
Is Toxoplasma gondii a Trigger of Bipolar Disorder?




Abstract
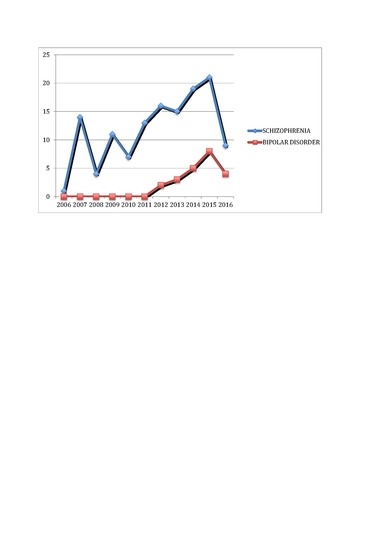
:
1. Introduction
2. Relationship between Toxoplasma gondii and Bipolar Disorder
3. The Role of T. gondii in the Etiopathogenesis of Psychiatric Disorders
4. The Association between Autoimmune/Inflammatory Dysregulation, Infectious Agents and Risk of Bipolar Disorder
5. Anti-Toxoplasmic Activity of Antipsychotics and Mood Stabilizers
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Judd, L.L.; Akiskal, H.S.; Schettler, P.J.; Endicott, J.; Leon, A.C.; Solomon, D.A.; Coryell, W.; Maser, J.D.; Keller, M.B. Psychosocial disability in the course of bipolar I and II disorders: A prospective, comparative, longitudinal study. Arch. Gen. Psychiatry 2005, 62, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.Y.; Patel, V.; Joestl, S.S.; March, D.; Insel, T.R.; Daar, A.S. Grand challenges in global mental health. Nature 2011, 475, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Whiteford, H.A.; Degenhardt, L.; Rehm, J.; Baxter, A.J.; Ferrari, A.J.; Erskine, H.E.; Charlson, F.J.; Norman, R.E.; Flaxman, A.D.; Johns, N.; et al. Global burden of disease attributable to mental and substance use disorders: Findings from the Global Burden of Disease Study 2010. Lancet 2013, 382, 1575–1586. [Google Scholar] [CrossRef]
- Gershon, E.S.; Alliey-Rodriguez, N.; Liu, C. After GWAS: Searching for genetic risk for schizophrenia and bipolar disorder. Am. J. Psychiatry 2011, 168, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Kerner, B. Genetics of bipolar disorder. Appl. Clin. Genet. 2014, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.M.; Scully, P.; Scott, L.V.; Dinan, T.G. Cytokine profiles in bipolar affective disorder: Focus on acutely ill patients. J. Affect. Disord. 2006, 90, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Stallings, C.; Origoni, A.; Boronow, J.; Yolken, R. Elevated serum levels of C-reactive protein are associated with mania symptoms in outpatients with bipolar disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Domínguez, A.; Hernández, M.E.; Berlanga, C.; Gutiérrez-Mora, D.; Moreno, J.; Heinze, G.; Pavón, L. Immune variations in bipolar disorder: Phasic differences. Bipolar. Disord. 2007, 9, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, N.; Doukhan, R.; Kurtlucan, O.; Tamouza, R.; Leboyer, M. Immunity, inflammation, and bipolar disorder: Diagnostic and therapeutic implications. Curr. Psychiatry Rep. 2013, 15, 387. [Google Scholar] [CrossRef] [PubMed]
- Canetta, S.E.; Bao, Y.; Co, M.D.; Ennis, F.A.; Cruz, J.; Terajima, M.; Shen, L.; Kellendonk, C.; Schaefer, C.A.; Brown, A.S. Serological documentation of maternal influenza exposure and bipolar disorder in adult offspring. Am. J. Psychiatry 2014, 171, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Muneer, A. Bipolar Disorder: Role of Inflammation and the Development of Disease Biomarkers. Psychiatry Investig. 2016, 13, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, J.D.; McIntyre, R.S. Bipolar Disorder and Inflammation. Psychiatr. Clin. N. Am. 2016, 39, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Remlinger-Molenda, A.; Wojciak, P.; Michalak, M.; Karczewski, J.; Rybakowski, J.K. Selected cytokine profiles during remission in bipolar patients. Neuropsychobiology 2012, 66, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Stallings, C.; Origoni, A.; Vaughan, C.; Katsafanas, E.; Khushalani, S.; Yolken, R. A combined marker of inflammation in individuals with mania. PLoS ONE 2013, 8, e73520. [Google Scholar] [CrossRef] [PubMed]
- Yolken, R.H.; Torrey, E.F. Are some cases of psychosis caused by microbial agents? A review of the evidence. Mol. Psychiatry 2008, 13, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Yolken, R.H.; Dickerson, F.B.; Torrey, E.F. Toxoplasma and schizophrenia. Parasite Immunol. 2009, 31, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Derkits, E.J. Prenatal infection and schizophrenia: A review of epidemiologic and translational studies. Am. J. Psychiatry 2010, 167, 261–280. [Google Scholar] [CrossRef] [PubMed]
- Arias, I.; Sorlozano, A.; Villegas, E.; de Dios Luna, J.; McKenney, K.; Cervilla, J.; Gutierrez, B.; Gutierrez, J. Infectious agents associated with schizophrenia: A meta-analysis. Schizophr. Res. 2012, 136, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Torrey, E.F.; Bartko, J.J.; Yolken, R.H. Toxoplasma gondii and other risk factors for schizophrenia: An update. Schizophr. Bull. 2012, 38, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, N.; Daban-Huard, C.; Lajnef, M.; Richard, J.R.; Delavest, M.; Godin, O.; Le Guen, E.; Vederine, F.E.; Lépine, J.P.; Jamain, S.; et al. Relationship between Toxoplasma gondii infection and bipolar disorder in a French sample. J. Affect. Disord. 2013, 148, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Sutterland, A.L.; Fond, G.; Kuin, A.; Koeter, M.W.; Lutter, R.; van Gool, T.; Yolken, R.; Szoke, A.; Leboyer, M.; de Haan, L. Beyond the association. Toxoplasma gondii in schizophrenia, bipolar disorder, and addiction: Systematic review and meta-analysis. Acta Psychiatr. Scand. 2015, 132, 161–179. [Google Scholar] [PubMed]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef] [PubMed]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk factors for Toxoplasma gondii infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.M.; Dubey, J.P. Toxoplasmosis: A history of clinical observations. Int. J. Parasitol. 2009, 39, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Bahia-Oliveira, L.M.; Jones, J.L.; Azevedo-Silva, J.; Alves, C.C.; Oréfice, F.; Addiss, D.G. Highly endemic, waterborne toxoplasmosis in north Rio de Janeiro state, Brazil. Emerg. Infect. Dis. 2003, 9, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Bamne, M.; Wood, J.; Chowdari, K.; Watson, A.M.; Celik, C.; Mansour, H.; Klei, L.; Gur, R.C.; Bradford, L.D.; Calkins, M.E.; et al. Evaluation of HLA polymorphisms in relation to schizophrenia risk and infectious exposure. Schizophr. Bull. 2012, 38, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Remington, J.S.; McLeod, R.; Thulliez, P.; Desmonts, G. Toxoplasmosis. In Infectious Diseases of the Fetus and Newborn Infant, 5th ed.; Remington, J.S., Klein, J., Eds.; WB Saunders: Philadelphia, PA, USA, 2001; pp. 205–346. [Google Scholar]
- Kaye, A. Toxoplasmosis: Diagnosis, treatment, and prevention in congenitally exposed infants. J. Pediatr. Health Care 2011, 25, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Hong, J. Microglia and inflammation-mediated neurodegeneration: Multiple triggers with a common mechanism. Prog. Neurobiol. 2005, 76, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Bergink, V.; Gibney, S.M.; Drexhage, H.A. Autoimmunity, inflammation, and psychosis: A search for peripheral markers. Biol. Psychiatry 2014, 75, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Havlícek, J.; Gasová, Z.G.; Smith, A.P.; Zvára, K.; Flegr, J. Decrease of psychomotor performance in subjects with latent ‘asymptomatic’ toxoplasmosis. Parasitology 2001, 122, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J.; Zitková, S.; Kodym, P.; Frynta, D. Induction of changes in human behaviour by the parasitic protozoan Toxoplasma gondii. Parasitology 1996, 113, 49–54. [Google Scholar]
- Flegr, J.; Preiss, M.; Klose, J.; Havlícek, J.; Vitáková, M.; Kodym, P. Decreased level of psychobiological factor novelty seeking and lower intelligence in men latently infected with the protozoan parasite Toxoplasma gondii Dopamine, a missing link between schizophrenia and toxoplasmosis? Biol. Psychol. 2003, 63, 253–268. [Google Scholar] [CrossRef]
- Hinze-Selch, D.; Däubener, W.; Erdag, S.; Wilms, S. The diagnosis of a personality disorder increases the likelihood for seropositivity to Toxoplasma gondii in psychiatric patients. Folia Parasitol. 2010, 57, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J. Effects of Toxoplasma on human behavior. Schizophr. Bull. 2007, 33, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J. Influence of latent Toxoplasma infection on human personality, physiology and morphology: Pros and cons of the Toxoplasma-human model in studying the manipulation hypothesis. J. Exp. Biol. 2013, 216, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Cook, T.B.; Brenner, L.A.; Cloninger, C.R.; Langenberg, P.; Igbide, A.; Giegling, I.; Hartmann, A.M.; Konte, B.; Friedl, M.; Brundin, L.; et al. “Latent” infection with Toxoplasma gondii: Association with trait aggression and impulsivity in healthy adults. J. Psychiatr. Res. 2015, 60, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Novotná, M.; Hanusova, J.; Klose, J.; Preiss, M.; Havlicek, J.; Roubalová, K.; Flegr, J. Probable neuroimmunological link between Toxoplasma and cytomegalovirus infections and personality changes in the human host. BMC. Infect. Dis. 2005, 5, 54. [Google Scholar]
- Berdoy, M.; Webster, J.P.; Macdonald, D.W. Fatal attraction in rats infected with Toxoplasma gondii. Proc. Biol. Sci. 2000, 267, 1591–1594. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A.; Kim, S.K.; Giacomini, N.; Boothroyd, J.C.; Sapolsky, R.M. Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc. Natl. Acad. Sci. USA 2007, 104, 6442–6447. [Google Scholar] [CrossRef] [PubMed]
- Gulinello, M.; Acquarone, M.; Kim, J.H.; Spray, D.C.; Barbosa, H.S.; Sellers, R.; Tanowitz, H.B.; Weiss, L.M. Acquired infection with Toxoplasma gondii in adult mice results in sensorimotor deficits but normal cognitive behavior despite widespread brain pathology. Microbes. Infect. 2010, 12, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Gatkowska, J.; Wieczorek, M.; Dziadek, B.; Dzitko, K.; Dlugonska, H. Behavioral changes in mice caused by Toxoplasma gondii invasion of brain. Parasitol. Res. 2012, 111, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A. Mechanisms of Host Behavioral Change in Toxoplasma gondii Rodent Association. PLoS Pathog. 2015, 11, e1004935. [Google Scholar] [CrossRef] [PubMed]
- Amminger, G.P.; McGorry, P.D.; Berger, G.E.; Wade, D.; Yung, A.R.; Phillips, L.J.; Harrigan, S.M.; Francey, S.M.; Yolken, R.H. Antibodies to infectious agents in individuals at ultra-high risk for psychosis. Biol. Psychiatry 2007, 61, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, S.; Pinto, B.; Bonuccelli, U.; Bruschi, F. Neurobiological studies on the relationship between toxoplasmosis and neuropsychiatric diseases. J. Neurol. Sci. 2015, 351, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Arling, T.A.; Yolken, R.H.; Lapidus, M.; Langenberg, P.; Dickerson, F.B.; Zimmerman, S.A.; Balis, T.; Cabassa, J.A.; Scrandis, D.A.; Tonelli, L.H.; et al. Toxoplasma gondii antibody titers and history of suicide attempts in patients with recurrent mood disorders. J. Nerv. Ment. Dis. 2009, 197, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Ling, V.J.; Lester, D.; Mortensen, P.B.; Langenberg, P.W.; Postolache, T.T. Toxoplasma gondii seropositivity and suicide rates in women. J. Nerv. Ment. Dis. 2011, 199, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Okusaga, O.; Langenberg, P.; Sleemi, A.; Vaswani, D.; Giegling, I.; Hartmann, A.M.; Konte, B.; Friedl, M.; Groer, M.W.; Yolken, R.H.; et al. Toxoplasma gondii antibody titers and history of suicide attempts in patients with schizophrenia. Schizophr. Res. 2011, 133, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.G.; Mortensen, P.B.; Norgaard-Pedersen, B.; Postolache, T.T. Toxoplasma gondii infection and self-directed violence in mothers. Arch. Gen. Psychiatry 2012, 69, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Esquivel, C.; Sánchez-Anguiano, L.F.; Arnaud-Gil, C.A.; López-Longoria, J.C.; Molina-Espinoza, L.F.; Estrada-Martínez, S.; Liesenfeld, O.; Hernández-Tinoco, J.; Sifuentes-Álvarez, A.; Salas-Martínez, C. Toxoplasma gondii infection and suicide attempts: A case-control study in psychiatric outpatients. J. Nerv. Ment. Dis. 2013, 201, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Torrey, E.F.; Bartko, J.J.; Lun, Z.R.; Yolken, R.H. Antibodies to Toxoplasma gondii in patients with schizophrenia: A meta-analysis. Schizophr. Bull. 2007, 33, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Stallings, C.; Origoni, A.; Katsafanas, E.; Schweinfurth, L.; Savage, C.; Khushalani, S.; Yolken, R. Antibodies to Toxoplasma gondii and cognitive functioning in schizophrenia, bipolar disorder, and nonpsychiatric controls. J. Nerv. Ment. Dis. 2014, 202, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, N.; Daban-Huard, C.; Lajnef, M.; Gadel, R.; Le Corvoisier, P.; Delavest, M.; Carde, S.; Lépine, J.P.; Jamain, S.; Houenou, J.; et al. Cognitive deterioration among bipolar disorder patients infected by Toxoplasma gondii is correlated to interleukin 6 levels. J. Affect. Disord. 2015, 179, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Pearce, B.D.; Kruszon-Moran, D.; Jones, J.L. The relationship between Toxoplasma gondii infection and mood disorders in the third National Health and Nutrition Survey. Biol. Psychiatry 2012, 72, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Delgado García, G.; Rodríguez Perdomo, E. Reactivity of toxoplasmin intradermal test in neurotic and manic-depressive patients. Rev. Cubana Med. Trop. 1980, 32, 35–39. [Google Scholar] [PubMed]
- Tedla, Y.; Shibre, T.; Ali, O.; Tadele, G.; Woldeamanuel, Y.; Asrat, D.; Aseffa, A.; Mihret, W.; Abebe, M.; Alem, A. Serum antibodies to Toxoplasma gondii and Herpesvidae family viruses in individuals with schizophrenia and bipolar disorder: A case-control study. Ethiop. Med. J. 2011, 49, 211–220. [Google Scholar] [PubMed]
- Cetinkaya, Z.; Yazar, S.; Gecici, O.; Namli, M.N. Anti-Toxoplasma gondii antibodies in patients with schizophrenia--preliminary findings in a Turkish sample. Schizophr. Bull. 2007, 33, 789–791. [Google Scholar] [CrossRef] [PubMed]
- Hamidinejat, H.; Ghorbanpoor, M.; Hosseini, H.; Alavi, S.M.; Nabavi, L.; Jalali, M.H.; Borojeni, M.P.; Jafari, H.; Mohammadaligol, S. Toxoplasma gondii infection in first-episode and inpatient individuals with schizophrenia. Int. J. Infect. Dis. 2010, 14, e978–e981. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Stallings, C.; Origoni, A.; Vaughan, C.; Katsafanas, E.; Khushalani, S.; Yolken, R. Antibodies to Toxoplasma gondii in individuals with mania. Bipolar. Disord. 2014, 16, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.R.; Beckie, T.M.; Brenner, L.A.; Beckstead, J.W.; Seyfang, A.; Postolache, T.T.; Groer, M.W. Relationship between Toxoplasma gondii and Mood Disturbance in Women Veterans. Mil. Med. 2015, 180, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Khademvatan, S.; Khajeddin, N.; Izadi, S.; Saki, J. Study of Toxoplasma gondii Infection in Patients with Bipolar Disorder. J. Med. Sci. 2013, 13, 215–220. [Google Scholar]
- Del Grande, C.; Contini, C.; Schiavi, E.; Rutigliano, G.; Maritati, M.; Seraceni, S.; Pinto, B.; Dell’Osso, L.; Bruschi, F. Bipolar Disorder With Psychotic Features and Ocular Toxoplasmosis. A Possible Pathogenetic Role of the Parasite? J. Nerv. Ment. Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Nørgaard-Pedersen, B.; Waltoft, B.L.; Sørensen, T.L.; Hougaard, D.; Yolken, R.H. Early infections of Toxoplasma gondii and the later development of schizophrenia. Schizophr. Bull. 2007, 33, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Blomström, Å.; Karlsson, H.; Gardner, R.; Jörgensen, L.; Magnusson, C.; Dalman, C. Associations Between Maternal Infection During Pregnancy, Childhood Infections, and the Risk of Subsequent Psychotic Disorder--A Swedish Cohort Study of Nearly 2 Million Individuals. Schizophr. Bull. 2016, 42, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Schaefer, C.A.; Quesenberry, C.P.; Liu, L., Jr.; Babulas, V.P.; Susser, E.S. Maternal exposure to toxoplasmosis and risk of schizophrenia in adult offspring. Am. J. Psychiatry 2005, 162, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Pedersen, C.B.; McGrath, J.J.; Hougaard, D.M.; Nørgaard-Petersen, B.; Mors, O.; Børglum, A.D.; Yolken, R.H. Neonatal antibodies to infectious agents and risk of bipolar disorder: A population-based case-control study. Bipolar. Disord. 2011, 13, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Freedman, D.; Bao, Y.; Shen, L.; Schaefer, C.A.; Brown, A.S. Maternal T. gondii, offspring bipolar disorder and neurocognition. Psychiatry Res. 2016, 243, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Buka, S.L.; Cannon, T.D.; Suzuki, J.; Viscidi, R.P.; Torrey, E.F.; Yolken, R.H. Serological pattern consistent with infection with type I Toxoplasma gondii in mothers and risk of psychosis among adult offspring. Microbes Infect. 2009, 11, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Coccaro, E.F.; Lee, R.; Groer, M.W.; Can, A.; Coussons-Read, M.; Postolache, T.T. Toxoplasma gondii infection: Relationship with aggression in psychiatric subjects. Clin. Psychiatry 2016, 77, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Lester, D. Predicting European suicide rates with physiological indices. Psychol. Rep. 2010, 107, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Träskman-Bendz, L.; Janelidze, S.; Langenberg, P.; Saleh, A.; Constantine, N.; Okusaga, O.; Bay-Richter, C.; Brundin, L.; Postolache, T.T. Toxoplasma gondii immunoglobulin G antibodies and nonfatal suicidal self-directed violence. J. Clin. Psychiatry 2012, 73, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Wang, G.H.; Li, Q.Y.; Shu, C.; Jiang, M.S.; Guo, Y. Prevalence of Toxoplasma infection in first-episode schizophrenia and comparison between Toxoplasma-seropositive and Toxoplasma-seronegative schizophrenia. Acta Psychiatr. Scand. 2006, 114, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Halonen, S.K.; Lyman, W.D.; Chiu, F.C. Growth and development of Toxoplasma gondii in human neurons and astrocytes. J. Neuropathol. Exp. Neurol. 1996, 55, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.R.; Pariante, C.M.; Everall, I.P. Glial cell abnormalities in major psychiatric disorders: The evidence and implications. Brain Res. Bull. 2001, 55, 585–595. [Google Scholar] [CrossRef]
- Konradt, C.; Ueno, N.; Christian, D.A.; Delong, J.H.; Pritchard, G.H.; Herz, J.; Bzik, D.J.; Koshy, A.A.; McGavern, D.B.; Lodoen, M.B.; et al. Endothelial cells are a replicative niche for entry of Toxoplasma gondii to the central nervous system. Nat. Microbiol. 2016, 1, 16001. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.C.; Langoni, H. Toxoplasma gondii: Host-parasite interaction and behavior manipulation. Parasitol. Res. 2009, 105, 893e8. [Google Scholar] [CrossRef] [PubMed]
- Courret, N.; Darche, S.; Sonigo, P.; Milon, G.; Buzoni-Gâtel, D.; Tardieux, I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006, 107, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Melzer, T.C.; Cranston, H.J.; Weiss, L.M.; Halonen, S.K. Host Cell Preference of Toxoplasma gondii Cysts in Murine Brain: A Confocal Study. J. Neuroparasitol. 2010, 1, N100505. [Google Scholar] [CrossRef] [PubMed]
- Berenreiterová, M.; Flegr, J.; Kuběna, A.A.; Němec, P. The distribution of Toxoplasma gondii cysts in the brain of a mouse with latent toxoplasmosis: Implications for the behavioral manipulation hypothesis. PLoS ONE 2011, 6, e28925. [Google Scholar] [CrossRef] [PubMed]
- McConkey, G.A.; Martin, H.L.; Bristow, G.C.; Webster, J.P. Toxoplasma gondii infection and behaviour—location, location, location? J. Exp. Biol. 2013, 216, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Sapolsky, R.M.; Vyas, A. Toxoplasma gondii infection induces dendritic retraction in basolateral amygdala accompanied by reduced corticosterone secretion. Dis. Model. Mech. 2013, 6, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Ferreira, L.R.; Alsaad, M.; Verma, S.K.; Alves, D.A.; Holland, G.N.; McConkey, G.A. Experimental Toxoplasmosis in Rats Induced Orally with Eleven Strains of Toxoplasma gondii of Seven Genotypes: Tissue Tropism, Tissue Cyst Size, Neural Lesions, Tissue Cyst Rupture without Reactivation, and Ocular Lesions. PLoS ONE 2016, 11, e0156255. [Google Scholar] [CrossRef] [PubMed]
- Coccaro, E.F.; Sripada, C.S.; Yanowitch, R.N.; Phan, K.L. Corticolimbic function in impulsive aggressive behavior. Biol. Psychiatry 2011, 69, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Hermes, G.; Ajioka, J.W.; Kelly, K.A.; Mui, E.; Roberts, F.; Kasza, K.; Mayr, T.; Kirisits, M.J.; Wollmann, R.; Ferguson, D.J.; et al. Neurological and behavioral abnormalities, ventricular dilatation, altered cellular functions, inflammation, and neuronal injury in brains of mice due to common, persistent, parasitic infection. J. Neuroinflamm. 2008, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Torrey, E.F.; Yolken, R.H. Toxoplasma gondii and schizophrenia. Emerg. Infect. Dis. 2003, 9, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Coccaro, E.F.; Lee, R.; Kavoussi, R.J. Aggression, suicidality, and intermittent explosive disorder: Serotonergic correlates in personality disorder and healthy control subjects. Neuropsychopharmacology 2010, 35, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Coccaro, E.F.; Lee, R.; Vezina, P. Cerebrospinal fluid glutamate concentration correlates with impulsive aggression in human subjects. J. Psychiatr. Res. 2013, 47, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J. How and why Toxoplasma makes us crazy. Trends Parasitol. 2013, 29, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Prandovszky, E.; Gaskell, E.; Martin, H.; Dubey, J.P.; Webster, J.P.; McConkey, G.A. The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS ONE 2011, 6, e23866. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, E.A.; Smith, J.E.; Pinney, J.W.; Westhead, D.R.; McConkey, G.A. A unique dual activity amino acid hydroxylase in Toxoplasma gondii. PLoS ONE 2009, 4, e4801. [Google Scholar] [CrossRef] [PubMed]
- Strobl, J.S.; Goodwin, D.G.; Rzigalinski, B.A.; Lindsay, D.S. Dopamine stimulates propagation of Toxoplasma gondii tachyzoites in human fibroblast and primary neonatal rat astrocyte cell cultures. J. Parasitol. 2012, 98, 1296–1299. [Google Scholar] [CrossRef] [PubMed]
- Stibbs, H.H. Changes in brain concentrations of catecholamines and indoleamines in Toxoplasma gondii infected mice. Ann. Trop. Med. Parasitol. 1985, 79, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, S.; Pinto, B.; Bruschi, F. Toxoplasmosis and neuropsychiatric disease: Can serological studies establish a clear relationship? Neurol. Sci. 2013, 34, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.; Chan, M.K.; Bahn, S. The potential of immune biomarkers to advance personalized medicine approaches for schizophrenia. J. Nerv. Ment. Dis. 2015, 203, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Mattei, D.; Westrin, Å.; Träskman-Bendz, L.; Brundin, L. Cytokine levels in the blood may distinguish suicide attempters from depressed patients. Brain. Behav. Immun. 2011, 25, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Janelidze, S.; Hagell, P.; Erhardt, S.; Samuelsson, M.; Minthon, L.; Hansson, O.; Björkqvist, M.; Träskman-Bendz, L.; Brundin, L. Interleukin-6 is elevated in the cerebrospinal fluid of suicide attempters and related to symptom severity. Biol. Psychiatry 2009, 66, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Munkholm, K.; Braüner, J.V.; Kessing, L.V.; Vinberg, M. Cytokines in bipolar disorder vs. healthy control subjects: A systematic review and meta-analysis. J. Psychiatr. Res. 2013, 47, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, J.D.; Cha, D.S.; Mansur, R.B.; McIntyre, R.S. Inflamed moods: A review of the interactions between inflammation and mood disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 53, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Reichenberg, A.; Yirmiya, R.; Schuld, A.; Kraus, T.; Haack, M.; Morag, A.; Pollmächer, T. Cytokine-associated emotional and cognitive disturbances in humans. Arch. Gen. Psychiatry 2001, 58, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends. Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Harrison, N.A.; Brydon, L.; Walker, C.; Gray, M.A.; Steptoe, A.; Critchley, H.D. Inflammation causes mood changes through alterations in subgenual cingulate activity and mesolimbic connectivity. Biol. Psychiatry 2009, 66, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Eisenberger, N.I.; Berkman, E.T.; Inagaki, T.K.; Rameson, L.T.; Mashal, N.M.; Irwin, M.R. Inflammation-induced anhedonia: Endotoxin reduces ventral striatum responses to reward. Biol. Psychiatry 2010, 68, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Brietzke, E.; Stertz, L.; Fernandes, B.S.; Kauer-Sant’anna, M.; Mascarenhas, M.; Escosteguy Vargas, A.; Chies, J.A.; Kapczinski, F. Comparison of cytokine levels in depressed, manic and euthymic patients with bipolar disorder. J. Affect. Disord. 2009, 116, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Söderlund, J.; Olsson, S.K.; Samuelsson, M.; Walther-Jallow, L.; Johansson, C.; Erhardt, S.; Landén, M.; Engberg, G. Elevation of cerebrospinal fluid interleukin-1ß in bipolar disorder. J. Psychiatry Neurosci. 2011, 36, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.M.; Su, T.P.; Tsai, S.J.; Wen-Fei, C.; Li, C.T.; Pei-Chi, T.; Mu-Hong, C. Comparison of inflammatory cytokine levels among type I/type II and manic/hypomanic/euthymic/depressive states of bipolar disorder. J. Affect. Disord. 2014, 166, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Boufidou, F.; Nikolaou, C.; Alevizos, B.; Liappas, I.A.; Christodoulou, G.N. Cytokine production in bipolar affective disorder patients under lithium treatment. J. Affect. Disord. 2004, 82, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Jung, H.G.; Myint, A.M.; Kim, H.; Park, S.H. Imbalance between pro-inflammatory and anti-inflammatory cytokines in bipolar disorder. J. Affect. Disord. 2007, 104, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Guloksuz, S.; Cetin, E.A.; Cetin, T.; Deniz, G.; Oral, E.T.; Nutt, D.J. Cytokine levels in euthymic bipolar patients. J. Affect. Disord. 2010, 126, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.I.; Kemp, D.E.; Soczynska, J.K.; McIntyre, R.S. Inflammation and the phenomenology, pathophysiology, comorbidity, and treatment of bipolar disorder: A systematic review of the literature. J. Clin. Psychiatry 2009, 70, 1078–1790. [Google Scholar] [CrossRef] [PubMed]
- Kapczinski, F.; Kauer-Sant’Anna, M.; Andreazza, A.C.; Bond, D.J.; Lam, R.W.; Young, L.T.; Yatham, L.N. Brain-derived neurotrophic factor and inflammatory markers in patients with early- vs. late-stage bipolar disorder. Int. J. Neuropsychopharmacol. 2009, 12, 447–458. [Google Scholar]
- Hinze-Selch, D.; Däubener, W.; Eggert, L.; Erdag, S.; Stoltenberg, R.; Wilms, S. A controlled prospective study of Toxoplasma gondii infection in individuals with schizophrenia: Beyond seroprevalence. Schizophr. Bull. 2007, 33, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M.; Schwarz, M.J.; Verkerk, R.; Mueller, H.H.; Zach, J.; Scharpé, S.; Steinbusch, H.W.; Leonard, B.E.; Kim, Y.K. Reversal of imbalance between kynurenic acid and 3-hydroxykynurenine by antipsychotics in medication-naïve and medication-free schizophrenic patients. Brain. Behav. Immun. 2011, 25, 1576–1581. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.L.; Llenos, I.C.; Dulay, J.R.; Weis, S. Upregulation of the initiating step of the kynurenine pathway in postmortem anterior cingulate cortex from individuals with schizophrenia and bipolar disorder. Brain Res. 2006, 16, 1073–1074. [Google Scholar] [CrossRef] [PubMed]
- Olsson, S.K.; Samuelsson, M.; Saetre, P.; Lindström, L.; Jönsson, E.G.; Nordin, C.; Engberg, G.; Erhardt, S.; Landén, M. Elevated levels of kynurenic acid in the cerebrospinal fluid of patients with bipolar disorder. J. Psychiatry Neurosci. 2010, 35, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.S.; Owe-Larsson, B.; Asp, L.; Kocki, T.; Adler, M.; Hetta, J.; Gardner, R.; Lundkvist, G.B.; Urbanska, E.M.; Karlsson, H. Activation of kynurenine pathway in ex vivo fibroblasts from patients with bipolar disorder or schizophrenia: Cytokine challenge increases production of 3-hydroxykynurenine. J. Psychiatr. Res. 2013, 47, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.; Kumar, V.; Hari Dass, S.A.; Vyas, A. Toxoplasma gondii infection enhances testicular steroidogenesis in rats. Mol. Ecol. 2013, 22, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J.; Lindová, J.; Kodym, P. Sex-dependent toxoplasmosis-associated differences in testosterone concentration in humans. Parasitology 2008, 135, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Bullmore, E.T.; Harrison, P. Is there a flame in the brain in psychosis? Biol. Psychiatry 2014, 75, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Benros, M.E.; Nielsen, P.R.; Nordentoft, M.; Eaton, W.W.; Dalton, S.O.; Mortensen, P.B. Autoimmune diseases and severe infections as risk factors for schizophrenia: A 30-year population-based register study. Am. J. Psychiatry 2011, 168, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Benros, M.E.; Eaton, W.W.; Mortensen, P.B. The epidemiologic evidence linking autoimmune diseases and psychosis. Biol. Psychiatry 2014, 75, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Monji, A.; Kato, T.A.; Mizoguchi, Y.; Horikawa, H.; Seki, Y.; Kasai, M.; Yamauchi, Y.; Yamada, S.; Kanba, S. Neuroinflammation in schizophrenia especially focused on the role of microglia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 42, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U. Developmental neuroinflammation and schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 42, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F.; Reutfors, J.; Osby, U.; Ekbom, A.; Montgomery, S.M. Coeliac disease and risk of mood disorders--a general population-based cohort study. J. Affect. Disord. 2007, 99, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Benros, M.E.; Waltoft, B.L.; Nordentoft, M.; Ostergaard, S.D.; Eaton, W.W.; Krogh, J.; Mortensen, P.B. Autoimmune diseases and severe infections as risk factors for mood disorders: A nationwide study. JAMA Psychiatry 2013, 70, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Avramopoulos, D.; Pearce, B.D.; McGrath, J.; Wolyniec, P.; Wang, R.; Eckart, N.; Hatzimanolis, A.; Goes, F.S.; Nestadt, G.; Mulle, J.; et al. Infection and inflammation in schizophrenia and bipolar disorder: A genome wide study for interactions with genetic variation. PLoS ONE 2015, 10, e0116696. [Google Scholar] [CrossRef] [PubMed]
- De Berardis, D.; Conti, C.M.; Campanella, D.; Carano, A.; Scali, M.; Valchera, A.; Serroni, N.; Pizzorno, A.M.; D’Albenzio, A.; Fulcheri, M.; et al. Evaluation of C-reactive protein and total serum cholesterol in adult patients with bipolar disorder. Int. J. Immunopathol. Pharmacol. 2008, 21, 319–324. [Google Scholar] [PubMed]
- Singh, B.; Chaudhuri, T.K. Role of C-reactive protein in schizophrenia: An overview. Psychiatry Res. 2014, 216, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Kazma, R.; Le Floch, E.; Bennabi, M.; Hamdani, N.; Bengoufa, D.; Dahoun, M.; Manier, C.; Bellivier, F.; Krishnamoorthy, R.; et al. Toxoplasma gondii exposure may modulate the influence of TLR2 genetic variation on bipolar disorder: A gene-environment interaction study. Int. J. Bipolar. Disord. 2016, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Schröder, N.W.; Schumann, R.R. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious disease. Lancet Infect. Dis. 2005, 5, 156–164. [Google Scholar] [CrossRef]
- van Well, G.T.; Sanders, M.S.; Ouburg, S.; Kumar, V.; van Furth, A.M.; Morré, S.A. Single nucleotide polymorphisms in pathogen recognition receptor genes are associated with susceptibility to meningococcal meningitis in a pediatric cohort. PLoS ONE 2013, 8, e64252. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Busson, M.; Etain, B.; Jamain, S.; Hamdani, N.; Boukouaci, W.; Amokrane, K.; Bennabi, M.; Le Guen, E.; Dargél, A.A.; et al. Polymorphism of Toll-like receptor 4 gene in bipolar disorder. J. Affect. Disord. 2014, 152–154, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Hamdani, N.; Busson, M.; Etain, B.; Bennabi, M.; Amokrane, K.; Boukouaci, W.; Fortier, C.; Marzais, F.; Bengoufa, D.; et al. Association between toll-like receptor 2 gene diversity and early-onset bipolar disorder. J. Affect. Disord. 2014, 165, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Hamdani, N.; Etain, B.; Bennabi, M.; Boukouaci, W.; Amokrane, K.; Fortier, C.; Marzais, F.; Bengoufa, D.; Bellivier, F.; et al. Genetic association between a ‘standing’ variant of NOD2 and bipolar disorder. Immunobiology 2014, 219, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Etain, B.; Lajnef, M.; Hamdani, N.; Bennabi, M.; Bengoufa, D.; Sundaresh, A.; Chaabane, A.B.; Bellivier, F.; Henry, C.; et al. Combined effect of TLR2 gene polymorphism and early life stress on the age at onset of bipolar disorders. PLoS ONE 2015, 10, e0119702. [Google Scholar] [CrossRef] [PubMed]
- Leweke, F.M.; Gerth, C.W.; Koethe, D.; Klosterkötter, J.; Ruslanova, I.; Krivogorsky, B.; Torrey, E.F.; Yolken, R.H. Antibodies to infectious agents in individuals with recent onset schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2004, 254, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Jones-Brando, L.; Torrey, E.F.; Yolken, R.H. Drugs used in the treatment of schizophrenia and bipolar disorder inhibit the replication of Toxoplasma gondii. Schizophr. Res. 2003, 62, 237–244. [Google Scholar] [CrossRef]
- Webster, J.P.; Lamberton, P.H.; Donnelly, C.A.; Torrey, E.F. Parasites as causative agents of human affective disorders? The impact of anti-psychotic, mood-stabilizer and anti-parasite medication on Toxoplasma gondii’s ability to alter host behaviour. Proc. Biol. Sci. 2006, 273, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Pezzella, N.; Bouchot, A.; Bonhomme, A.; Pingret, L.; Klein, C.; Burlet, H.; Balossier, G.; Bonhomme, P.; Pinon, J.M. Involvement of calcium and calmodulin in Toxoplasma gondii tachyzoite invasion. Eur. J. Cell Biol. 1997, 74, 92–101. [Google Scholar] [PubMed]
- Johannessen, C.U. Mechanisms of action of valproate: A commentatory. Neurochem. Int. 2000, 37, 103–110. [Google Scholar] [CrossRef]
- Fond, G.; Macgregor, A.; Tamouza, R.; Hamdani, N.; Meary, A.; Leboyer, M.; Dubremetz, J.F. Comparative analysis of anti-toxoplasmic activity of antipsychotic drugs and valproate. Eur. Arch. Psychiatry Clin. Neurosci. 2014, 264, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, D.G.; Strobl, J.S.; Lindsay, D.S. Evaluation of five antischizophrenic agents against Toxoplasma gondii in human cell cultures. J. Parasitol. 2011, 97, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, D.G.; Strobl, J.; Mitchell, S.M.; Zajac, A.M.; Lindsay, D.S. Evaluation of the mood-stabilizing agent valproic acid as a preventative for toxoplasmosis in mice and activity against tissue cysts in mice. J. Parasitol. 2008, 94, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Boyer, L.; Gaman, A.; Laouamri, H.; Attiba, D.; Richard, J.R.; Delavest, M.; Houenou, J.; Le Corvoisier, P.; Charron, D.; et al. Treatment with anti-toxoplasmic activity (TATA) for Toxoplasma positive patients with bipolar disorders or schizophrenia: A cross-sectional study. J. Psychiatr. Res. 2015, 63, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Selle, V.; Schalkwijk, S.; Vázquez, G.H.; Baldessarini, R.J. Treatments for acute bipolar depression: Meta-analyses of placebo-controlled, monotherapy trials of anticonvulsants, lithium and antipsychotics. Pharmacopsychiatry 2014, 47, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Musetti, L.; Del Grande, C.; Marazziti, D.; Dell’Osso, L. Treatment of bipolar depression. CNS Spectr. 2013, 18, 177–187. [Google Scholar] [CrossRef] [PubMed]
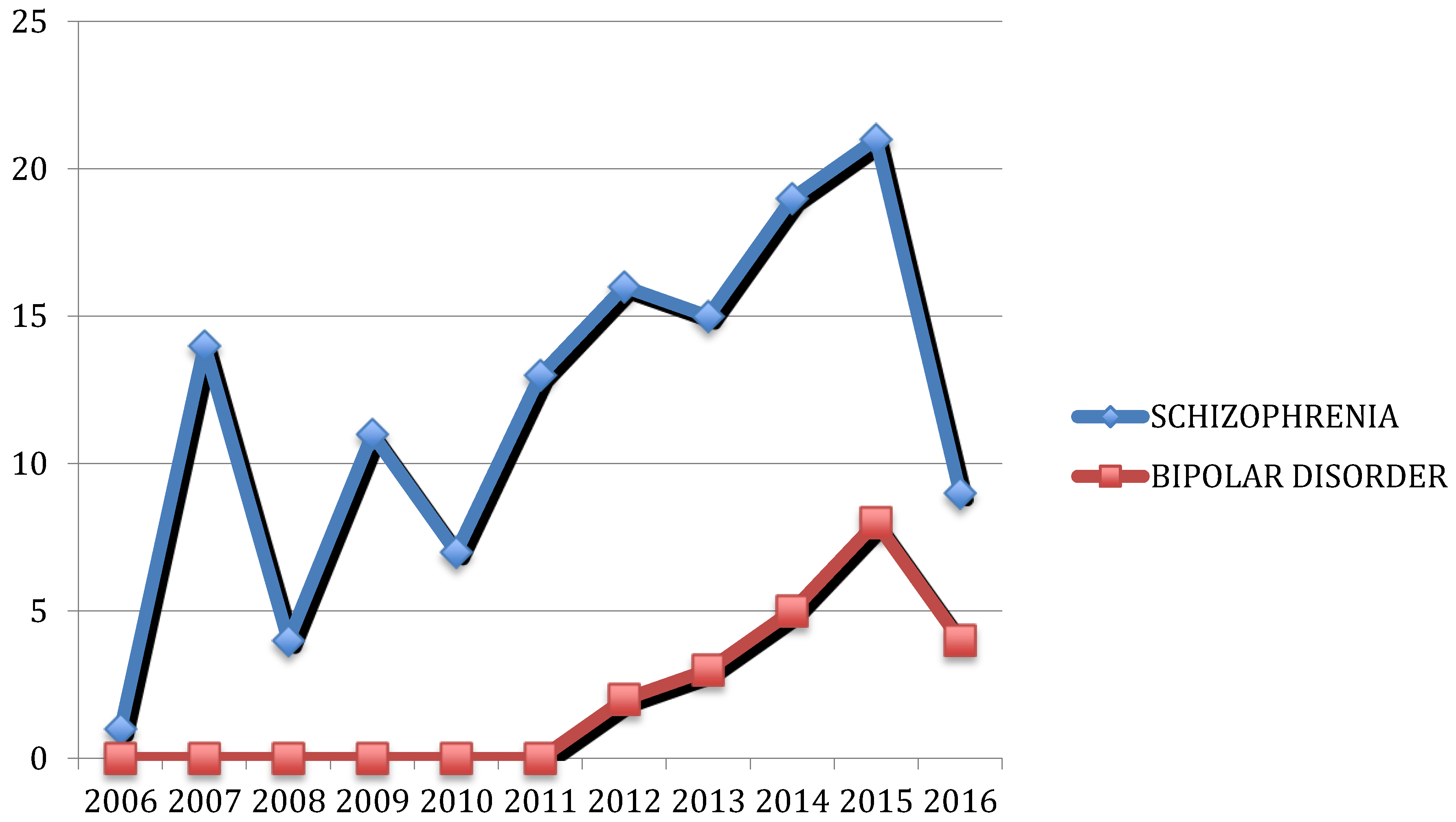

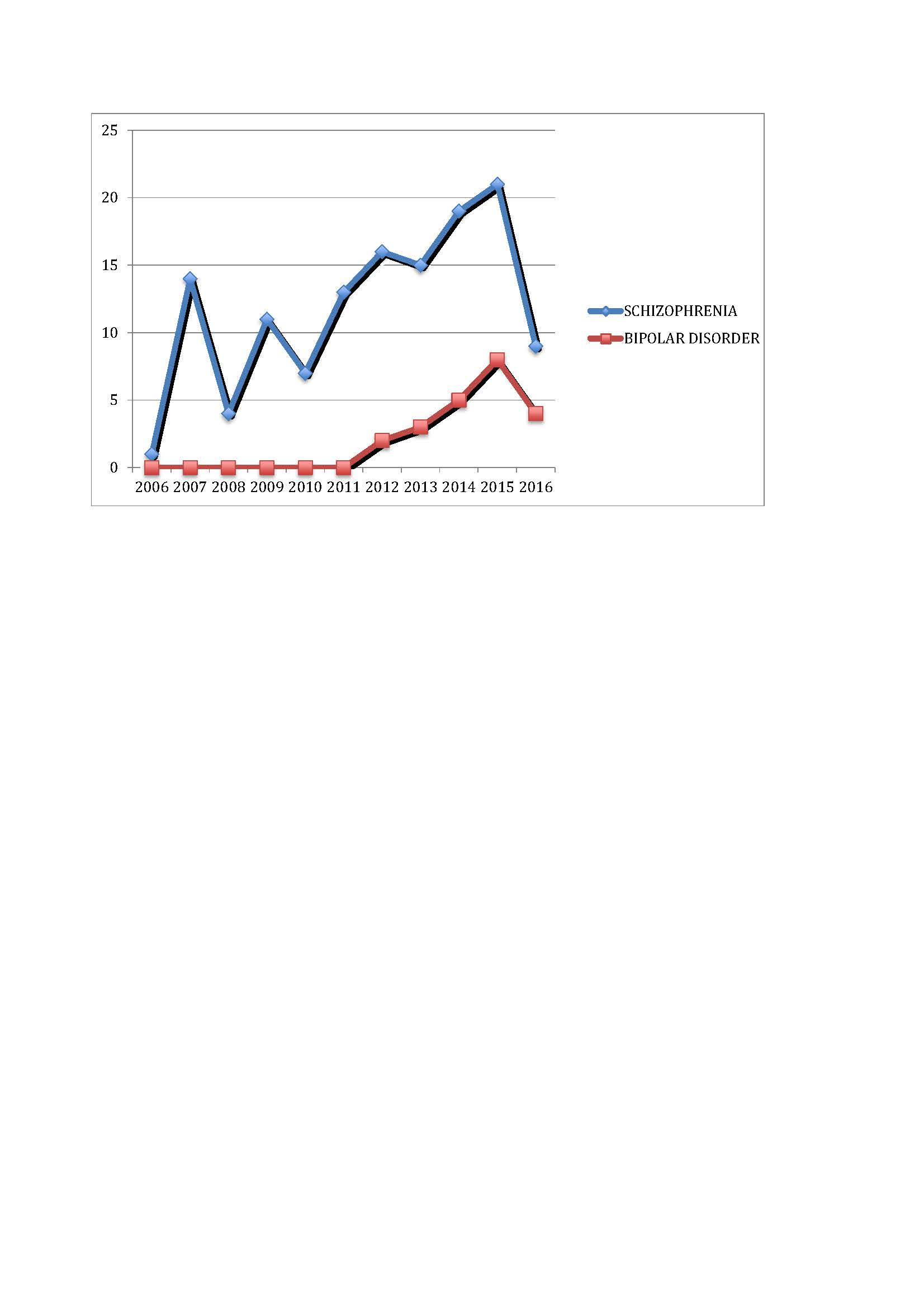{kind=link}
{kind=link}
| Authors (Year), Country | Study Design | Toxo Determination | Sample Size | Description of Association | Limitations |
|---|---|---|---|---|---|
| Delgado Garcia, Rodriguez Perdomo (1980) [57], Cuba | Case-control study | Toxoplasmin intradermal test | 270 individuals (50 pts with manic-depressive psychosis, 120 neurotics, 100 HC) | The highest percentage of reactors was found among pts with manic-depressive psychosis (66.0%); the intensity of reaction was also higher among these pts. | Small sample size |
| Hinze-Selch et al. (2010) [36], Germany | Cross-sectional study | IgG levels | 1110 (270 SCH, 465 MD, 67 schizoaffective disorder, 87 BD, 214 HC) | Additional diagnosis of a personality disorder is significantly associated with T. gondii infection among all psychiatric inpatients. | Small sample size of BD pts |
| Tedla et al. (2011) [58], Ethiopia | Case-control study | IgG levels | 495 (214 SCH, 171 BD, 71 HC) | Higher T. gondii seroprevalence in individuals with SCH (OR = 4.7) and BD (OR = 3). | No IgM determination |
| Pearce et al. (2012) [56], USA | Cross-sectional study | IgG levels | 7440 (individuals 15–39 years old from the Third National Health and Nutrition Examination Survey) | Significant relationship between Toxoplasma seropositivity and lifetime history of a BD I subtype with manic and depressive episodes (adjusted OR = 2.4, p < 0.05). No statistically-significant association between T. gondii seroprevalence and a history of MD or dysthymia. | Small number of respondents with both BD I and positive serology for T. gondii; no IgM determination |
| Hamdani et al. (2013) [20], France | Case-control study | IgM/IgG levels | 216 (110 BD, 106 HC) | BD pts had significantly higher seropositivity to T. gondii and higher anti-T. gondii IgG titers as compared to HC (76.9% vs. 48.2%, p = 0.00045; 3.06 ± 1.07 vs. 2.07 ± 1.9, p = 0.002). IgM antibodies detection was negative for the whole sample. Seropositive participants had a 3.6-fold odds risk of BD as compared to seronegative group (p < 0.0001) (adjusted OR = 2.40, p = 0.026). | Small sample size |
| Dickerson et al. (2014) [61], USA | Cross-sectional study | IgM/IgG levels assessed up to three time points | 1114 (57 individual with mania, 743 other psychiatric groups, 314 non-psychiatric controls) | Significantly elevated levels of anti-T. gondii IgM in pts with mania as compared to non-psychiatric controls (OR = 2.33, p < 0.04 at hospital admission; OR = 2.32, p < 0.02 at study entry during the hospital stay). Significant difference between the prevalences of increased levels of anti-T. gondii IgM at the baseline and the follow-up time-point (t = 2.97, p < 0.003). Significantly elevated levels of IgG antibodies to T. gondii (OR = 2.42, p < 0.04) in the recent onset psychosis group alone. | Time of T. gondii infection is unknown; clinical assessments only during the hospital stay; lack of measurement of symptom scores at hospital admission |
| Khademvatan et al. (2013) [63], Iran | Cross-sectional study | IgM/IgG levels | 317 (117 BD I; 200 HC) | No significant difference in prevalence of anti-T. gondii IgG antibodies between BD I pts and HC (31.62% vs. 26.5%, p = 0.3). No significant difference between IgM levels in the two groups. | No BD II pts |
| Duffy et al. (2015) [62], USA | Cross-sectional study | IgM/IgG levels | 70 (veteran women) | Positive relationships between T. gondii and depressive symptoms, anger, confusion and overall mood disturbance. | Small sample size |
| Authors (Year), Country | Study Design | Toxo Determination | Sample Size | Description of Association | Limitations |
|---|---|---|---|---|---|
| Xiao et al. (2009) [70], USA | Case-control study | IgG levels and serotyping | 837 (219 sera from pregnant women whose children developed SCH and affective psychotic illnesses in adult life and 618 matched unaffected control mothers) | No association between maternal antibodies to T. gondii and risk of psychoses in the offspring. Only the offspring of mothers with a serological pattern consistent with Toxoplasma type I infection are at significantly increased risk for the development of psychosis (p = 0.03). The elevated risk for subsequent psychoses associated with maternal infection with the type I strain is statistically significant for patients with affective psychosis (p = 0.005), but not for those with schizophrenic and other non-affective psychosis. | Moderate sensitivity of serotyping; unknown indicators of the timing of maternal infection |
| Mortensen et al. (2011) [68], Denmark | Case-control study | IgG levels (neonatal dried blood spots) | 154 (127 BD pts and 127 matched controls) | No association between marker of T. gondii prenatal infection and risk of BD. | Small study samples; absence of information about specific serotypes of Toxoplasma; unknown indicators of the timing of maternal infection |
| Freedman et al. (2016) [69], USA | Case-control study | IgG levels (maternal sera) | 255 (85 BD cases and 170 comparison subjects) | Maternal T. gondii IgG is not associated with the risk of BD in offspring. | Small sample size; absence of information about specific serotypes of Toxoplasma; unable to determine when the infection occurred in relation to the pregnancy or birth |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Grande, C.; Galli, L.; Schiavi, E.; Dell’Osso, L.; Bruschi, F. Is Toxoplasma gondii a Trigger of Bipolar Disorder? Pathogens 2017, 6, 3. https://doi.org/10.3390/pathogens6010003
Del Grande C, Galli L, Schiavi E, Dell’Osso L, Bruschi F. Is Toxoplasma gondii a Trigger of Bipolar Disorder? Pathogens. 2017; 6(1):3. https://doi.org/10.3390/pathogens6010003
Chicago/Turabian StyleDel Grande, Claudia, Luca Galli, Elisa Schiavi, Liliana Dell’Osso, and Fabrizio Bruschi. 2017. "Is Toxoplasma gondii a Trigger of Bipolar Disorder?" Pathogens 6, no. 1: 3. https://doi.org/10.3390/pathogens6010003
APA StyleDel Grande, C., Galli, L., Schiavi, E., Dell’Osso, L., & Bruschi, F. (2017). Is Toxoplasma gondii a Trigger of Bipolar Disorder? Pathogens, 6(1), 3. https://doi.org/10.3390/pathogens6010003