
Filtration of the Microalga Amphidinium carterae by the Polychaetes Sabella spallanzanii and Branchiomma luctuosum: A New Tool for the Control of Harmful Algal Blooms?




Abstract
1. Introduction
2. Material and Methods
2.1. Polychaete Sampling
2.2. Microalgal Culture Conditions
2.3. Amphidinium carterae Retention and Clearance Rate Calculation
2.4. Statistical Analysis
3. Results
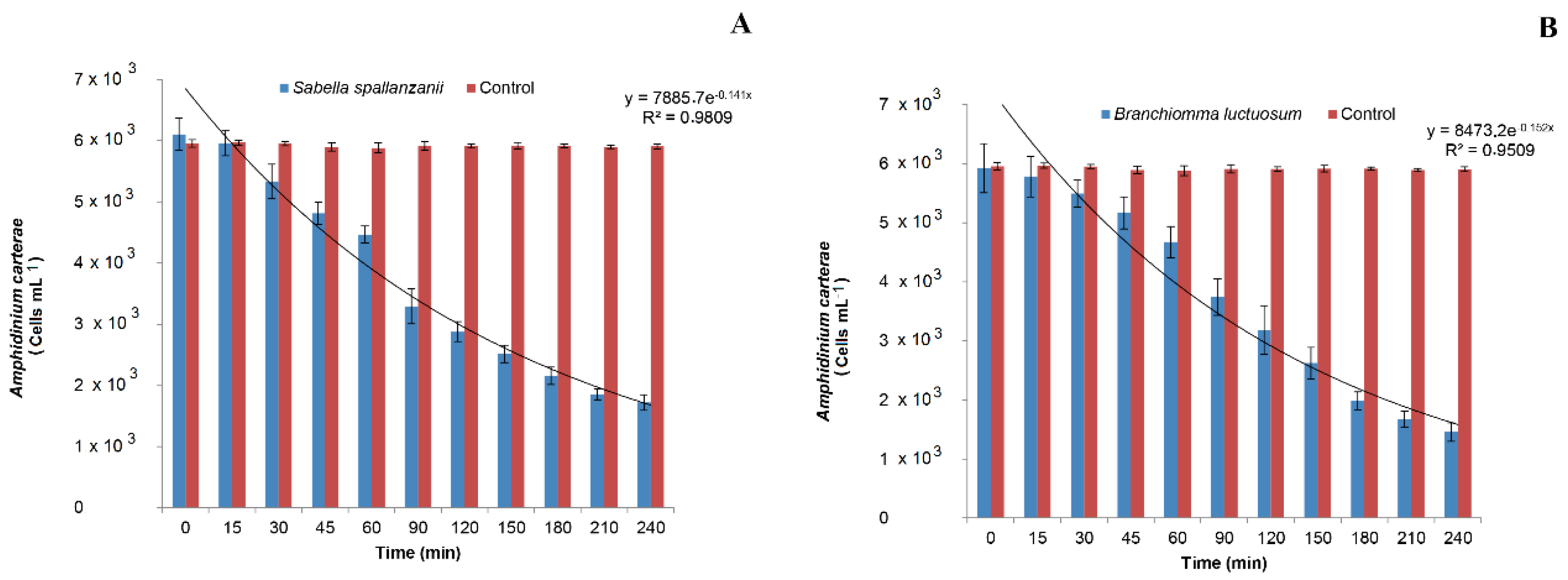
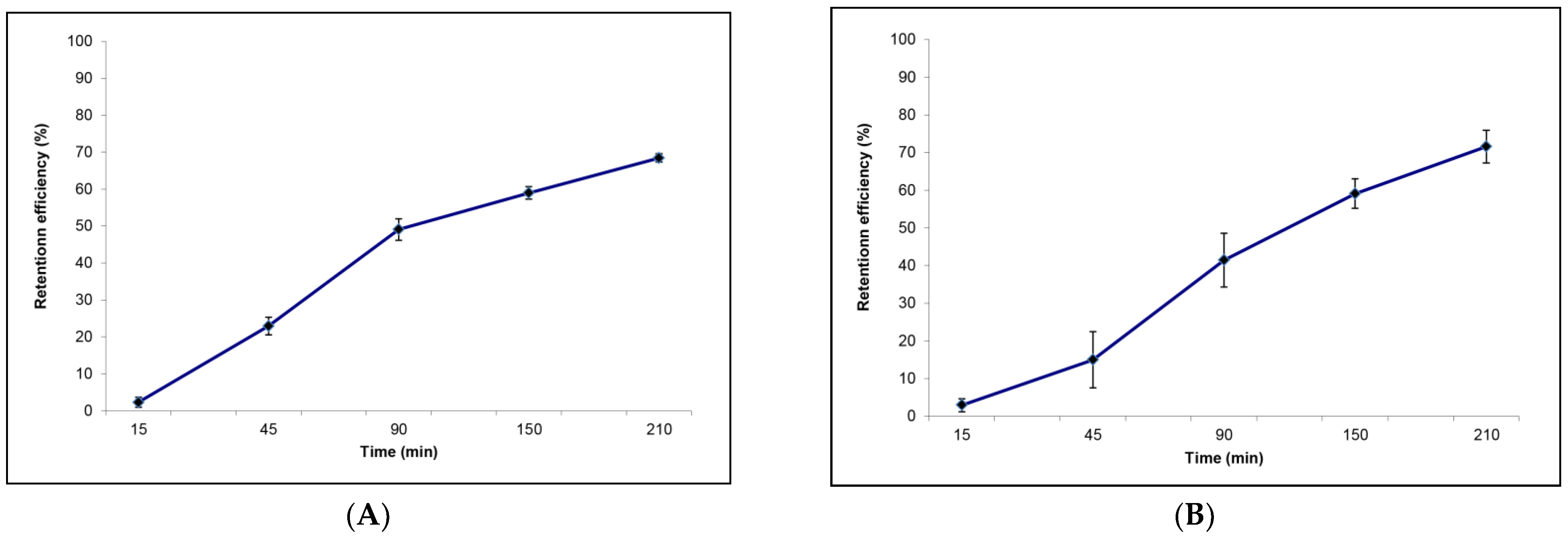
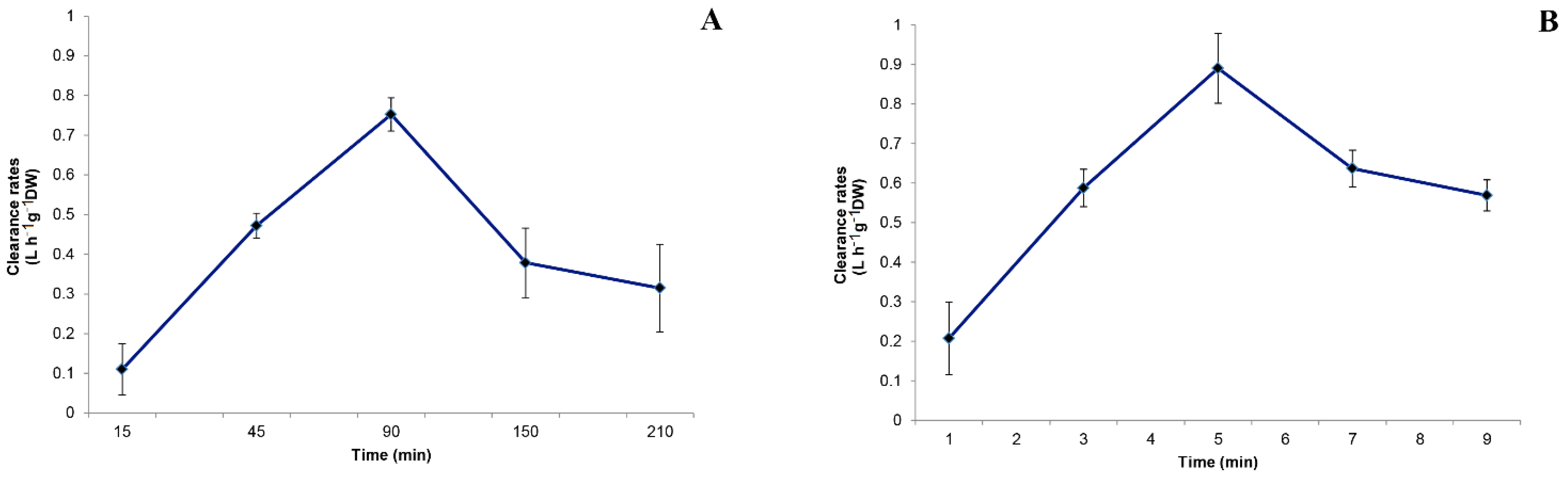
3.1. Filtration Activity on Amphidinium carterae by Sabella spallanzanii
3.2. Filtration activity on Amphidinium carterae by Branchiomma luctuosum
4. Discussion
- -
- The potentially harmful microalgal species A. carterae, here used, when present in the surrounding environment, can be an integral part of the natural diet of the two investigated polychaetes. The Cmax and highest retention efficiencies obtained are 0.936 ± 0.151 L h−1 g−1 DW and 68% for S. spallanzanii, and 1.15 ± 0.204 L h−1 g−1 DW and 72% for B. luctuosum, respectively.
- -
- The here-obtained results on the filtration capability of the polychaetes S. spallanzanii and B. luctuosum on the harmful dinoflagellate A. carterae suggest that these invertebrates represent a new tool to reduce the impact of harmful algae on marine life and could constitute a potential advantage in comparison to the up-to-now used bioremediation methodologies in the bioremediation process. Current strategies to reduce the potential economic and human health effects from HABs to fisheries and aquaculture include chemical (e.g., flocculation [43]), physical, and biological measures [4]. Among these strategies, flocculation has been classified as the most cost-effective and convenient way to rapidly remove algae [58,59,60]. Flocculation by the sole use of natural clay, however, requires a high dosage in order to attain a fairly high (> 90%) removal efficiency [61,62]. New methods were tested to further increase the removal efficiency of natural clay minerals for the mitigation, for example, of A. carterae [41,43], but in general, the use of clay remains often too rudimental, confined, or problematic for large-scale implementation and could potentially have detrimental effects on filter-feeding invertebrates [63,64]. For these reasons, in many countries, severe environmental controls preclude the use of these techniques [3]. As regards physical methods, often used to minimize HABs’ impacts on aquaculture plants, they consist, for example, of enhanced flushing and aeration of cages and/or feeding cessation that would help to manage the problem, but only in a small volume of water [10]. Finally, biological methods are environmentally friendly and can be specific for algal cells, without impact on non-HAB species. These include the use of bacteria, fungi, phages, macroalgae or seagrass, protozoa, zooplankton, and filter-feeding shellfish [4].
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Fernandez-Tejedor, M.; Ismael, A.; Montresor, M.; Mozetič, P.; Taş, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2020, 102, 101843. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2019, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Song, X.; Cao, X.; Liu, Y. Mitigation and Control of Harmful Algal Blooms. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 403–424. [Google Scholar]
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Dechraoui Bottein, M.-Y.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H.; et al. Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts. Commun Earth Env. 2021, 2, 1–10. [Google Scholar] [CrossRef]
- Anderson, D.M. HABs in a changing world: A perspective on harmful algal blooms, their impacts, and research and management in a dynamic era of climactic and environmental change. In Proceedings of the Harmful Algae 2012, 15th International Conference on Harmful Algae, Changwon Gyeongnam, Korea, 29 October–2 November 2012. [Google Scholar]
- Koike, K. A red tide off the Myanmar coast: Morphological and genetic identification of the dinoflagellate composition. Harmful Algae 2013, 27, 149–158. [Google Scholar]
- Kudela, R.M.; Berdalet, E.; Bernard, S.; Burford, M.; Fernand, L.; Lu, S.; Roy, S.; Tester, P.; Usup, G.; Magnien, R.; et al. Harmful Algal Blooms—A Scientific Summary for Policy Makers; IOC/INF-1320; UNESCO: Paris, France, 2015; p. 20. [Google Scholar]
- Pörtner, H.O.; Roberts, D.C.; Masson-Delmotte, V.; Zhai, P.; Tignor, M.; Poloczanska, E.; Mintenbeck, K.; Alegría, A.; Nicolai, M.; Okem, A.; et al. IPCC Special Report on the Ocean and Cryosphere in A Changing Climate; IPCC Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019; Volume 1. [Google Scholar]
- Kim, H.G. Mitigation and controls of HABs. In Ecology of Harmful Algae; Granéli, E., Turner, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 327–338. [Google Scholar]
- Ostroumov, S. Some aspects of water filtering activity of filter feeders. Hydrobiologia 2005, 542, 275–286. [Google Scholar] [CrossRef]
- Ostroumov, S.A. Water Quality and Conditioning in Natural Ecosystems: Biomachinery Theory of Self-Purification of Water. Russ. J. Gen. Chem. 2017, 87, 3199–3204. [Google Scholar] [CrossRef]
- Ostroumov, S.; Widdows, J. Inhibition of mussel suspension feeding by surfactants of three classes. Hydrobiologia 2006, 556, 381–386. [Google Scholar] [CrossRef]
- Licciano, M.; Stabili, L.; Giangrande, A. Clearance rates of Sabella spallanzanii and Branchiomma luctuosum (Annelida: Polychaeta) on a pure culture of Vibrio alginolyticus. Water Res. 2005, 39, 4375–4384. [Google Scholar] [CrossRef]
- Licciano, M.; Stabili, L.; Giangrande, A.; Cavallo, R.A. Bacterial accumulation by Branchiomma luctuosum (Annelida: Polychaeta): A tool for biomonitoring marine systems and restoring polluted waters. Mar. Environ. Res. 2007, 63, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Licciano, M.; Terlizzi, A.; Giangrande, A.; Cavallo, R.A.; Stabili, L. Filter-feeder macroinvertebrates as key players in bacterioplankton biodiversity control: A case of study with Sabella spallanzanii (Polychaeta: Sabellidae). Mar. Environ. Res. 2007, 64, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Widdows, J. Feeding physiology of Cerastoderma edule in response to a wide range of seston concentrations. Mar. Ecol. Prog. Ser. 1997, 152, 175–186. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Larsen, P.S. Filter-feeding in marine macro-invertebrates: Pump characteristics, modelling and energy cost. Biol. Rev. Camb. Philos. Soc. 1995, 70, 67–106. [Google Scholar] [CrossRef] [PubMed]
- Riisgård, H.U.; Larsen, P.S. Particle capture mechanisms in suspension-feeding invertebrates. Mar. Ecol. Prog. Ser. 2010, 418, 255–293. [Google Scholar] [CrossRef]
- Stabili, L.; Licciano, M.; Giangrande, A.; Fanelli, G.; Cavallo, R.A. Sabella spallanzanii filter-feeding on bacterial community: Ecological implications and applications. Mar. Environ. Res. 2006, 61, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Stabili, L.; Licciano, M.; Lezzi, M.; Giangrande, A. Microbiological accumulation by the Mediterranean invasive alien species Branchiomma bairdi (Annelida, Sabellidae): Potential tool for bioremediation. Mar. Pollut. Bull. 2014, 86, 325–331. [Google Scholar] [CrossRef]
- Riisgård, H.U. Properties and energy cost of the muscular piston pump in the suspension feeding polychaete Chaetopterus variopedatus. Mar. Ecol. Prog. Ser. 1989, 56, 157–168. [Google Scholar] [CrossRef]
- Riisgård, H.U. Suspension feeding in the polychaete Nereis diversicolor. Mar. Ecol. Prog. Ser. 1991, 70, 29–37. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Vedel, A.; Boye, H.; Larsen, P.S. Filternet structure and pumping activity in the polychaete Nereis diversicolor: Effects of temperature and pump modelling. Mar. Ecol. Prog. Ser. 1992, 83, 79–89. [Google Scholar] [CrossRef]
- Shulman, G.E.; Finenko, G.A. Bioenergetics of Hydrobionts; Naukova Dumka Press: Kiev, Ukraine, 1990. [Google Scholar]
- Nicol, E.A.T. The Feeding Mechanism, Formation of the Tube, and Physiology of Digestion in Sabella pavonina. Trans. Roy. Soc. Edinb. 1930, 56, 537–598. [Google Scholar] [CrossRef]
- Mayer, S. Particle capture in the crown of the ciliary suspension feeding polychaete Sabella penicillus: Videotape recordings and interpretations. Mar. Biol. 1994, 119, 571–582. [Google Scholar] [CrossRef]
- Bonar, D.B. Feeding and tube construction in Chone mollis Bush (Polychaeta, Sabellidae). J. Exp. Mar. Biol. Ecol. 1972, 9, 1–18. [Google Scholar] [CrossRef]
- Clapin, G. The filtration rate, oxygen consumption and biomass of the introduced polychaete Sabella spallanzanii Gmelin within Cockburn Sound: Can They Control Phytoplankton Level and is it An Efficient Filter Feeder? Honours Thesis, Edith Cowan University, Joondalup, Western Australia, Australia, 1996; p. 90. [Google Scholar]
- Dales, R.P. Preliminary observations on the role of the coelomic cells in food storage and transport in certain polychaetes. J. Mar. Biol. Ass. UK 1957, 36, 91–110. [Google Scholar] [CrossRef]
- Fitzsimons, G. Feeding and tube-building in Sabellastarte magnifica (Shaw) (Sabellidae: Polychaeta). Bull. Mar. Sci. 1965, 15, 642–671. [Google Scholar]
- Lewis, D.B. Feeding and tube-building in the Fabriciinae, (Annelida, Polychaeta). Proc. Linnean Soc. London 1968, 179, 37–49. [Google Scholar] [CrossRef]
- Shumway, S.E.; Bogdanowicz, C.; Dean, D. Oxygen consumption and feeding rates of the sabellid polychaete, Myxicola infundibulum (Renier). Comp. Biochem. Physiol. 1988, 90, 425–428. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Ivarsson, N.M. The crown-filament pump of the suspension feeding polychaete Sabella penicillus: Filtration, effects of temperature, and energy cost. Mar. Ecol. Prog. Ser. 1990, 62, 249–257. [Google Scholar] [CrossRef]
- Wells, G.P. The respiratory significance of the crown in the polychaete worms Sabella and Myxicola. Proc. Roy. Soc. B 1952, 140, 70–82. [Google Scholar]
- Gárate-Lizárraga, I. Proliferation of Amphidinium carterae Gymnodiniales, Gymnodiniacea in Bahía de La Paz, Gulf of California. CICIMAR Oceán 2012, 272, 37–49. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I.; González-Armas, R.; Verdugo-Díaz, G.; Okolodkov, Y.B.; Pérez-Cruz, B.; Díaz-Ortíz, J.A. Seasonality of the dinoflagellate Amphidinium cf. carterae (Dinophyceae: Amphidiniales) in Bahía de la Paz, Gulf of California. Mar. Pollut. Bull. 2019, 146, 532–541. [Google Scholar] [CrossRef]
- Murray, S.A.; Kohli, G.S.; Farrell, H.; Spiers, Z.B.; Place, A.R.; Dorantes-Aranda, J.J.; Ruszczyk, J. A fish kill associated with a bloom of Amphidinium carterae in a coastal lagoon in Sydney, Australia. Harmful Algae 2015, 49, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Aquino-Cruz, A.; Okolodkov, Y.B. Impact of increasing water temperature on growth, photosynthetic efficiency, nutrient consumption, and potential toxicity of Amphidinium cf. carterae and Coolia monotis (Dinoflagellata). Rev. Biol. Mar. Oceanogr. 2016, 51, 565–580. [Google Scholar] [CrossRef]
- Yan, M.; Leung, P.T.Y.; Gu, J.; Veronica, T.T.; Lam, V.T.T.; Murray, J.S.; Harwood, D.T.; Wai, T.C.; Paul, K.S.; Lam, P.K.S. Hemolysis associated toxicities of benthic dinoflagellates from Hong Kong Waters. Mar. Pollut. Bull. 2020, 155, 111114. [Google Scholar] [CrossRef]
- Chen, J.; Pan, G. Harmful algal blooms mitigation using clay/soil/sand modified with xanthan and calcium hydroxide. J. Appl. Phycol. 2012, 24, 1183–1189. [Google Scholar] [CrossRef]
- Liu, S.; Yu, Z.; Song, X.; Cao, X. Effects of modified clay on the physiological and photosynthetic activities of Amphidinium carterae Hulburt. Harmful Algae 2017, 70, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Chen, J.; Donald, M.; Anderson, D.M. Modified local sands for the mitigation of harmful algal blooms. Harmful Algae 2011, 10, 381–387. [Google Scholar] [CrossRef]
- Caroppo, C.; Bisci, A.P. First data on the benthic assemblages of harmful microalgal species in the Gulf of Taranto (Northern Ionian Sea). Rapp. Commun. Int. Mer Médit. 2010, 39, 341. [Google Scholar]
- Pagliara, P.; Caroppo, C. Toxicity assessment of Amphidinium carterae, Coolia cfr. monotis and Ostreopsis cfr. ovata (Dinophyta) isolated from the northern Ionian Sea (Mediterranean Sea). Toxicon 2012, 60, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrates. Select Readings; Berg, C.J., Ed.; Hutchinson Ross Publishing Co: Stroudsberg, PA, USA, 1983; pp. 108–132. [Google Scholar]
- Edler, L.; Elbrächter, M. The Utermöhl method for quantitative phytoplankton analysis. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; UNESCO IOC Manuals and Guides n. 55; UNESCO: Paris, France, 2010; pp. 13–20. [Google Scholar]
- Coughlan, J. The estimation of filtering rate from the clearance of suspensions. Mar. Biol. 1969, 2, 356–358. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; pp. 1–504. [Google Scholar]
- Shumway, S.E.; Egmond, H.P.V.; Hurst, J.W.; Bean, L.L. Management of shellfish resources. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 1995; pp. 433–474. [Google Scholar]
- Berdalet, E.; Fleming, L.E.; Gowen, R.; Davidson, K.; Hess, P.; Backer, L.C.; Moore, S.K.; Hoagland, P.; Enevoldsen, H. Marine harmful algal blooms, human health and wellbeing: Challenges and opportunities in the 21st century. J. Mar. Biol. Assoc. UK 2016, 96, 61–91. [Google Scholar] [CrossRef]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Eutrophication of U.S. Freshwaters: Analysis of Potential Economic Damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar] [CrossRef]
- Buhr, K.J. Suspension-feeding and assimilation efficiency in Lanice conchilega (Polychaeta). Mar. Biol. 1976, 38, 373–383. [Google Scholar] [CrossRef]
- Dales, R.P. Observations on the respiration of the sabellid polychaete Schizobranchia insignis. Biol. Bull. 1961, 121, 82–91. [Google Scholar] [CrossRef]
- Klockner, K. Zur Ökologie von Pomatoceros triqueter (Serpulidae, Polychaeta). Helgol. Wiss. Meeresunters 1978, 31, 219–223. [Google Scholar] [CrossRef]
- Lemmens, J.W.T.J.; Clapin, G.; Lavery, P.; Cary, J. Filtering capacity of seagrass meadows and other habitats of Cockburn Sound, Western Australia. Mar. Ecol. Prog. Ser. 1996, 143, 187–200. [Google Scholar] [CrossRef][Green Version]
- Cugier, P.; Struski, C.; Blanchard, M.; Mazurié, J.; Pouvreau, S.; Olivier, F.; Trigui, J.R.; Thiébaut, E. Assessing the role of benthic filter feeders on phytoplankton production in a shellfish farming site: Mont Saint Michel Bay, France. J. Mar. Syst. 2010, 82, 21–34. [Google Scholar] [CrossRef]
- Kim, H.G. An overview on the occurrences of harmful algal blooms (HABs) and mitigation strategies in Korean coastal waters. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.J., Eds.; Terrapub and Nagasaki University Publication: Tokyo, Japan, 2010; pp. 121–131. [Google Scholar]
- Pan, G.; Miao, X.; Bi, L.; Zhang, H.; Wang, L.; Wang, L.; Wang, Z.; Chen, J.; Ali, J.; Pan, M.; et al. Modified local soil (MLS) technology for harmful algal bloom control, sediment remediation, and ecological restoration. Water 2019, 11, 1123. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Higham, C.J.; Blum, P.; Sengco, M.R.; Anderson, D.M. Removal of harmful algal cells (Karenia brevis) and toxins from seawater culture by clay flocculation. Harmful Algae 2004, 3, 141–148. [Google Scholar] [CrossRef]
- Pan, G.; Zhang, M.M.; Chen, H.; Zou, H.; Yan, H. Removal of cyanobacterial blooms in Taihu Lake using local soils. I. Equilibrium and kinetic screening on the flocculation of Microcystis aeruginosa using commercially available clays and minerals. Environ. Pollut. 2006, 141, 195e200. [Google Scholar] [CrossRef]
- Sengco, M.R.; Li, A.; Tugend, K.; Kulis, D.; Anderson, D.M. Removal of red- and brown-tide cells using clay flocculation. I. Laboratory culture experiments with Gymnodinium breve and Aureococcus anophagefferens. Mar. Ecol. Prog. Ser. 2001, 210, 41–53. [Google Scholar] [CrossRef]
- Anderson, D.M. Approaches to monitoring, control and management of harmful algal blooms (HABs). Ocean Coast Manag. 2009, 52, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Shumway, S.E.; Frank, D.M.; Ewart, L.M. Effect of yellow clay on clearance rate in seven species of benthic, filter-feeding invertebrates. Aquac. Res. 2003, 34, 1391–1402. [Google Scholar] [CrossRef]
- Maguire, J.; Cusack, C.; Ruiz-Villarreal, M.; Silkeb, J.; McElligotta, D.; Davidson, K. Applied simulations and integrated modelling for the understanding of toxic and harmful algal blooms (ASIMUTH): Integrated HAB forecast systems for Europe’s Atlantic Arc 2016. Harmful Algae 2016, 53, 160–166. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Bresnan, E.; Enevoldsen, H.; Schweibold, L.; Zingone, A. Call to contribute to Global HAB Status Report. Harmful Algae News 2017, 58, 1–3. [Google Scholar]
- Zingone, A.; Enevoldsen, H.; Hallegraeff, G.M. Are HABs and their societal impacts expanding and intensifying? A call for answers from the HAB scientific community. In Proceedings of the Marine and Fresh-Water Harmful Algae, 17th International Conference on Harmful Algae 2016; Proenca, L.A.O., Hallegraeff, G.M., Eds.; International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2017; pp. 14–17. [Google Scholar]
- Stabili, L.; Cecere, E.; Licciano, M.; Petrocelli, A.; Sicuro, B.; Giangrande, A. Integrated Multitrophic Aquaculture By-Products with Added Value: The Polychaete Sabella spallanzanii and the Seaweed Chaetomorpha linum as Potential Dietary Ingredients. Mar. Drugs 2019, 17, 677. [Google Scholar] [CrossRef] [PubMed]
- Giangrande, A.; Pierri, C.; Del Pasqua, M.; Gravili, C.; Gambi, M.C.; Gravina, M.F. Mediterranean Sea in check: The other side of biological invasions. Mar. Ecol. 2020, 41, e12583. [Google Scholar] [CrossRef]
- Pal, M.; Yesankar, P.J.; Dwivedi, A.; Asifa Qureshi, A. Biotic control of harmful algal blooms (HABs): A brief review. J. Environ. Manag. 2020, 268, 110687. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | MS | F | p |
|---|---|---|---|---|
| Po | 1 | 157,564,720.3712 | 6543.83 | *** |
| Ti | 10 | 8,376,261.4636 | 347.88 | *** |
| Po × Ti | 10 | 8,036,449.4879 | 333.76 | *** |
| Residual | 110 | 24,078.35 | ||
| TOT | 131 | |||
| Cochran Test Transformation | C = 0.1854 (p < 0.05) None | |||
| SNK Test | AT < AC | |||
| Po(Ti) | (AT T0 = AC T0) | |||
| (AT T1 = AC T1) | ||||
| AT T0 > AT T1 > AT T2 > AT T3 > AT T4 > AT T5 > AT T6 > AT T7 > AT T8 > AT T9 > AT T10 | ||||
| Ti(Po) | (AT T10 = AT T11) | |||
| AC T0 = AC T1= AC T2 = AC T3 = AC T4 = AC T5 = AC T6 = AC T7 = AC T8 = AC T9 = AC T10 = AC11 | ||||
| Source of Variation | DF | MS | F | p |
|---|---|---|---|---|
| Po | 1 | 149,460,080.9 | 2990.75 | *** |
| Ti | 10 | 8,872,472.823 | 177.54 | *** |
| Po × Ti | 10 | 8,561,803.017 | 171.32 | *** |
| Residual | 110 | 49,974.1591 | ||
| TOT | 131 | |||
| Cochran Test Transformation | C = 0.1902 (p < 0.05) None | |||
| SNK Test | AT < AC | |||
| Po(Ti) | (AT T0 = AC T0) | |||
| (AT T1 = AC T1) | ||||
| AT T0 > AT T1 > AT T2 > AT T3 > AT T4 > AT T5 > AT T6 > AT T7 > AT T8 > AT T9 > AT T10 | ||||
| Ti(Po) | (AT T10 = AT T11) | |||
| AC T0 = AC T1= AC T2 = AC T3 = AC T4 = AC T5 = AC T6 = AC T7 = AC T8 = AC T9 = AC T10 = AC11 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabili, L.; Licciano, M.; Giangrande, A.; Caroppo, C. Filtration of the Microalga Amphidinium carterae by the Polychaetes Sabella spallanzanii and Branchiomma luctuosum: A New Tool for the Control of Harmful Algal Blooms? Microorganisms 2022, 10, 156. https://doi.org/10.3390/microorganisms10010156
Stabili L, Licciano M, Giangrande A, Caroppo C. Filtration of the Microalga Amphidinium carterae by the Polychaetes Sabella spallanzanii and Branchiomma luctuosum: A New Tool for the Control of Harmful Algal Blooms? Microorganisms. 2022; 10(1):156. https://doi.org/10.3390/microorganisms10010156
Chicago/Turabian StyleStabili, Loredana, Margherita Licciano, Adriana Giangrande, and Carmela Caroppo. 2022. "Filtration of the Microalga Amphidinium carterae by the Polychaetes Sabella spallanzanii and Branchiomma luctuosum: A New Tool for the Control of Harmful Algal Blooms?" Microorganisms 10, no. 1: 156. https://doi.org/10.3390/microorganisms10010156
APA StyleStabili, L., Licciano, M., Giangrande, A., & Caroppo, C. (2022). Filtration of the Microalga Amphidinium carterae by the Polychaetes Sabella spallanzanii and Branchiomma luctuosum: A New Tool for the Control of Harmful Algal Blooms? Microorganisms, 10(1), 156. https://doi.org/10.3390/microorganisms10010156