
Establishment and Partial Characterization of Three Novel Permanent Cell Lines Originating from European Freshwater Fish Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Establishment of the Primary Cells
2.2. Subcultivation and Storage of the Cells
2.3. Growth Characteristics Investigations
2.4. Molecular Investigations
2.5. Viral Susceptibility Assays
2.6. Titration of the Viruses
3. Results
3.1. Establishment of the Cell Lines
3.2. Growth Characteristics
3.3. Molecular Study
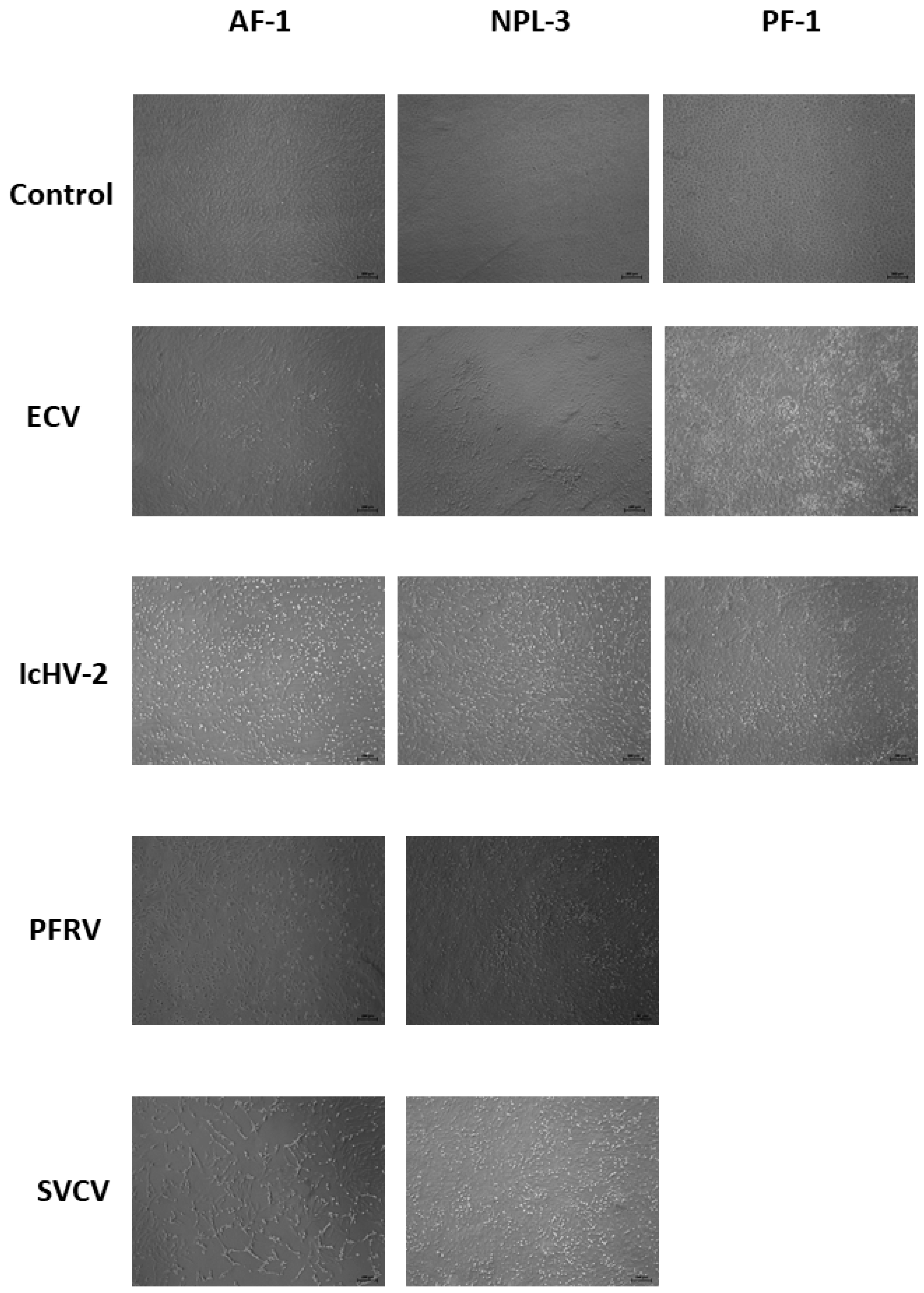
3.4. Cell Line Susceptibility Assays and Virus Titrations
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeong, Y.J.; Kim, K.I. A New Cell Line Derived from the Caudal Fin of the Dwarf Gourami (Trichogaster lalius) and Its Susceptibility to Fish Viruses. Biology 2023, 12, 829. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Zhang, Q.; Wang, N.; Wang, L.; Chen, Z.; Yan, L.; Yan, K.; Hu, G.; Chen, S. Establishment and characterization of cell line from fin of Chinense bahaba (Bahaba taipingensis) and transcriptome analysis of its response to megalocytivirus SKIV-SD strain infection. Aquaculture 2024, 592, 741108. [Google Scholar] [CrossRef]
- Meng, X.Y.; Wang, Z.H.; Yu, X.D.; Zhang, Q.Y.; Ke, F. Development and characterization of a skin cell line from Chinese perch (Siniperca chuatsi) and its application in aquatic animal viruses. J. Fish. Dis. 2022, 45, 1439–1449. [Google Scholar] [CrossRef]
- Jeffery, K.R.; Bateman, K.; Bayley, A.; Feist, S.W.; Hulland, J.; Longshaw, C.; Stone, D.; Woolford, G.; Way, K. Isolation of a cyprinid herpesvirus 2 from goldfish, Carassius auratus (L.), in the UK. J. Fish. Dis. 2007, 30, 649–656. [Google Scholar] [CrossRef]
- Dharmaratnam, A.; Kumar, R.; Valaparambil, B.S.; Sood, N.; Pradhan, P.K.; Das, S.; Swaminathan, T.R. Establishment and characterization of fantail goldfish fin (FtGF) cell line from goldfish. PeerJ 2020, 8, e9373. [Google Scholar] [CrossRef] [PubMed]
- Noguera, P.A.; Grunow, B.; Klinger, M.; Lester, K.; Collet, B.; Del-Pozo, J. Atlantic salmon cardiac primary cultures: An in vitro model to study viral host pathogen interactions and pathogenesis. PLoS ONE 2017, 12, e0181058. [Google Scholar] [CrossRef]
- Zhang, H.; Sridhar, A.; Delrez, N.; He, B.; Fourny, S.; Gao, Y.; Donohoe, O.; Vanderplasschen, A.F.C. Development Using Bioluminescence Imaging of a Recombinant Anguillid Herpesvirus 1 Vaccine Candidate Associated with Normal Replication In Vitro but Abortive Infection In Vivo. Vaccines 2024, 12, 1423. [Google Scholar] [CrossRef]
- Boutier, M.; Ronsmans, M.; Ouyang, P.; Fournier, G.; Reschner, A.; Rakus, K.; Wilkie, G.S.; Farnir, F.; Bayrou, C.; Lieffrig, F.; et al. Rational development of an attenuated recombinant cyprinid herpesvirus 3 vaccine using prokaryotic mutagenesis and in vivo bioluminescent imaging. PLoS Pathog. 2015, 11, e1004690. [Google Scholar] [CrossRef]
- Goswami, M.; Yashwanth, B.S.; Trudeau, V.; Lakra, W.S. Role and relevance of fish cell lines in advanced in vitro research. Mol. Biol. Rep. 2022, 49, 2393–2411. [Google Scholar] [CrossRef]
- Lakra, W.S.; Swaminathan, T.R.; Joy, K.P. Development, characterization, conservation and storage of fish cell lines: A review. Fish. Physiol. Biochem. 2011, 37, 1–20. [Google Scholar] [CrossRef]
- Thangaraj, R.; Narendrakumar, L.; Geetha, P.; Shanmuganathan, A.; Dharmaratnam, A.; Nithianantham, S. Comprehensive update on inventory of finfish cell lines developed during the last decade (2010–2020). Rev. Aquac. 2021, 13, 2248–2288. [Google Scholar] [CrossRef]
- Wolf, K.; Mann, J.A. Poikilotherm vertebrate cell lines and viruses: A current listing for fishes. In Vitro 1980, 16, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, J.; Han, S.; Wang, Y.; Chen, L. Establishment and characterization of a fin cell line from Takifugu obscurus and its application to fish immunology. J. Fish. Biol. 2025, 106, 1134–1147. [Google Scholar] [CrossRef]
- Hedrick, R.; Gilad, O.; Yun, S.; Spangenberg, J.; Marty, G.; Nordhausen, R.; Kebus, M.; Bercovier, H.; Eldar, A. A herpesvirus associated with mass mortality of juvenile and adult koi, a strain of common carp. J. Aquat. Anim. Health 2000, 12, 44–57. [Google Scholar] [CrossRef]
- Fijan, N.; Sulimanovic, D.; Bearzotti, M.; Muzinic, D.; Zwillenberg, L.; Chilmonczyk, S.; Vautherot, J.; Dekinkelin, P. Some properties of the Epithelioma-papulosum-cyprini (EPC) cell-line from carp CYPRINUS-CARPIO. Ann. Virol. 1983, 134, 207–220. [Google Scholar] [CrossRef]
- Hedrick, R.; McDowell, T.; Rosemark, R.; Aronstein, D.; Lannan, C. 2 cell-lines from white sturgeon. Trans. Am. Fish. Soc. 1991, 120, 528–534. [Google Scholar] [CrossRef]
- Shchelkunova, T.; Kolbassova, Y.; IS, S. Fish virus susceptibility of continuous cell lines established from Siberian sturgeon tissues. Tsitologiya 2006, 48, 814–815. [Google Scholar]
- Jing, H.; Lin, X.; Xu, L.; Gao, L.; Zhang, M.; Wang, N.; Wu, S. Establishment and characterization of a heart-derived cell line from goldfish (Carassius auratus). Fish. Physiol. Biochem. 2017, 43, 977–986. [Google Scholar] [CrossRef]
- Puechmaille, S.J.; Wibbelt, G.; Korn, V.; Fuller, H.; Forget, F.; Mühldorfer, K.; Kurth, A.; Bogdanowicz, W.; Borel, C.; Bosch, T.; et al. Pan-European distribution of white-nose syndrome fungus (Geomyces destructans) not associated with mass mortality. PLoS ONE 2011, 6, e19167. [Google Scholar] [CrossRef]
- Hopple, J.S.; Vilgalys, R. Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: Divergent domains, outgroups, and monophyly. Mol. Phylogenet Evol. 1999, 13, 1–19. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Fehér, E.; Doszpoly, A.; Horváth, B.; Marton, S.; Forró, B.; Farkas, S.L.; Bányai, K.; Juhász, T. Whole genome sequencing and phylogenetic characterization of brown bullhead (Ameiurus nebulosus) origin ranavirus strains from independent disease outbreaks. Infect. Genet. Evol. 2016, 45, 402–407. [Google Scholar] [CrossRef]
- Juhasz, T.; Woynarovichne, L.; Csaba, G.; Farkas, L.; Dan, A. Isolation of ranavirus causing mass mortality in brown bullheads (Ameiurus nebulosus) in Hungary. Magy. Allatorvosok Lapja 2013, 135, 763–768. [Google Scholar]
- Alborali, L.; Bovo, G.; Lavazza, A.; Capellaro, H.; Guadagnini, P.F. Isolation of a herpesvirus in breeding catfish (Ictalurus melas). Bull. Eur. Assoc. Fish. Pathol. 1996, 16, 134–137. [Google Scholar]
- Borzák, R.; Haluk, T.; Bartha, D.; Doszpoly, A. Complete genome sequence and analysis of ictalurid herpesvirus 2. Arch. Virol. 2018, 163, 1083–1085. [Google Scholar] [CrossRef]
- Doszpoly, A.; Shaalan, M.; El-Matbouli, M. Silver Nanoparticles Proved to Be Efficient Antivirals In Vitro against Three Highly Pathogenic Fish Viruses. Viruses 2023, 15, 1689. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.; Muench, H. A Simple Method of Estimating Fifty Percent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Kirsner, R.; Margolis, D.; Baldursson, B.; Petursdottir, K.; Davidsson, O.; Weir, D.; Lantis, J. Fish skin grafts compared to human amnion/chorion membrane allografts: A double-blind, prospective, randomized clinical trial of acute wound healing. Wound Repair Regen. 2020, 28, 75–80. [Google Scholar] [CrossRef]
- Benjaminson, M.; Gilchriest, J.; Lorenz, M. In vitro edible muscle protein production system (MPPS): Stage 1, fish. Acta Astronaut. 2002, 51, 879–889. [Google Scholar] [CrossRef]
- Groeneveld, L.F.; Gregusson, S.; Guldbrandtsen, B.; Hiemstra, S.J.; Hveem, K.; Kantanen, J.; Lohi, H.; Stroemstedt, L.; Berg, P. Domesticated Animal Biobanking: Land of Opportunity. PLoS Biol. 2016, 14, e1002523. [Google Scholar] [CrossRef]
- Ahne, W. Fish cell-culture—Fibroblastic line (PG) from ovaries of juvenile pike (Esox lucius). In Vitr. -J. Tissue Cult. Assoc. 1979, 15, 839–840. [Google Scholar] [CrossRef]
- Abonyi, F.; Varga, A.; Sellyei, B.; Eszterbauer, E.; Doszpoly, A. Juvenile Wels Catfish (Silurus glanis) Display Age-Related Mortality to European Catfish Virus (ECV) under Experimental Conditions. Viruses 2022, 14, 1832. [Google Scholar] [CrossRef] [PubMed]
- Ahne, W.; Schlotfeldt, H.; Thomsen, I. Fish Viruses—Isolation of an icosahedral cytoplasmic deoxyribovirus from sheatfish (Silurus glanis). J. Vet. Med. Ser. B-Zentralblatt Fur Vet. Reihe B-Infect. Dis. Vet. Public Health 1989, 36, 333–336. [Google Scholar] [CrossRef]
- Bigarre, L.; Cabon, J.; Baud, M.; Pozet, F.; Castric, J. Ranaviruses associated with high mortalities in catfish in France. Bull. Eur. Assoc. Fish Pathol. 2008, 28, 163–168. [Google Scholar]
- Pozet, F.; Morand, M.; Moussa, A.; Torhy, C.; Dekinkelin, P. Isolation and preliminary characterization of a pathogenic icosahedral deoxyribovirus from the catfish Ictalurus melas. Dis. Aquat. Org. 1992, 14, 35–42. [Google Scholar] [CrossRef]
- Bloch, B.; Larsen, J. An iridovirus-like agent aqssociated with systemic infection in cultured turbot Scophtalmus maximus fry in Denmark. Dis. Aquat. Org. 1993, 15, 235–240. [Google Scholar] [CrossRef]
- Tapiovaara, H.; Olesen, N.J.; Lindén, J.; Rimaila-Pärnänen, E.; von Bonsdorff, C.H. Isolation of an iridovirus from pike-perch Stizostedion lucioperca. Dis. Aquat. Organ. 1998, 32, 185–193. [Google Scholar] [CrossRef]
- Langdon, J.; Humphrey, J.; Williams, L.; Hyatt, A.; Westbury, H.A. First virus isolation from Australian fish: An iridovirus-like pathogen from redfin perch, Perca fluviatilis L. J. Fish. Dis. 1986, 9, 263–268. [Google Scholar] [CrossRef]
- Ahne, W.; Bjorklund, H.V.; Essbauer, S.; Fijan, N.; Kurath, G.; Winton, J.R. Spring viremia of carp (SVC). Dis. Aquat. Organ. 2002, 52, 261–272. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kelly, R.K.; Nielsen, O. Epidermal hyperplasias of northern pike (Esox lucius) associated with herpesvirus and C-type particles. Arch. Virol. 1984, 79, 255–272. [Google Scholar] [CrossRef]
- Coffee, L.L.; Casey, J.W.; Bowser, P.R. Pathology of tumors in fish associated with retroviruses: A review. Vet. Pathol. 2013, 50, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Leskisenoja, K.; Macrae, R.; Hawley, L.M.; Subramaniam, K.; Waltzek, T.B.; Richard, J.; Josefsson, C.; Valtonen, E.T. An alloherpesvirus infection of European perch Perca fluviatilis in Finland. Dis. Aquat. Organ. 2018, 128, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Sellyei, B.; Baska, F.; Varga, Á.; Borzák, R.; Doszpoly, A. Molecular detection of a novel cyprinid herpesvirus in roach (Rutilus rutilus) and asp (Leuciscus aspius) showing typical signs of carp pox disease. Arch. Virol. 2020, 165, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Tarján, Z.L.; Pénzes, J.J.; Tóth, R.P.; Benkő, M. First detection of circovirus-like sequences in amphibians and novel putative circoviruses in fishes. Acta Vet. Hung. 2014, 62, 134–144. [Google Scholar] [CrossRef]
- Dorson, M.; Torchy, C.; Chilmonczyk, S.; de Kinkelin, P. A rhabdovirus pathogenic for perch, Perca fluviatilis L.: Isolation and preliminary study. J. Fish. Dis. 1984, 7, 241–245. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| SVCV | PFRV | ECV | IcHV-2 | |
|---|---|---|---|---|
| EPC | 7.81 × 107 | 1.05 × 107 | 3.16 × 105 | 1.39 × 105 |
| AF-1 | 1.86 × 106 * | 1.63 × 107 | 4.39 × 104 * | 4.39 × 105 |
| NPL-3 | 6.58 × 106 * | 7.81 × 105 | 3.16 × 106 * | 1.86 × 107 |
| PF-1 | - | - | 4.39 × 105 | 4.39 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doszpoly, A. Establishment and Partial Characterization of Three Novel Permanent Cell Lines Originating from European Freshwater Fish Species. Pathogens 2025, 14, 531. https://doi.org/10.3390/pathogens14060531
Doszpoly A. Establishment and Partial Characterization of Three Novel Permanent Cell Lines Originating from European Freshwater Fish Species. Pathogens. 2025; 14(6):531. https://doi.org/10.3390/pathogens14060531
Chicago/Turabian StyleDoszpoly, Andor. 2025. "Establishment and Partial Characterization of Three Novel Permanent Cell Lines Originating from European Freshwater Fish Species" Pathogens 14, no. 6: 531. https://doi.org/10.3390/pathogens14060531
APA StyleDoszpoly, A. (2025). Establishment and Partial Characterization of Three Novel Permanent Cell Lines Originating from European Freshwater Fish Species. Pathogens, 14(6), 531. https://doi.org/10.3390/pathogens14060531