1. Introduction
Fusarium ear and stalk rot are ubiquitous diseases of maize with high economic impact in agriculture [
1]. Several
Fusarium species infecting maize are known to produce toxic secondary metabolites, called mycotoxins, which impair grain quality and threaten the safety of animal feed and food products [
2,
3]. Among the most important
Fusarium species inciting pre- and post-harvest ear rot of maize are
F. graminearum and
F. verticillioides [
4,
5,
6], but also other species such as
Fusarium poae [
7,
8],
Fusarium proliferatum [
1],
F. subglutinans [
9], and
F. temperatum [
10] are frequently reported. Ear infection is typically characterized by the growth of white or reddish mycelium with rot induced on the cob and on stored grains.
F. subglutinans, which is a member of the
Fusarium fujikuroi species complex (FFSC), predominantly occurs in temperate climate regions.
F. subglutinans was elevated to the species level in 1983 by Nelson et al. [
11], after separation from
F. verticillioides. Later on, in 2002, Steenkamp et al. [
12] reported two cryptic species within a set of isolates of
F. subglutinans, obtained from several locations, based on phylogenetic concordance analyses of six nuclear regions, and suggested that both subspecies justify separation into two individual taxa. Phylogenetic separation was further supported by mycotoxin analyses revealing the production of beauvericin (BEA), which was exclusively produced by European isolates belonging to the so-called group 1 of
F. subglutinans [
13]. Furthermore, separation of strains was based on differences in climatic requirements, since isolates of
F. subglutinans group 1 were frequently collected in cooler regions like Germany, Poland, and Austria while
F. subglutinans group 2 prevailed in warmer and dryer regions such as Slovakia, Italy, and Serbia [
13]. Several other studies also reported the detection of mycotoxins, such as beauvericin [
14,
15] fusaproliferin (FUSA) [
16], moniliformin (MON) [
17], and rarely fumonisin B1 (FB1) [
18], produced by the subgroup of
F. subglutinans, presumably
F. subglutinans group 1. In 2011, this cryptic subgroup was classified as a novel species, establishing
F. temperatum as
species nova, corresponding to the formerly known group 1 of
F. subglutinans [
10]. The mycotoxin profile of
F. temperatum is not yet fully clarified. While the production of beauvericin was consistently found in all strains of
F. temperatum, moniliformin and fumonisins were produced only by a single isolate of
F.temperatum as reported by Scauflaire et al. [
19]. Similarly, production of beauvericin, moniliformin, fusaproliferin, and fumonisins was reported for
F. temperatum strains from Argentina [
20]. Further studies report on
F. temperatum infection on maize causing seedling blight and root rot [
21] and ear rot, as well as head blight on wheat [
22].
The aim of the study was to determine the occurrence of F. temperatum and F. subglutinans on maize ears and stalks in Germany and to assess their pathogenicity relative to each other and to other common Fusarium ear rot and head blight pathogens. In addition, the mycotoxin profiles of F. temperatum and F. subglutinans were compared.
3. Discussion
In the monitoring of 2017 and 2018,
F. temperatum was found to be the third most often occurring
Fusarium species on maize cobs in Germany. Among all samples, 17% of cob and 17% of stalk samples were infected with
F. temperatum while only 2% samples were infected with
F. subglutinans. Similar findings have recently been reported from several European countries, including Poland [
33], France [
34], and Belgium [
10], as well as China [
22] and South Korea [
35].
Environmental factors like precipitation and humidity are known to strongly affect the occurrence and disease severity of several
Fusarium species [
1,
11,
36,
37]. During the two years of monitoring, weather conditions in July differed considerably. In 2017, high precipitation (110.4 mm) occurred and mean temperatures were around 18.8 °C, while in 2018 precipitation was low (40.4 mm) and temperatures were high (20.6 °C). Nonetheless, the frequency of
F. subglutinans did not change between the years, while the occurrence of
F. temperatum slightly increased in 2018, suggesting warmer temperatures and low precipitation to be favorable for the latter. Similar tendencies were observed in the inoculation experiments in climate chambers. Disease severity on ears inoculated with
F. temperatum was highest at 24 °C, while infection with
F. subglutinans peaked at 21 °C. Even before
F. temperatum and
F. subglutinans were defined as individual species, Moretti et al. [
13] suggested that a separation into two subgroups may be based on different temperature and humidity requirements, which may have resulted from physiological changes in their sites of origin. Similarly, numerous studies related the occurrence of
F. temperatum and
F. subglutinans to temperature and humidity conditions. Moretti et al. [
13] observed that
F. subglutinans occurred more often in warmer and drier regions such as Italy, Slovakia, and Serbia, while
F. temperatum was reported more often from Germany, Poland, Austria, and Switzerland. These findings correspond to several studies from Belgium [
10], China [
22] and Argentina [
20] indicating higher frequencies of
F. temperatum in moderate to cool and moist regions with mean temperatures of 18 °C or lower, while other studies reported
F. temperatum more often in Poland [
33] and Germany [
38] following dry conditions. Marin et al. [
39] demonstrated that the growth rate of most
Fusarium ssp. increased with increasing water activity (a
w value); however, the growth rate of
F. proliferatum and
F. subglutinans decreased at 25 °C when the a
w value increased from 0.980 to 0.995. Further studies are needed to clarify the effect of temperature and precipitation on the occurrence of and disease incidence caused by
F. temperatum and
F. subglutinans.
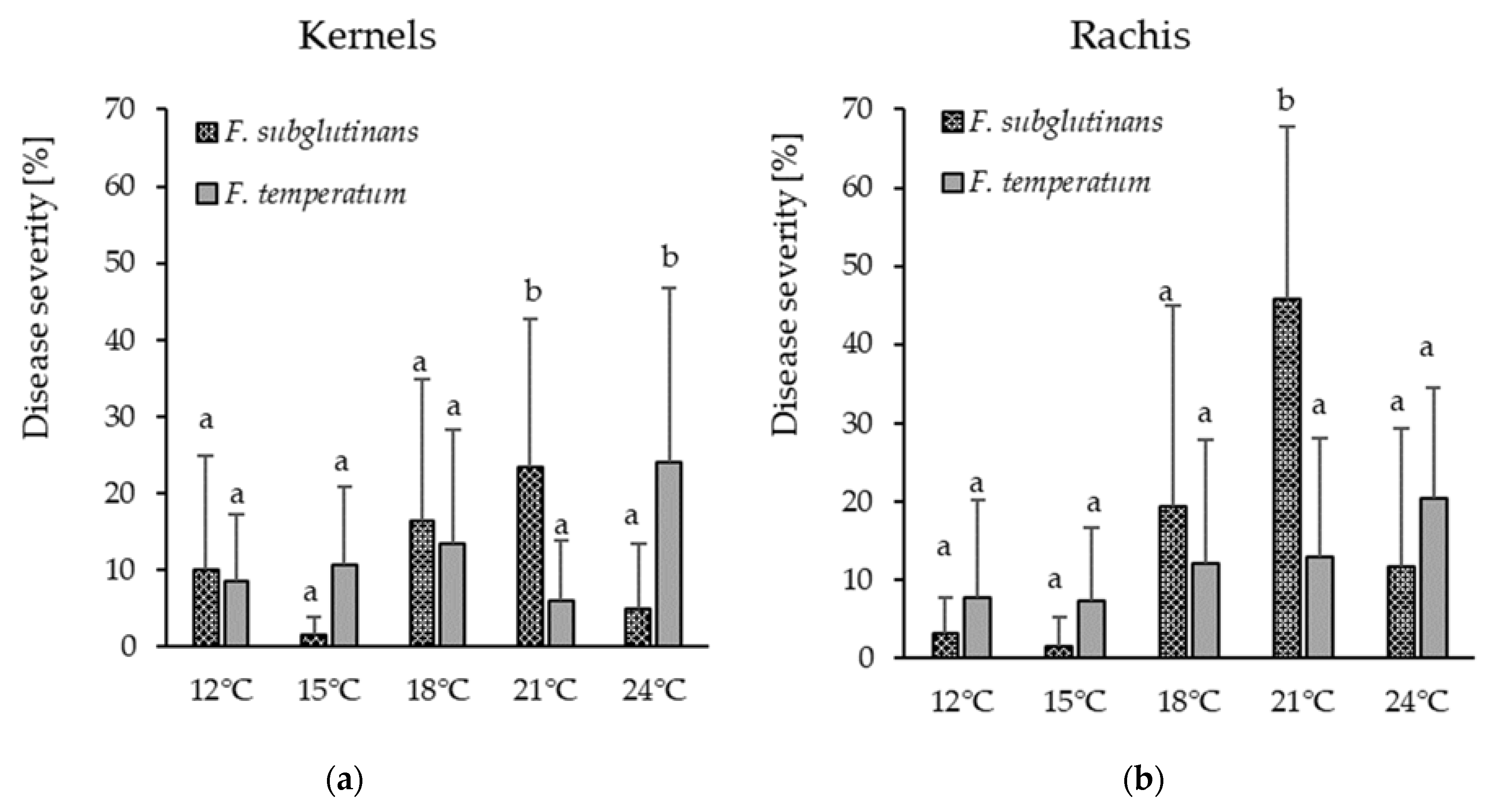
Field inoculation studies at five locations in Germany in 2017 and 2018 showed
F. temperatum to be the most aggressive
Fusarium species in maize, even as compared to
F. graminearum and
F. verticillioides. However, no significant differences in pathogenicity were observed between
F. temperatum and
F. subglutinans at a field site in 2018. This observation confirms the particular importance of
F. temperatum as an ear rot pathogen in maize cultivation in Germany and other locations with similar climate. The low visual infection rate of
F. verticillioides may be explained by symptomless infection and endophytic colonization of maize ears; therefore, disease symptoms may not reflect plant colonization and mycotoxin concentration accurately [
40,
41,
42,
43].
Inoculation of maize stalks with toothpicks showed that
F. temperatum and
F. subglutinans are pathogenic on the stalks. However, compared to common stalk rot pathogens like
F. graminearum,
F. culmorum, and
F. crookwellense, disease severity was relatively low. This corresponds to the results of Levic et al. [
44] and Scauflaire et al. [
19], who reported the formation of necrotic lesions and symptoms like wilting, stunting, and rotting on stalks and leaf sheaths by
F. temperatum and
F. subglutinans yet lower disease severity as compared to
F. crookwellense,
F. verticillioides,
F. culmorum, and
F. graminearum [
21].
Crop residues of maize infected with
Fusarium spp. are considered a major inoculum source for
Fusarium diseases in small grain cereals in Europe, such as seedling and root rot at the seedling stage and
Fusarium head blight during anthesis [
45,
46].
Fusarium head blight of wheat is mainly caused by
F. graminearum,
F. culmorum,
F. poae,
Fusarium tricinctum, and
Fusarium avenaceum [
47,
48,
49]. In addition,
F. subglutinans was reported to rarely infect wheat, causing contamination with MON in small grain cereals from central to north-east European countries [
48,
50].
F. proliferatum occasionally infects wheat, causing contamination with fumonisins and BEA [
51,
52]. In our study, all
F. temperatum and
F. subglutinans isolates were able to infect winter wheat and cause
Fusarium head blight at anthesis. The severity of disease caused by
F. graminearum was highest and the colonization of the plant advanced with the highest rate, while 21 days post infection no significant differences between
F. temperatum and
F. graminearum were found with regard to disease severity and thousand-kernel weight. In line with our results, the investigations of Wang et al. [
22] demonstrated pathogenicity of
F. temperatum and
F. subglutinans on wheat; however, the aggressiveness of tested strains was significantly lower than the aggressiveness of a control strain of
Fusarium asiaticum.Identification of
F. temperatum and
F. subglutinans was carried out by species-specific PCR assays [
23,
53] and strengthened by the analysis of the marker gene
TEF-1α, as previously reported [
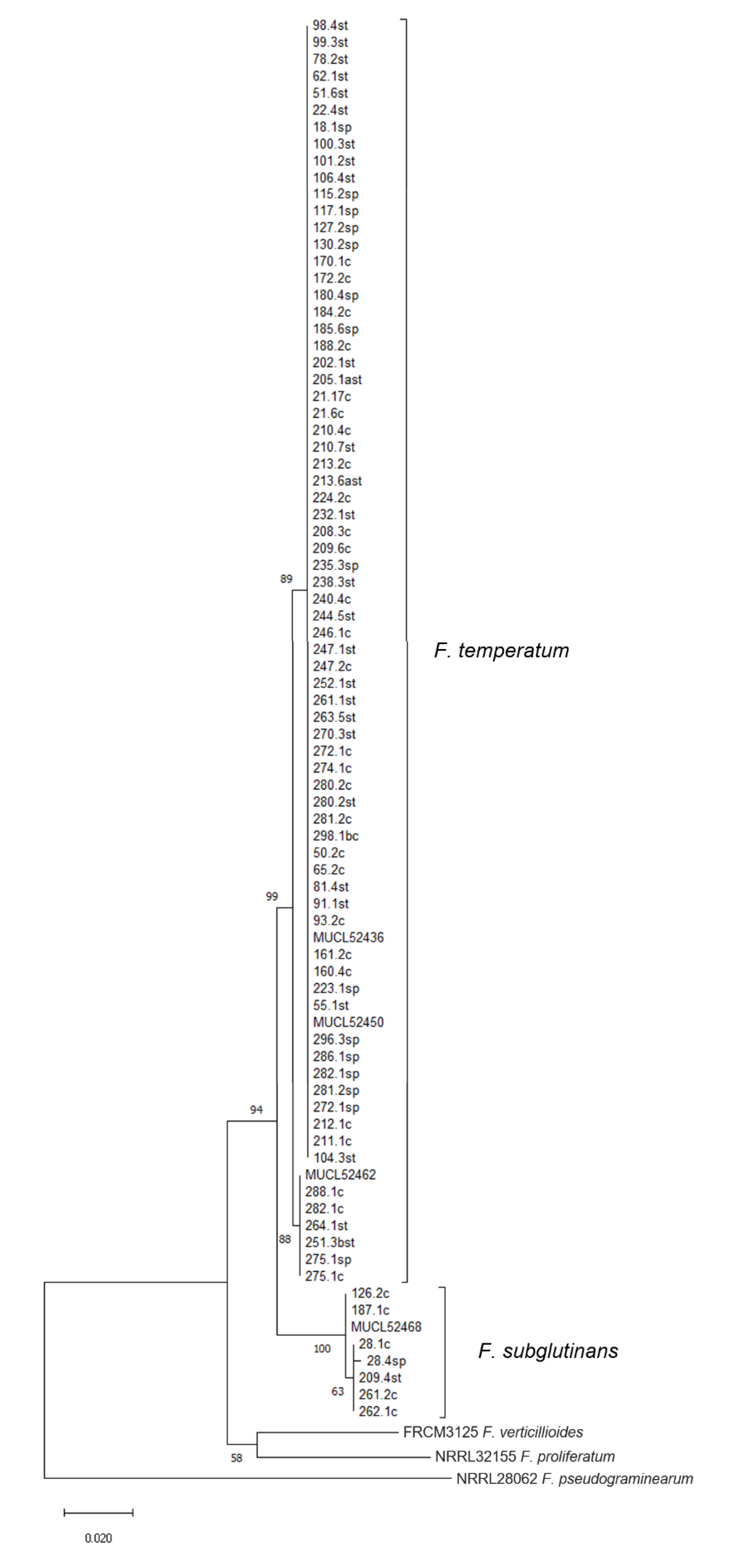
10]. Phylogenetic analysis enabled the assignment of all obtained isolates to their respective species, as supported by bootstrap values. Separation of isolates and references suggest a rather uniform population of
F. temperatum, when compared with phylogenetic investigations recently published [
9,
34,
36,
54,
55,
56]. The analysis showed that the isolates of
F. temperatum are genetically divided into two groups, as supported by a high bootstrap values. This has also been reported by Shin et al. [
55] for isolates of
F. temperatum from Korea, even though the isolates were obtained from a single location. We were unable to link this grouping to morphological characteristics nor to mycotoxin profiles of the respective isolates. The analysis of
TEF-1α is highly recommended for taxonomical identification of species in the genus
Fusarium but reliable separation shall be verified by the investigation of additional informative loci [
57]. Hence, we selected the gene for the DNA-directed RNA polymerase II subunit (
RPB2) for verification of distinct groups that we observed in the analysis of
TEF-1α. The formation of clades could not be reproduced with
RPB2 amplified according to Lofgren et al. [
58], using a 763 bp portion of the gene located at the 5′ end. The associated phylogram, based on multiple sequence alignments for a small set of isolates and reference strains, is provided in
Figure S2. Therefore, we assume that the grouping of nucleotide sequences of
TEF-1α does not reflect the genetic relatedness among the strains.
In order to evaluate the risk of mycotoxin contamination upon infection with
F. temperatum and
F. subglutinans, in vitro cultures were screened for the presence of eight mycotoxins (BEA, MON, FA, FUSA, ENNB, ENNA1, FB1, and FB2), selected according to Scauflaire et al. [
19] and others [
14,
17,
18]. We detected BEA in 58 cultures exclusively of
F. temperatum, supporting the suitability of BEA production for the separation of
F. temperatum from
F. subglutinans, similarly as BEA production separated
F. verticillioides from
F. proliferatum [
24]. The production of MON, FA, and FUSA was confirmed in almost all tested cultures of
F. temperatum and
F. subglutinans. Previous studies suggested that MON may not be produced universally by
F. temperatum [
19]. Based on our results, both
F. temperatum and
F. subglutinans produced MON but the amounts varied among isolates. The production of MON and FA by some isolates was so low that it could escape detection. As we found only single isolates showing low production of MON and FA, further studies need to be conducted to clarify this finding. The amounts of FA were low, as reported from other studies [
59]. Even though FA exerts low toxicity at levels normally detected in natural infections, synergistic effects have been reported between DON and FA in pigs and FB1 and FA in chicken eggs [
60]. Although DON is not produced by the species of the FFSC, both DON and FB1 are common in maize grains and contamination with multiple mycotoxins may occur. FA thus potentially increases the risk of mycotoxin exposure via maize consumption.
In the present study, all strains of
F. temperatum and
F. subglutinans were FUSA-producers. Even though contamination with FUSA and also BEA are rarely reported in literature [
61], a significant role of these toxins in the natural toxicity of the producing species, also in association with other toxins, such as MON, was suggested [
16]. The biological activity of FUSA remains to be fully elucidated. We were not able to detect ENNB or ENNA1 in any analyzed rice culture; however, low amounts of ENNB were detected in three maize cobs naturally infected with
F. temperatum, harvested in 2017 (data not shown). These maize cobs were co-infected with
F. temperatum and
F. avenaceum, which was likely responsible for the production of ENNB [
6]. Production of any enniatins has shown to be a rare event among isolates of
F. temperatum so it may not be considered a mycotoxin characteristic of
F. temperatum [
19]. Even though the ability to synthesize enniatins is a common feature of some trichothecene producing species of
Fusarium, such as
F. avenaceum, [
6,
62] it has rarely been observed for any species of the GFSC [
13]. Enniatins are less toxic than trichothecenes, such as deoxynivalenol. Their function in pathogenesis on maize is still unknown.
The production of fumonisins has been reported in a few cases for both
F. temperatum and
F. subglutinans, even though the associated
FUM-cluster could not be detected in their genomes. The production of these toxins by both species in maize plants [
35] can be explained by spontaneous infection of the plants with other species. Wang at al. [
22] reported production of fumonisins by
F. temperarum but because they analyzed grains from the field rather than axenic cultures and have not described the analytical method adequately, their results have not been considered here. We detected fumonisins in only one culture of
F. temperatum, likely due to contamination, supporting the current view that
F. temperatum does not produce fumonisins. FB1 and FB2 were also detected in one culture of
F. subglutinans. Even though
F. subglutinans was occasionally reported to produce fumonisins [
63], which is a common feature among members of the FFSC, both classical [
18] as well as modern studies [
64,
65,
66,
67] convincingly showed that
F. subglutinans does not produce fumonisins. Mycotoxin production found in a small set of cultures of
F. verticillioides and
F. proliferatum confirmed the established mycotoxin spectra of these species [
59,
68,
69,
70].
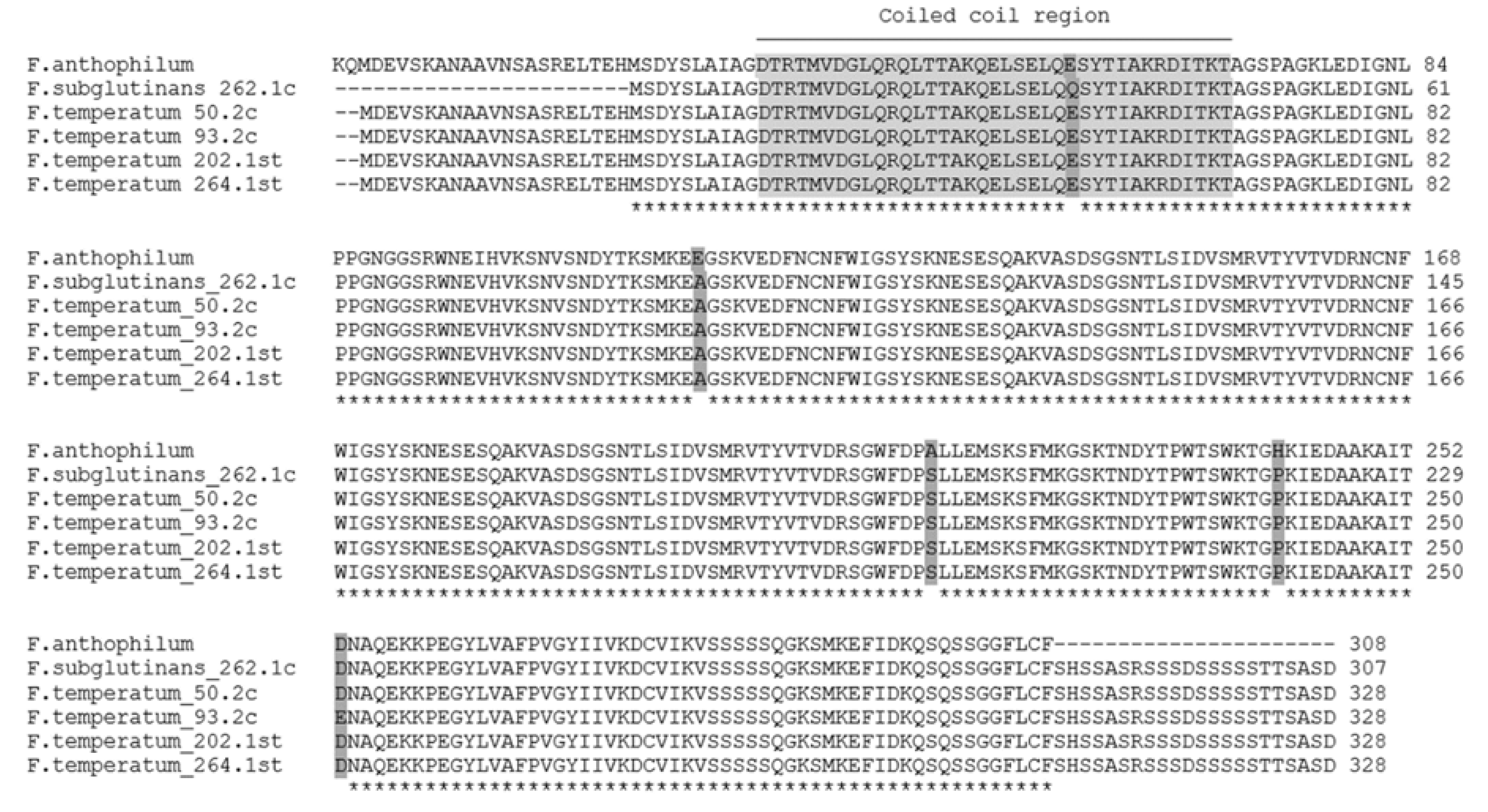
Gene clusters required for synthesis of sphinganine-analog metabolites, such as fumonisins, in
Fusarium spp. are conserved [
65,
66,
67]. Sequence analysis of the gene amplified from
F. subglutinans and
F. temperatum with primers for the gene
FUM1 revealed that the product was unrelated to
FUM1. Interestingly, a gene with a high sequence similarity to the product was found in the genome of the closely related species
F. anthophilum. The amino acid sequence predicted a coiled coil region (
Figure 2), possibly indicating involvement of a hypothetical protein in the regulation of gene expression.
F. anthophilum is a member of the American clade of the FFSC, which includes fumonisin-nonproducing species
F. temperatum and
F. subglutinans and fumonisin-producing species
F. anthophilum and
Fusarium bulbicola [
65]. The authors assume a combination of loss of the respective genes during species divergence and horizontal gene transfer, leading to the loss or retention of fumonisin synthesis.
The results obtained in the present study indicate a high degree of variability in BEA, MON, FA, and FUSA production among isolates of F. temperatum. We found isolates with a comparably low toxicity, producing low amounts of FA, BEA, MON, and FUSA, and highly toxic isolates. Our results support the assumption of lower toxigenic risk due to infection of maize with F. subglutinans as compared to F. temperatum, especially regarding the production of BEA.
In conclusion, the present investigations indicate that F. temperatum occurs more frequently on maize cobs and is more aggressive than previously known, and thus represents an elevated threat of food and feed contamination to growers, processing industries, and consumers. In addition, F. temperatum may enhance the risk of head blight on wheat if grown in rotation with maize.



 and
and 



{kind=link}
{kind=link}
{kind=link}