
Serological Evidence for Circulation of Influenza D Virus in the Ovine Population in Italy

 , , , ,
, , , ,  , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Reference Viruses
2.3. Serum Samples
2.4. HI Assay
2.5. VN Assay
2.6. Data Analysis
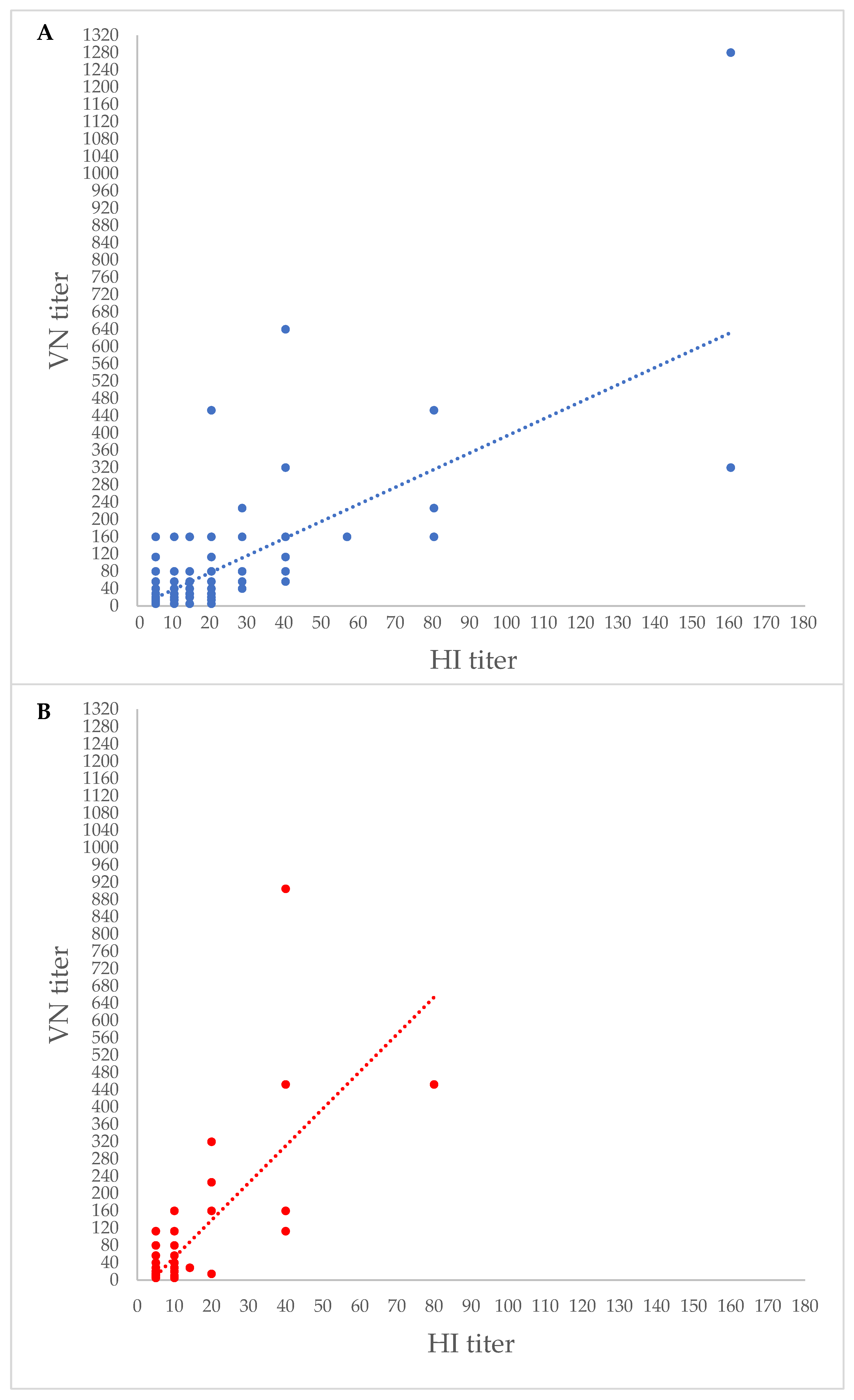
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a novel swine influenza virus from Oklahoma in 2011 which is distantly related to human influenza C viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and Swine/proposal for a new genus in the Orthomyxoviridae family. mBio 2014, 5, e00031-14. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel influenza D virus/Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Foni, E.; Chiapponi, C.; Baioni, L.; Zanni, I.; Merenda, M.; Rosignoli, C.; Kyriakis, C.S.; Luini, M.V.; Mandola, M.L.; Bolzoni, L.; et al. Influenza D in Italy: Towards a better understanding of an emerging viral infection in swine. Sci. Rep. 2017, 7, 11660. [Google Scholar] [CrossRef]
- Ferguson, L.; Luo, K.; Olivier, A.K.; Cunningham, F.L.; Blackmon, S.; Hanson-Dorr, K.; Sun, H.; Baroch, J.; Lutman, M.W.; Quade, B.; et al. Influenza D Virus Infection in Feral Swine Populations, United States. Emerg. Infect. Dis. 2018, 24, 1020–1028. [Google Scholar] [CrossRef]
- Gorin, S.; Fablet, C.; Quéguiner, S.; Barbier, N.; Paboeuf, F.; Hervé, S.; Rose, N.; Simon, G. Assessment of Influenza D Virus in Domestic Pigs and Wild Boars in France: Apparent Limited Spread within Swine Populations Despite Serological Evidence of Breeding Sow Exposure. Viruses. 2019, 12, 25. [Google Scholar] [CrossRef]
- Murakami, S.; Odagiri, T.; Melaku, S.K.; Bazartseren, B.; Ishida, H.; Takenaka-Uema, A.; Muraki, Y.; Sentsui, H.; Horimoto, T. Influenza D Virus Infection in Dromedary Camels, Ethiopia. Emerg. Infect. Dis. 2019, 25, 1224–1226. [Google Scholar] [CrossRef]
- O’Donovan, T.; Donohoe, L.; Ducatez, M.F.; Meyer, G.; Ryan, E. Seroprevalence of influenza D virus in selected sample groups of Irish cattle, sheep and pigs. Ir. Vet. J. 2019, 72, 11. [Google Scholar] [CrossRef]
- Oliva, J.; Eichenbaum, A.; Belin, J.; Gaudino, M.; Guillotin, J.; Alzieu, J.P.; Nicollet, P.; Brugidou, R.; Gueneau, E.; Michel, E.; et al. Serological Evidence of Influenza D Virus Circulation Among Cattle and Small Ruminants in France. Viruses 2019, 11, 516. [Google Scholar] [CrossRef]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza D virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological evidence for the co-circulation of two lineages of influenza D viruses in equine populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Marchi, S.; Manini, I.; Kistner, O.; Li, F.; Piu, P.; Manenti, A.; Biuso, F.; Sreenivasan, C.; Druce, J.; et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses 2019, 12, 30. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Montomoli, E.; Di Bartolo, I.; Ostanello, F.; Chiapponi, C.; Marchi, S. Detection of antibodies against influenza D virus in swine veterinarians in Italy in 2004. J. Med. Virol. 2022, 94, 2855–2859. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza d virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Gaudino, M.; Moreno, A.; Snoeck, C.J.; Zohari, S.; Saegerman, C.; O’Donovan, T.; Ryan, E.; Zanni, I.; Foni, E.; Sausy, A.; et al. Emerging Influenza D virus infection in European livestock as determined in serology studies: Are we underestimating its spread over the continent? Transbound Emerg. Dis. 2021, 68, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, S.; Murakami, S.; Shindo, K.; Horimoto, T.; Sagara, H.; Noda, T.; Kawaoka, Y. Influenza C and D viruses package eight organized ribonucleoprotein complexes. J. Virol. 2018, 92, e02084-17. [Google Scholar] [CrossRef]
- Huang, C.; Yu, J.; Hause, B.M.; Park, J.Y.; Sreenivasan, C.; Uprety, T.; Sheng, Z.; Wang, D.; Li, F. Emergence of new phylogenetic lineage of Influenza D virus with broad antigenicity in California, United States. Emerg. Microbes Infect. 2021, 10, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Yesilbag, K.; Toker, E.B.; Ates, O. Recent strains of influenza D virus create a new genetic cluster for European strains. Microb. Pathog. 2022, 172, 105769. [Google Scholar] [CrossRef]
- Chiapponi, C.; Faccini, S.; Fusaro, A.; Moreno, A.; Prosperi, A.; Merenda, M.; Baioni, L.; Gabbi, V.; Rosignoli, C.; Alborali, G.L.; et al. Detection of a New Genetic Cluster of Influenza D Virus in Italian Cattle. Viruses 2019, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of two distinct genetic and antigenic lineages of proposed influenza D virus in cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef]
- Mazzetto, E.; Bortolami, A.; Fusaro, A.; Mazzacan, E.; Maniero, S.; Vascellari, M.; Beato, M.S.; Schiavon, E.; Chiapponi, C.; Terregino, C.; et al. Replication of Influenza D Viruses of Bovine and Swine Origin in Ovine Respiratory Explants and Their Attachment to the Respiratory Tract of Bovine, Sheep, Goat, Horse, and Swine. Front. Microbiol. 2020, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Kwasnik, M.; Rola, J.; Rozek, W. Influenza D in Domestic and Wild Animals. Viruses 2023, 15, 2433. [Google Scholar] [CrossRef] [PubMed]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef] [PubMed]
- Sanogo, I.N.; Kouakou, C.; Batawui, K.; Djegui, F.; Byarugaba, D.K.; Adjin, R.; Adjabli, K.; Wabwire-Mangen, F.; Erima, B.; Atim, G.; et al. Serological Surveillance of Influenza D Virus in Ruminants and Swine in West and East Africa, 2017–2020. Viruses 2021, 13, 1749. [Google Scholar] [CrossRef]
- Zhai, S.L.; Zhang, H.; Chen, S.N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.H.; Wen, X.H.; Wei, W.K.; Wang, D.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef]
- Tumino, S.; Tolone, M.; Galluzzo, P.; Migliore, S.; Sechi, T.; Bordonaro, S.; Puleio, R.; Carta, A.; Loria, G.R. Alternative Molecular Tools for the Fight against Infectious Diseases of Small Ruminants/Native Sicilian Sheep Breeds and Maedi-Visna Genetic Susceptibility. Animals 2022, 12, 1630. [Google Scholar] [CrossRef]
- Clopper, C.J.; Pearson, E.S. The use of confidence or fiducial limits illustrated in the case of the binomial. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Saegerman, C.; Salem, E.; Ait Lbacha, H.; Alali, S.; Zouagui, Z.; Meyer, G.; Ducatez, M.F. Formal estimation of the seropositivity cut-off of the hemagglutination inhibition assay in field diagnosis of influenza D virus in cattle and estimation of the associated true prevalence in Morocco. Transbound. Emerg. Dis. 2021, 68, 1392–1399. [Google Scholar] [CrossRef]
- Gaudino, M.; Chiapponi, C.; Moreno, A.; Zohari, S.; O’Donovan, T.; Quinless, E.; Sausy, A.; Oliva, J.; Salem, E.; Fusade-Boyer, M.; et al. Evolutionary and temporal dynamics of emerging influenza D virus in Europe (2009-22). Virus Evol. 2022, 8, veac081. [Google Scholar] [CrossRef] [PubMed]
- Hirst, G.K. The agglutination of red cells by allantoic fluid of chick embryos infected with influenza virus. Science 1941, 94, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.M.; Gerlach, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J. Clin. Virol. 2019, 119, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Reber, A.; Katz, J. Immunological assessment of influenza vaccines and immune correlates of protection. Expert Rev. Vaccines 2013, 12, 519–536. [Google Scholar] [CrossRef]
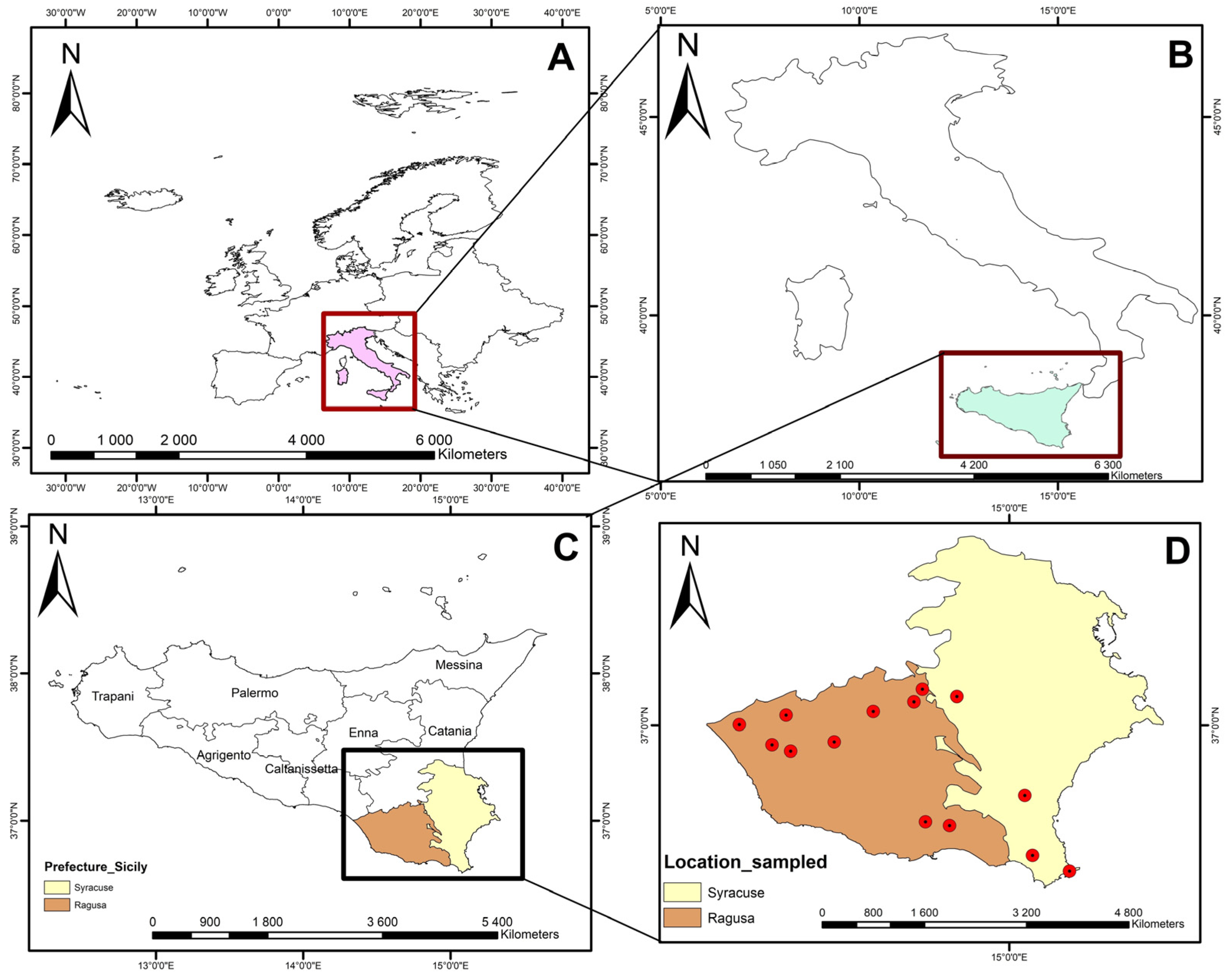
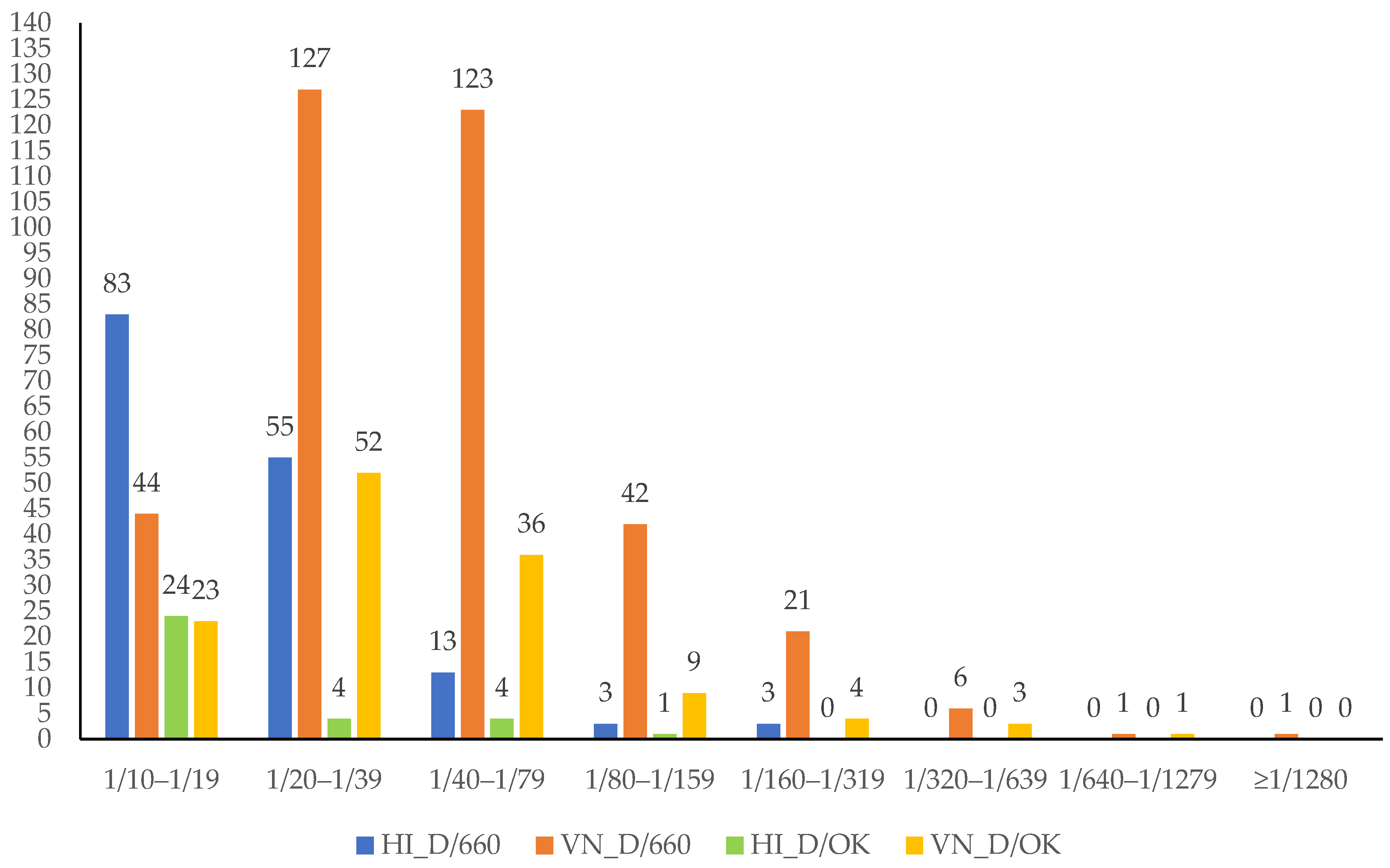
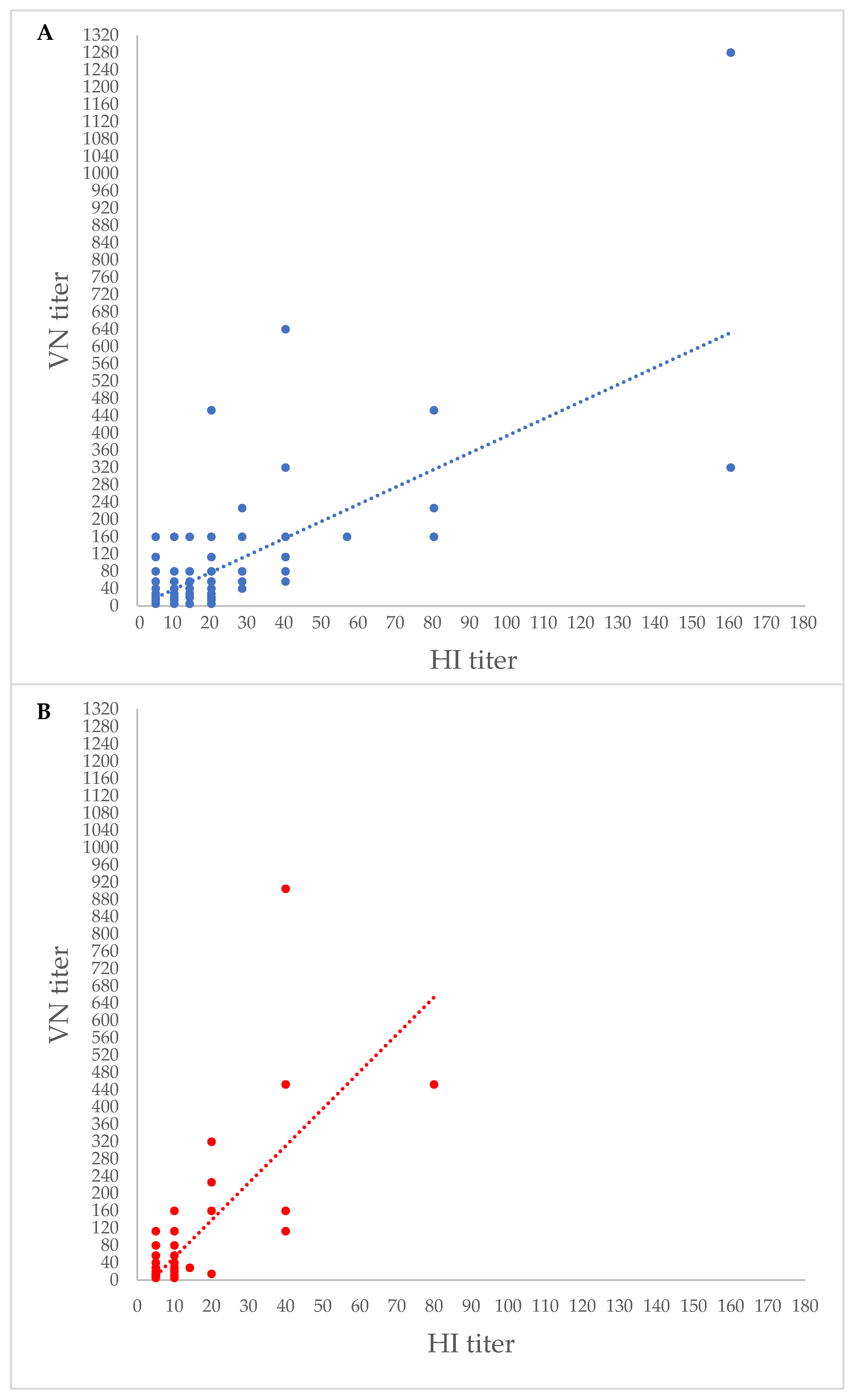

{kind=link}
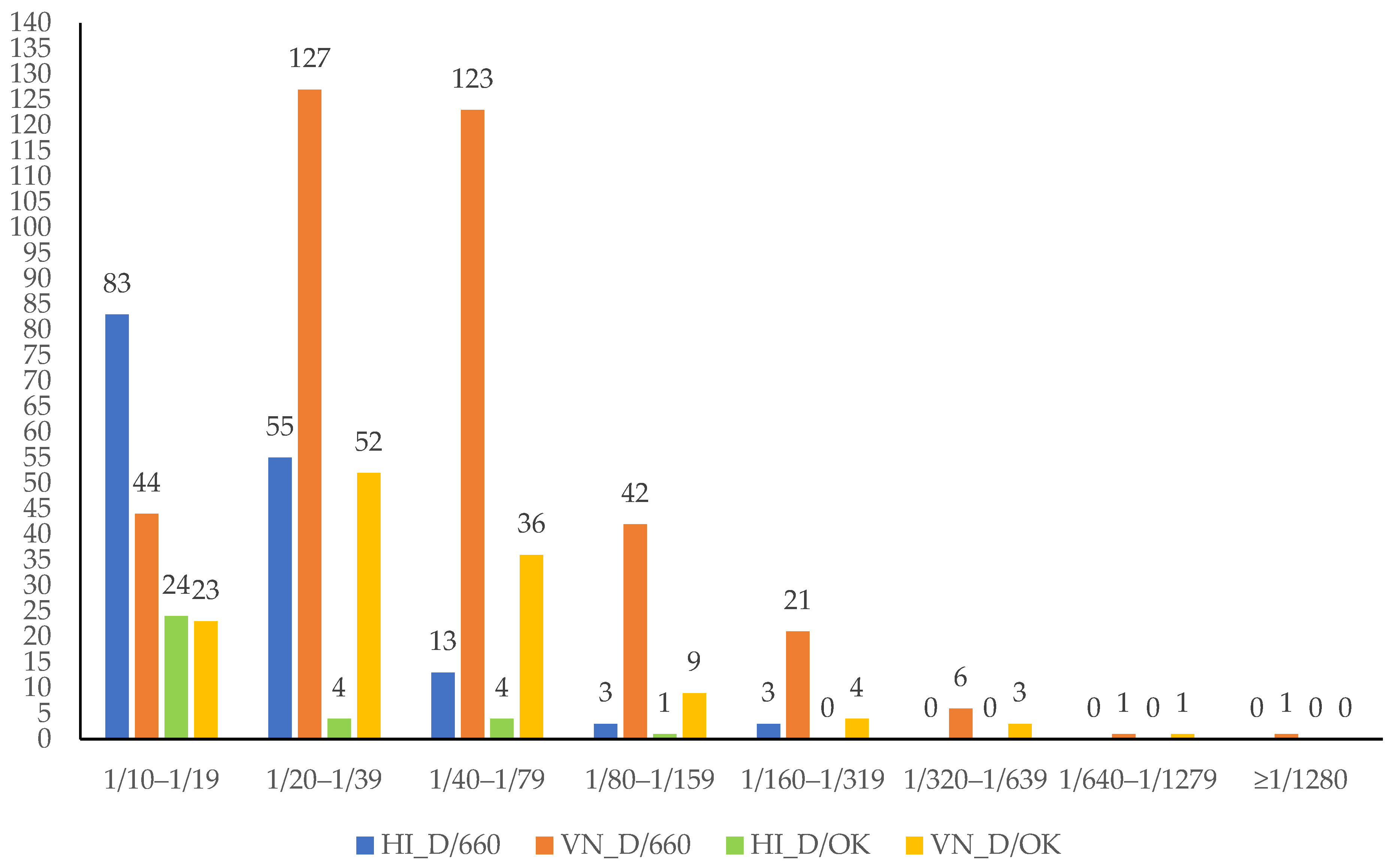{kind=link}
{kind=link}
| Prefecture | Sheep Herd | Location and Zip Code | Samples Collected |
|---|---|---|---|
| RG | A | Acate 97011, | 50 |
| D | Comiso, 97013, | 65 | |
| E | Baucino, Vittoria, 97019 | 64 | |
| F | Niscescia, Vittoria, 97019 | 43 | |
| I | Linfante, Vittoria, 97019 | 70 | |
| K | Ispica, 97014 | 50 | |
| L | Giarratana, 97010 | 35 | |
| M | Chiaramonte Gulfi, 97012 | 20 | |
| O | Modica, 97015 | 10 | |
| Total RG | #9 | 407 | |
| SR | B | Buscemi, 96010 | 57 |
| C | Vallefame, 96010 | 7 | |
| G | Noto, 96017 | 57 | |
| H | Pachino, 96018 | 12 | |
| J | Palazzolo Acreide, 96010 | 45 | |
| N | Portopalo di Capo Passero, 96010 | 15 | |
| Total SR | #6 | 193 | |
| Total | #15 | 600 |
| Prefecture | Sheep Herd | Samples Collected | HI_D/660 Titre | VN_ D/660 Titre | HI_ D/OK Titre | VN_ D/OK Titre | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| ≥1/10 | <1/10 | ≥1/10 | <1/10 | ≥1/10 | <1/10 | ≥1/10 | <1/10 | |||
| RG | A | 50 | 12 | 38 | 32 | 18 | 4 | 46 | 7 | 43 |
| D | 65 | 8 | 57 | 34 | 31 | 1 | 64 | 4 | 61 | |
| E | 64 | 22 | 42 | 37 | 27 | 5 | 59 | 9 | 55 | |
| F | 43 | 11 | 32 | 16 | 27 | 2 | 41 | 4 | 39 | |
| I | 70 | 29 | 41 | 39 | 31 | 1 | 69 | 15 | 55 | |
| K | 50 | 21 | 29 | 41 | 9 | 1 | 49 | 16 | 34 | |
| L | 35 | 7 | 28 | 23 | 12 | 2 | 33 | 11 | 24 | |
| M | 20 | 2 | 18 | 13 | 7 | 0 | 20 | 5 | 15 | |
| O | 10 | 7 | 3 | 8 | 2 | 4 | 6 | 5 | 5 | |
| Total RG | #9 | 407 | 119 | 288 | 243 | 164 | 20 | 387 | 76 | 331 |
| SR | B | 57 | 3 | 54 | 43 | 14 | 2 | 55 | 9 | 48 |
| C | 7 | 0 | 7 | 6 | 1 | 0 | 7 | 0 | 7 | |
| G | 57 | 10 | 47 | 30 | 27 | 5 | 52 | 11 | 46 | |
| H | 12 | 0 | 12 | 8 | 4 | 2 | 10 | 4 | 8 | |
| J | 45 | 20 | 25 | 26 | 19 | 4 | 41 | 23 | 22 | |
| N | 15 | 5 | 10 | 9 | 6 | 0 | 15 | 5 | 10 | |
| Total SR | #6 | 193 | 38 | 155 | 122 | 71 | 13 | 180 | 52 | 141 |
| Total | #15 | 600 | 157 | 443 | 365 | 235 | 33 | 567 | 128 | 472 |
| Prefecture | Sheep Herd | HI Assay | VN Assay | D/660 | D/OK | HI and VN Assays | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D/660 | D/OK | D/660 | D/OK | HI Assay | VN Assay | HI Assay | VN Assay | D/660 | D/OK | ||
| RG | A | 4 | 6 | 9 | 3 | 2 | |||||
| D | 0 | 4 | 8 | 1 | 0 | ||||||
| E | 4 | 8 | 21 | 5 | 3 | ||||||
| F | 2 | 4 | 11 | 2 | 2 | ||||||
| I | 0 | 13 | 27 | 1 | 0 | ||||||
| K | 1 | 16 | 19 | 1 | 1 | ||||||
| L | 0 | 9 | 7 | 2 | 0 | ||||||
| M | 0 | 4 | 2 | 0 | 0 | ||||||
| O | 4 | 5 | 7 | 4 | 4 | ||||||
| Total RG | #9 | 15 | 69 | 111 | 19 | 12 | |||||
| SR | B | 0 | 9 | 3 | 2 | 0 | |||||
| C | 0 | 0 | 0 | 0 | 0 | ||||||
| G | 3 | 9 | 10 | 5 | 3 | ||||||
| H | 0 | 3 | 0 | 2 | 0 | ||||||
| J | 4 | 20 | 18 | 4 | 4 | ||||||
| N | 0 | 5 | 4 | 0 | 0 | ||||||
| Total SR | #6 | 7 | 46 | 35 | 13 | 7 | |||||
| Total | #15 | 22 | 115 | 146 | 32 | 19 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanave, G.; Camero, M.; Coppola, C.; Marchi, S.; Cascone, G.; Salina, F.; Coltraro, M.; Odigie, A.E.; Montomoli, E.; Chiapponi, C.; et al. Serological Evidence for Circulation of Influenza D Virus in the Ovine Population in Italy. Pathogens 2024, 13, 162. https://doi.org/10.3390/pathogens13020162
Lanave G, Camero M, Coppola C, Marchi S, Cascone G, Salina F, Coltraro M, Odigie AE, Montomoli E, Chiapponi C, et al. Serological Evidence for Circulation of Influenza D Virus in the Ovine Population in Italy. Pathogens. 2024; 13(2):162. https://doi.org/10.3390/pathogens13020162
Chicago/Turabian StyleLanave, Gianvito, Michele Camero, Chiara Coppola, Serena Marchi, Giuseppe Cascone, Felice Salina, Miriana Coltraro, Amienwanlen E. Odigie, Emanuele Montomoli, Chiara Chiapponi, and et al. 2024. "Serological Evidence for Circulation of Influenza D Virus in the Ovine Population in Italy" Pathogens 13, no. 2: 162. https://doi.org/10.3390/pathogens13020162
APA StyleLanave, G., Camero, M., Coppola, C., Marchi, S., Cascone, G., Salina, F., Coltraro, M., Odigie, A. E., Montomoli, E., Chiapponi, C., Cicirelli, V., Martella, V., & Trombetta, C. M. (2024). Serological Evidence for Circulation of Influenza D Virus in the Ovine Population in Italy. Pathogens, 13(2), 162. https://doi.org/10.3390/pathogens13020162