KP-4 | – | – | – | – | – | – | – | – | – | – | 20.4 | 19.9 |
| Papain | – | – | – | – | – | – | – | – | – | – | – | 21.7 |
| Substrate | Kcat/Km (s−1 M−1) | |
|---|---|---|
| MP-2 | MP-4 | |
| Z-LR-AMC | 3.9 × 104 | 7.4 × 104 |
| Z-FR-AMC | 3.2 × 104 | 3.8 × 104 |
| Z-RR-AMC | 9.2 × 104 | NH * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lê, H.G.; Kang, J.-M.; Võ, T.C.; Yoo, W.G.; Lee, K.H.; Na, B.-K. Biochemical Properties of Two Plasmodium malariae Cysteine Proteases, Malapain-2 and Malapain-4. Microorganisms 2022, 10, 193. https://doi.org/10.3390/microorganisms10010193
Lê HG, Kang J-M, Võ TC, Yoo WG, Lee KH, Na B-K. Biochemical Properties of Two Plasmodium malariae Cysteine Proteases, Malapain-2 and Malapain-4. Microorganisms. 2022; 10(1):193. https://doi.org/10.3390/microorganisms10010193
Chicago/Turabian StyleLê, Hương Giang, Jung-Mi Kang, Tuấn Cường Võ, Won Gi Yoo, Kon Ho Lee, and Byoung-Kuk Na. 2022. "Biochemical Properties of Two Plasmodium malariae Cysteine Proteases, Malapain-2 and Malapain-4" Microorganisms 10, no. 1: 193. https://doi.org/10.3390/microorganisms10010193
APA StyleLê, H. G., Kang, J.-M., Võ, T. C., Yoo, W. G., Lee, K. H., & Na, B.-K. (2022). Biochemical Properties of Two Plasmodium malariae Cysteine Proteases, Malapain-2 and Malapain-4. Microorganisms, 10(1), 193. https://doi.org/10.3390/microorganisms10010193