Natural Feed Additives Modulate Immunity and Mitigate Infection with Sphaerospora molnari (Myxozoa: Cnidaria) in Common Carp: A Pilot Study



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
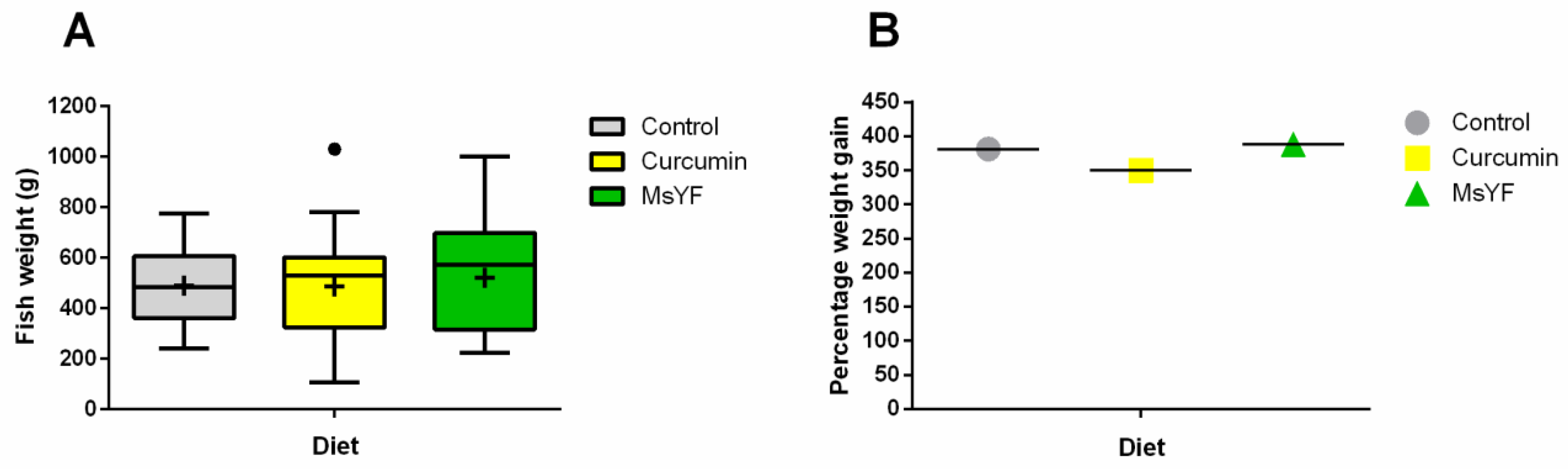
2.1. Diets Induce Similar Weight Gain in All Investigated Groups
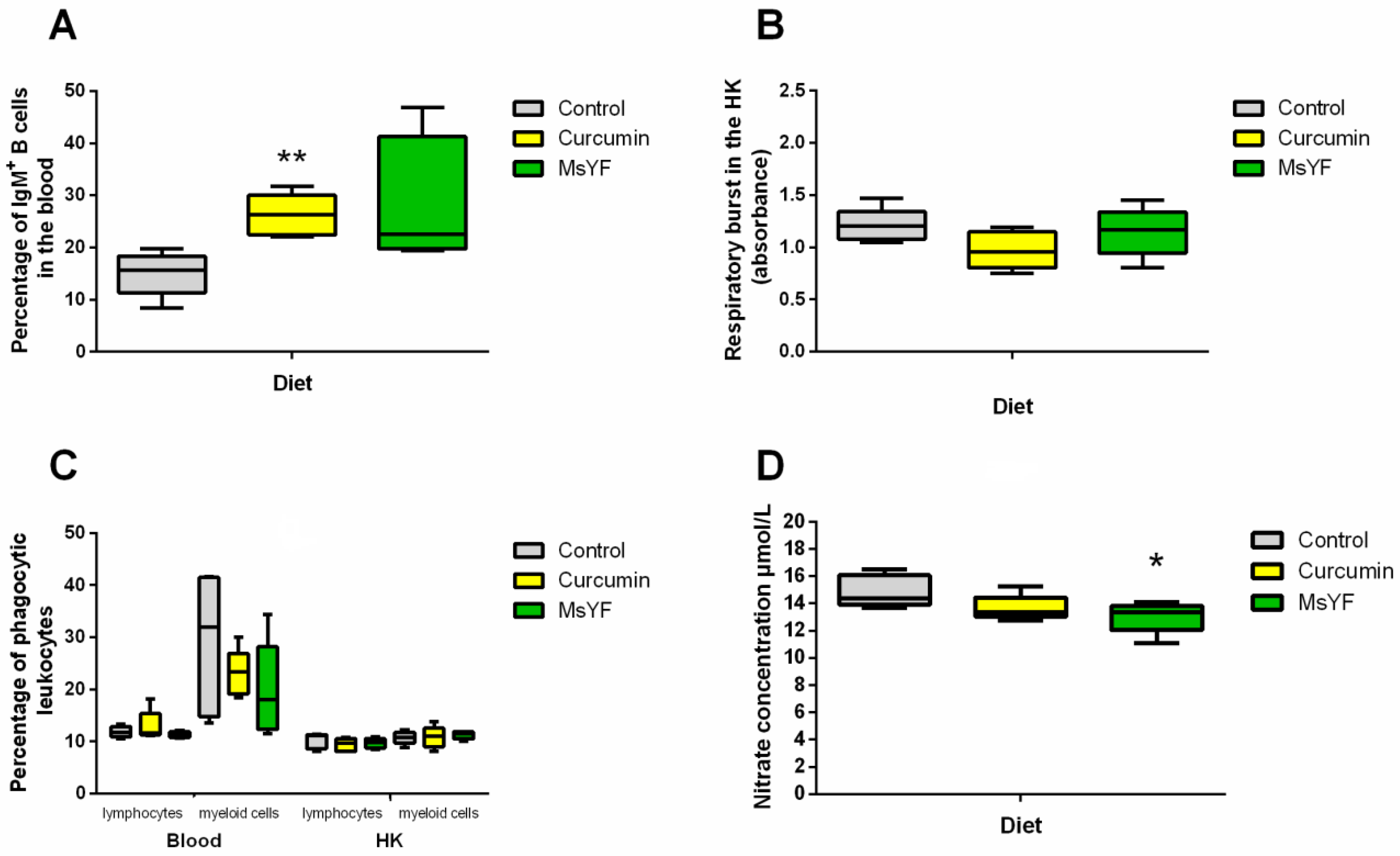
2.2. Functional Diets Induce Changes in the Immune Parameters of Uninfected Carp
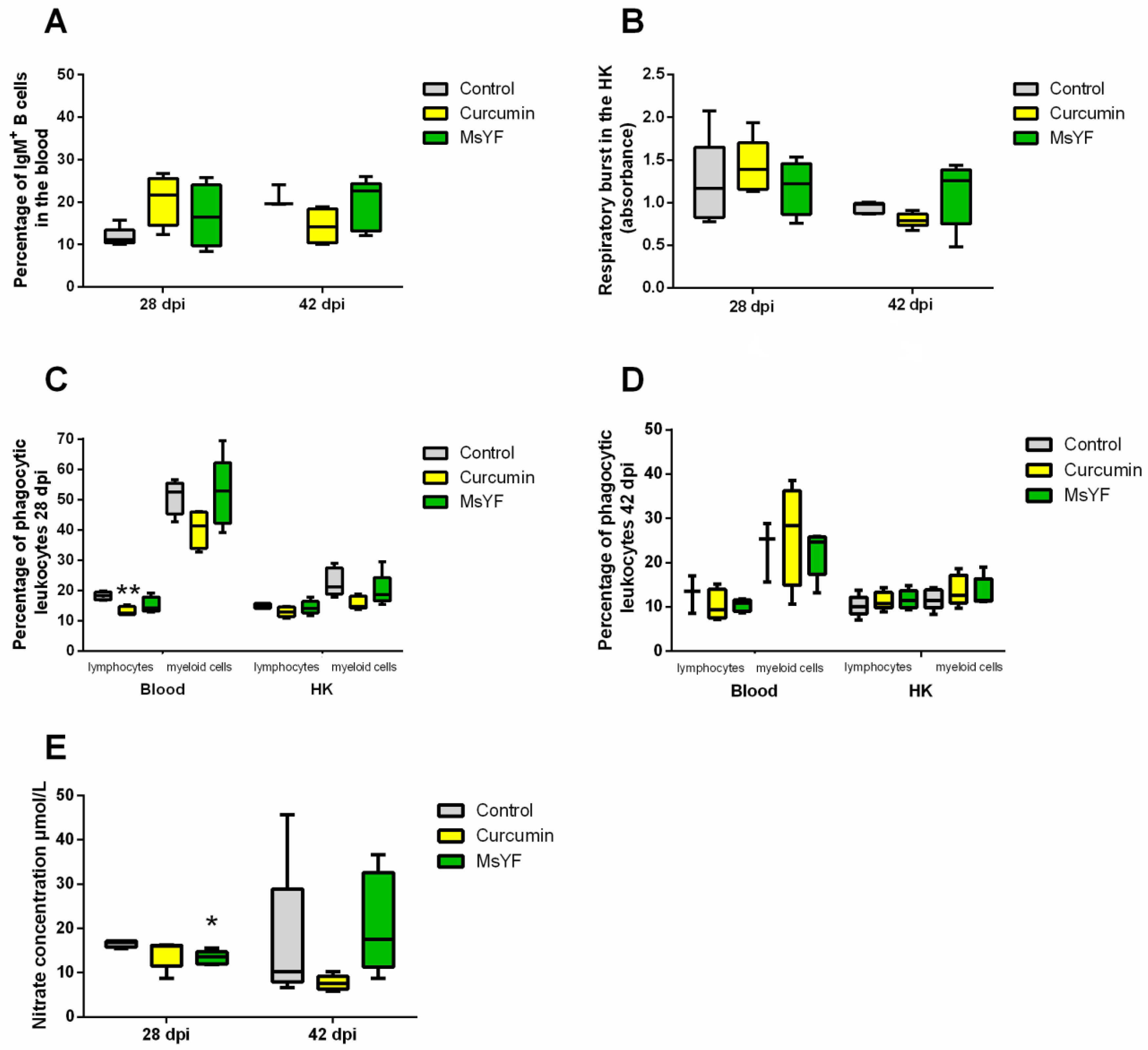
2.3. Myxozoan Infection Causes Differential Responses in Carp Fed Curcumin vs. Yeast-Enriched Diets
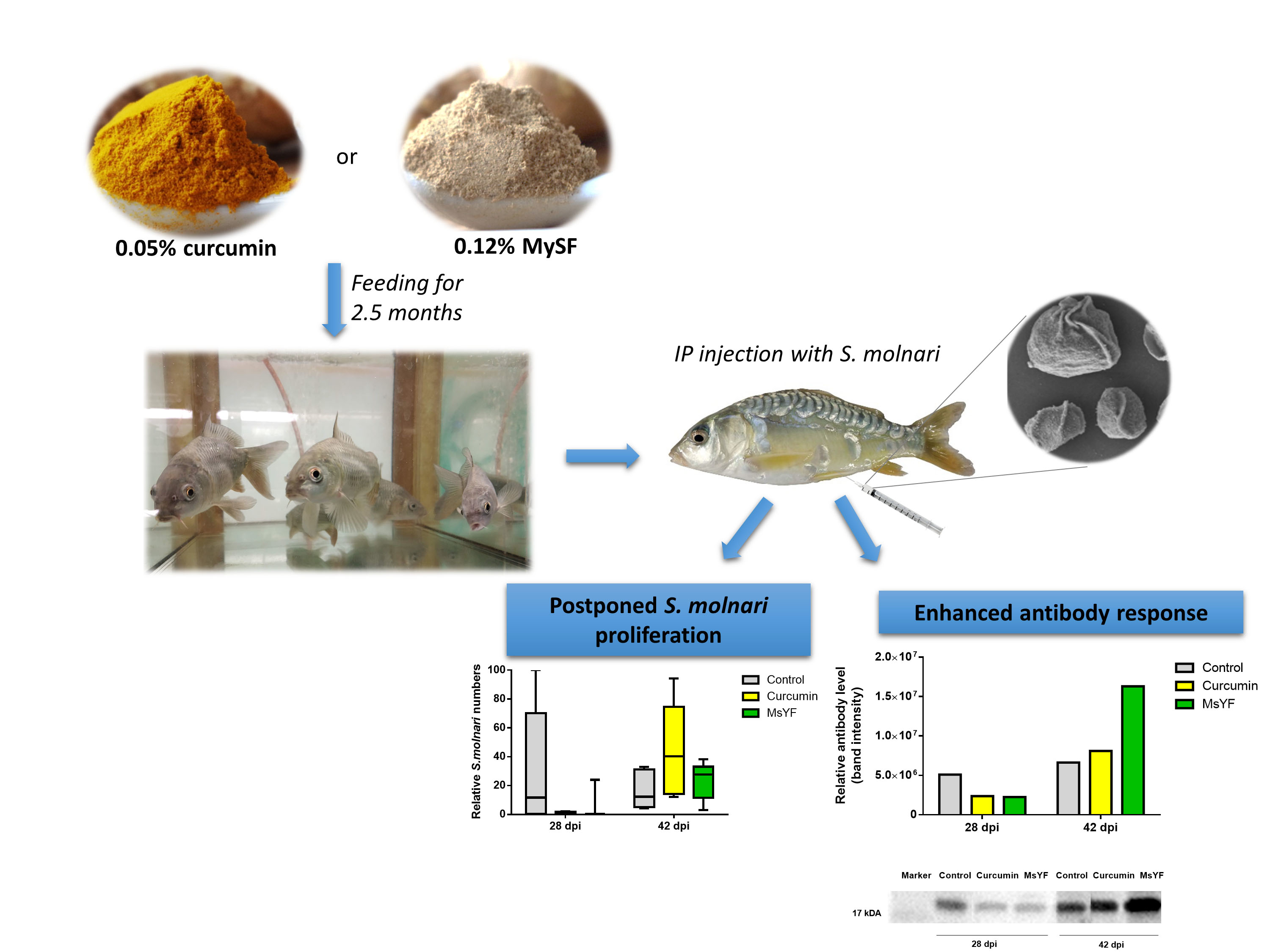
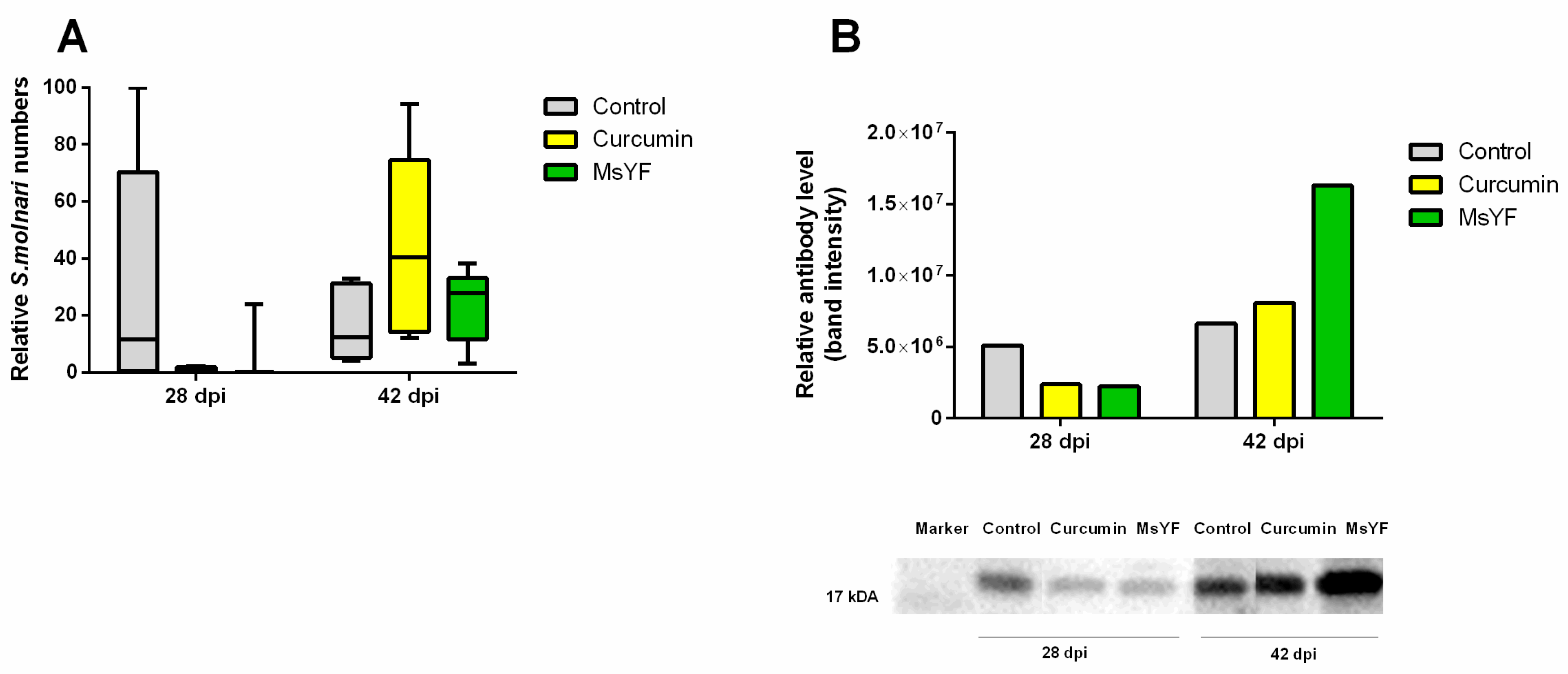
2.4. Functional Diets Decrease S. molnari Numbers and Enhance Specific Immunity
3. Discussion
4. Materials and Methods
4.1. Isolation of S. Molnari Blood Stages
4.2. Experimental Design
4.3. Sampling
4.4. Isolation of Leukocytes from Blood and HK
4.5. Respiratory Burst
4.6. Phagocytosis
4.7. Nitric Oxide (NO) Quantification
4.8. B cell Numbers
4.9. S. molnari Quantification
4.10. Detection of S. molnari-Specific Antibodies
4.11. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedrick, R.P.; Adkison, M.A.; El-Matbouli, M.; MacConnell, E. Whirling disease: Re-emergence among wild trout. Immunol. Rev. 1998, 166, 365–376. [Google Scholar] [CrossRef]
- Hedrick, R.P.; MacConnell, E.; de Kinkelin, P. Proliferative kidney disease of salmonid fish. Ann. Rev. Fish Dis. 1993, 3, 277–290. [Google Scholar] [CrossRef]
- Bowser, P.R.; Conroy, J.D. Histopathology of gill lesions in Channel catfish associated with Henneguya. J. Wildl. Dis. 1985, 21, 177–179. [Google Scholar] [CrossRef]
- Ronza, P.; Robledo, D.; Bermúdez, R.; Losada, A.; Pardo, B.; Martinez, P.; Quiroga, M. Integrating genomic and morphological approaches in fish pathology research: The case of turbot (Scophthalmus maximus) Enteromyxosis. Front. Genet. 2019, 10, 26. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Gil-Solsona, R.; Estensoro, I.; Piazzon, M.C.; Martos-Sitcha, J.A.; Picard-Sánchez, A.; Fuentes, J.; Sancho, J.V.; Calduch-Giner, J.A.; Hernández, F.; et al. Disruption of gut integrity and permeability contributes to enteritis in a fish-parasite model: A story told from serum metabolomics. Parasit. Vect. 2019, 12, 486. [Google Scholar] [CrossRef]
- Gómez, D.; Bartholomew, J.; Sunyer, J.O. Biology and mucosal immunity to myxozoans. Dev. Comp. Immunol. 2014, 43, 243–256. [Google Scholar] [CrossRef]
- Carraro, L.; Bertuzzo, E.; Mari, L.; Fontes, I.; Hartikainen, H.; Strepparava, N.; Schmidt-Posthaus, H.; Wahli, T.; Jokela, J.; Gatto, M.; et al. Integrated field, laboratory, and theoretical study of PKD spread in a Swiss prealpine river. Proc. Natl. Acad. Sci. USA 2017, 114, 201713691. [Google Scholar] [CrossRef]
- Okamura, B.; Hartikainen, H.; Schmidt-Posthaus, H.; Wahli, T. Life cycle complexity, environmental change and the emerging status of salmonid proliferative kidney disease. Freshw. Biol. 2011, 56, 735–753. [Google Scholar] [CrossRef]
- Bruneaux, M.; Visse, M.; Gross, R.; Pukk, L.; Saks, L.; Vasemägi, A. Parasite infection and decreased thermal tolerance: Impact of proliferative kidney disease (PKD) on a wild salmonid fish in the context of climate change. Funct. Ecol. 2017, 31, 216–226. [Google Scholar] [CrossRef]
- Debes, P.; Gross, R.; Vasemägi, A. Quantitative genetic variation in, and environmental effects on, pathogen resistance and temperature-dependent disease severity in a wild trout. Am. Nat. 2017, 190, 244–265. [Google Scholar] [CrossRef]
- Skovgaard, A.; Buchmann, K. Tetracapsuloides bryosalmonae and PKD in juvenile wild salmonids in Denmark. Dis. Aquat. Org. 2012, 101, 33–42. [Google Scholar] [CrossRef][Green Version]
- Hutchins, P.R.; Sepulveda, A.J.; Martin, R.; Hopper, L. A probe-based quantitative PCR assay for detecting Tetracapsuloides bryosalmonae in fish tissue and environmental DNA water samples. Conserv. Genet. Resour. 2018, 10, 317–319. [Google Scholar] [CrossRef]
- Robbins, J. Tiny Invader, Deadly to Fish, Shuts Down a River in Montana. New York Times. 2016. Available online: https://www.nytimes.com/2016/08/24/us/tiny-parasite-invader-deadly-to-fish-shuts-down-yellowstone-river-in-montana.html (accessed on 20 November 2020).
- Holzer, A.S.; Hartigan, A.; Patra, S.; Pecková, H.; Eszterbauer, E. Molecular fingerprinting of the myxozoan community in common carp suffering swim bladder inflammation (SBI) identifies multiple etiological agents. Parasit. Vect. 2014, 7, 398. [Google Scholar] [CrossRef][Green Version]
- Davies, I.; Rodger, G.A. Review of the use of ivermectin as a treatment for sea lice (Lepeophtheirus salmonis (Krøyer) and Caligus elongatus Nordmann) infestation in farmed Atlantic salmon (Salmo salar L.). Aquac. Res. 2001, 31, 869–883. [Google Scholar] [CrossRef]
- Collier, L.M.; Pinn, E.H. An assessment of the acute impact of the sea lice treatment ivermectin on a benthic community. J. Exp. Mar. Biol. Ecol. 1998, 230, 131–147. [Google Scholar] [CrossRef]
- Whipple, M.J.; Gannam, A.L.; Bartholomew, J.L. Inability to control ceratomyxosis in rainbow trout and steelhead with oral treatments of glucan immunostimulant or the fumagillin analog TNP-470. N. Am. J. Aquac. 2002, 64, 1–9. [Google Scholar] [CrossRef]
- Palenzuela, O.; Del Pozo, R.; Piazzon, M.C.; Isern-Subich, M.M.; Ceulemans, S.; Coutteau, P.; Sitja-Bobadilla, A. Effect of a functional feed additive on mitigation of experimentally induced gilthead sea bream Sparus aurata enteromyxosis. Dis. Aquat. Org. 2020, 138, 111–120. [Google Scholar] [CrossRef]
- Korytář, T.; Wiegertjes, G.; Zusková, E.; Tomanová, A.; Lisnerová, M.; Patra, S.; Sieranski, V.; Šíma, R.; born-Torrijos, A.; Wentzel, A.S.; et al. The kinetics of cellular and humoral immune responses of common carp to presporogonic development of the myxozoan Sphaerospora molnari. Parasit. Vect. 2019, 12, 208. [Google Scholar] [CrossRef]
- Haddad, M.; Sauvain, M.; Deharo, E. Curcuma as a parasiticidal agent: A review. Planta Med. 2011, 77, 672–678. [Google Scholar] [CrossRef]
- Cao, L.; Ding, W.; Du, J.; Jia, R.; Liu, Y.; Zhao, C.; Shen, Y.; Yin, G. Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinus carpio var. Jian) with CCl4-induced liver damage. Fish Shellfish Immunol. 2015, 43, 150–157. [Google Scholar] [CrossRef]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2007, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Mallo, N.; DeFelipe, A.P.; Folgueira, I.; Sueiro, R.A.; Lamas, J.; Leiro, J.M. Combined antiparasitic and anti-inflammatory effects of the natural polyphenol curcumin on turbot scuticociliatosis. J. Fish Dis. 2017, 40, 205–217. [Google Scholar] [CrossRef]
- Brunet, L.R. Nitric oxide in parasitic infections. Int. Immunopharmacol. 2001, 1, 1457–1467. [Google Scholar] [CrossRef]
- Petit, J.; Wiegertjes, G.F. Long-lived effects of administering β-glucans: Indications for trained immunity in fish. Dev. Comp. Immunol. 2016, 64, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, M.; Vax, A.; Formosa, C.; Martin-Yken, H.; Dague, E.; François, J.M. A combined chemical and enzymatic method to determine quantitatively the polysaccharide components in the cell wall of yeasts. FEMS Yeast Res. 2014, 14, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Abarca, A.; Bethke, J.; Narvaez, E.; Flores, R.; Mercado, L. Parameters to evaluate the immunostimulant effect of Zymosan A in head kidney leucocytes (HKL) of salmonids. Lat. Am. J. Aquat. Res. 2012, 40, 545–552. [Google Scholar] [CrossRef]
- Bridle, A.R.; Carter, C.G.; Morrison, R.N.; Nowak, B.F. The effect of β-glucan administration on macrophage respiratory burst activity and Atlantic salmon, Salmo salar L., challenged with amoebic gill disease–evidence of inherent resistance. J. Fish Dis. 2005, 28, 347–356. [Google Scholar] [CrossRef]
- Cook, M.T.; Hayball, P.J.; Hutchinson, W.; Nowak, B.; Hayball, J.D. The efficacy of a commercial beta-glucan preparation, EcoActiva, on stimulating respiratory burst activity of head-kidney macrophages from pink snapper (Pagrus auratus), Sparidae. Fish Shellfish Immunol. 2001, 11, 661–672. [Google Scholar] [CrossRef]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish Physiol. Biochem. 2013, 39, 431–457. [Google Scholar] [CrossRef]
- Rawling, M.D.; Pontefract, N.; Rodiles, A.; Anagnostara, I.; Leclercq, E.; Schiavone, M.; Castex, M.; Merrifield, D.L. The effect of feeding a novel multistrain yeast fraction on European seabass (Dicentrachus labrax) intestinal health and growth performance. J. World Aquac. Soc. 2019, 50, 1108–1122. [Google Scholar] [CrossRef]
- Aakre, R.; Wergeland, H.I.; Aasjord, P.M.; Endresen, C. Enhanced antibody response in Atlantic salmon (Salmo salar L.) to Aeromonas salmonicida cell wall antigens using a bacterin containing β-1,3-M-glucan as adjuvant. Fish Shellfish Immunol. 1994, 4, 47–61. [Google Scholar] [CrossRef]
- Chen, D.; Ainsworth, A.J. Glucan administration potentiates immune defence mechanisms of. channel catfish, Ictalurus punctatus Rafinesque. J. Fish Dis. 1992, 15, 295–304. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A. Fish immune response to myxozoan parasites. Parasite 2008, 15, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Ogawa, K.; Wakabayashi, H. Humoral immune response of carp Cyprinus carpio to Myxobolus artus (Myxozoa: Myxobolidae) infection. J. Fish Biol. 1993, 43, 441–450. [Google Scholar] [CrossRef]
- Saulnier, D.; Kinkelin, P. Antigenic and biochemical study of PKX, the myxosporean causative agent of proliferative kidney disease of salmonid fish. Dis. Aquat. Org. 1996, 27, 103–114. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Palenzuela, O.; Riaza, A.; Macías, M.A.; Alvarez-Pellitero, P. Protective acquired immunity to Enteromyxum scophthalmi (Myxozoa) is related to specific antibodies in Psetta maxima (L.) (Teleostei). Scand. J. Immunol. 2007, 66, 26–34. [Google Scholar] [CrossRef]
- Sommerset, I.; Krossøy, B.; Biering, E.; Frost, P. Vaccines for fish in aquaculture. Expert Rev. Vaccines 2005, 4, 89–101. [Google Scholar] [CrossRef]
- Korytář, T.; Chan, J.T.H.; Vancová, M.; Holzer, A.S. Blood feast: Exploring the erythrocyte-feeding behaviour of the myxozoan Sphaerospora molnari. Paras. Immunol. 2020, 42, e12683. [Google Scholar] [CrossRef]
- Li, J.; Barreda, D.R.; Zhang, Y.-A.; Boshra, H.; Gelman, A.E.; LaPatra, S.; Tort, L.; Sunyer, J.O. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Nat. Immunol. 2006, 7, 1116–1124. [Google Scholar] [CrossRef]
- Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. Tissue preservation and total DNA extraction from fish stored at ambient temperature using buffers containing high concentration of urea. Fish. Sci. 1996, 62, 727–730. [Google Scholar] [CrossRef]
- Holzer, A.S.; Sommerville, C.; Wootten, R. Molecular relationships and phylogeny in a community of myxosporeans and actinosporeans based on their 18S rDNA sequences. Int. J. Parasitol. 2004, 34, 1099–1111. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 European Union; Licensee MDPI, Basel, Switzerland. This is an open access article distributed under the terms of the Creative Commons Attribution IGO License (http://creativecommons.org/licenses/by/3.0/igo/legalcode), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In any reproduction of this article there should not be any suggestion that EU or this article endorse any specific organisation or products. The use of the EU logo is not permitted.
Share and Cite
Ganeva, V.O.; Korytář, T.; Pecková, H.; McGurk, C.; Mullins, J.; Yanes-Roca, C.; Gela, D.; Lepič, P.; Policar, T.; Holzer, A.S. Natural Feed Additives Modulate Immunity and Mitigate Infection with Sphaerospora molnari (Myxozoa: Cnidaria) in Common Carp: A Pilot Study. Pathogens 2020, 9, 1013. https://doi.org/10.3390/pathogens9121013
Ganeva VO, Korytář T, Pecková H, McGurk C, Mullins J, Yanes-Roca C, Gela D, Lepič P, Policar T, Holzer AS. Natural Feed Additives Modulate Immunity and Mitigate Infection with Sphaerospora molnari (Myxozoa: Cnidaria) in Common Carp: A Pilot Study. Pathogens. 2020; 9(12):1013. https://doi.org/10.3390/pathogens9121013
Chicago/Turabian StyleGaneva, Vyara O., Tomáš Korytář, Hana Pecková, Charles McGurk, Julia Mullins, Carlos Yanes-Roca, David Gela, Pavel Lepič, Tomáš Policar, and Astrid S. Holzer. 2020. "Natural Feed Additives Modulate Immunity and Mitigate Infection with Sphaerospora molnari (Myxozoa: Cnidaria) in Common Carp: A Pilot Study" Pathogens 9, no. 12: 1013. https://doi.org/10.3390/pathogens9121013
APA StyleGaneva, V. O., Korytář, T., Pecková, H., McGurk, C., Mullins, J., Yanes-Roca, C., Gela, D., Lepič, P., Policar, T., & Holzer, A. S. (2020). Natural Feed Additives Modulate Immunity and Mitigate Infection with Sphaerospora molnari (Myxozoa: Cnidaria) in Common Carp: A Pilot Study. Pathogens, 9(12), 1013. https://doi.org/10.3390/pathogens9121013