Abstract
Toxoplasmosis caused by Toxoplasma gondii is an important zoonosis of human and animal health significance. Current chemical therapeutics have side effects, and no commercially available vaccine is licensed for the prevention of toxoplasmosis in humans and most animals. Developing a safe and effective vaccine with long-term protection against T. gondii infection is necessary to control toxoplasmosis. HAD2a is a key member of the haloacid dehalogenase (HAD) phosphatase family, which is essential for T. gondii daughter budding. However, the role of HAD2a in T. gondii virulence remains unknown. In this study, we successfully constructed the had2a gene knockout strain in the T. gondii-type I RH strain (RHΔhad2a) and determined its role in virulence and vaccination. These results demonstrate that HAD2a played an important role in parasite daughter budding and in vitro replication. Knockout of the had2a gene attenuated the virulence of the T. gondii-type I RH strain. Vaccination with RHΔhad2a tachyzoites induced a Th1-biased immune response, provided partial protection against acute T. gondii infection in mice by highly virulent tachyzoites of RH and PYS (ToxoDB#9, Chinese I) strains, and conferred strong protection against challenge infection by cysts and oocysts of the less virulent type II Pru strain. These results demonstrate that T. gondii had2a is important for its in vitro proliferation and virulence in mice and that RHΔhad2a may be used as a candidate strain to generate a multiple gene knockout live-attenuated strain or be collaboratively applied with other live-attenuated strains to confer more effective protection against T. gondii infection.
1. Introduction
Toxoplasma gondii is an obligate intracellular protozoan that infects almost all warm-blooded animals and approximately 25 to 30% of the world’s human population [1,2,3]. Foodborne transmission is the primary route of T. gondii infection in humans [4,5]. The consumption of undercooked or raw meat products contaminated with cysts or ingestion of vegetables or water contaminated with oocysts may result in T. gondii infection [1,2,3,4,5]. Infection with T. gondii severely threatens the health of pregnant women and immunocompromised patients (such as HIV and organ transplant patients) [5,6,7]. Primary T. gondii infection during pregnancy may cause premature delivery, stillbirth, congenital abnormalities, developmental disabilities, growth retardation and meningitis in fetuses [5,6,8,9]. Although significant advances have been achieved in studies of T. gondii biology and epidemiology, current toxoplasmosis treatment, mainly pharmacotherapy, can only manage acute and reactivated infections without the clearing of T. gondii tissue cysts [2,5,8].
Preventing T. gondii infection or clearing latent infection may be achieved by vaccination. Toxovax, a live-attenuated vaccine derived from T. gondii S48 tachyzoites and licensed in a few countries, has been used in sheep against toxoplasmosis, especially preventing miscarriage in pregnant sheep [10]. However, this licensed vaccine has some application defects, such as a relatively short shelf life, uncomplete block to congenital infection and a possible reversion to a virulent phenotype [8]. Significant toxoplasmosis vaccine development has been explored in the last few decades, such as live-attenuated strains, DNA vaccines, epitope vaccines, exosome-based, carbohydrate-based and nanoparticle-based vaccination [8,11,12]. With the application of the CRISPR-Cas9 gene-editing tool in T. gondii research, several knockout strains of important T. gondii genes have been tested in vaccination studies in mice [8,13,14]. These virulence-attenuated vaccine strains include RHΔtkl1 [13], RHΔgra17Δnpt1 [15], RHΔompdcΔuprt [16], ME49Δcdpk3 [17] and PruΔcdpk2 [18], which have provided efficient protection against acute, chronic, and congenital infection.
HAD2a (TGGT1_289910), which possesses a haloacid dehalogenase (HAD) phosphatase domain, is essential for T. gondii daughter budding [19]. Conditional knockout of the had2a gene leads to incomplete cytokinesis, conjoined daughters, and disrupted proliferation [19]. However, the role of HAD2a in T. gondii virulence remains unknown. The in vitro replication defect caused by the conditional knockout of the had2a gene indicates that the had2a gene mutant strain might have replication defect and attenuated virulence, suggesting a potential role as a live-attenuated vaccine. Here, we successfully constructed the had2a gene knockout strain RHΔhad2a and characterized the virulence of RHΔhad2a in mice. The immunization results revealed that RHΔhad2a vaccination significantly induced specific anti-T. gondii antibodies and conferred partial protection against T. gondii acute and chronic infection. Our findings suggest that HAD2a is important for T. gondii virulence and RHΔhad2a can be used as a parental strain to generate an attenuated multiple gene knockout vaccine strain.
2. Materials and Methods
2.1. Animals and Ethics Statement
The female Kunming mice (6–8 weeks old) used in the experiment were purchased from the Laboratory Animal Center of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Lanzhou, China). To reduce the impact of stress response on the experiment, mice were placed in a temperature-controlled room with a 12 h light/dark cycle and provided with sufficient sterile water and food for free intake. The study was approved by Animal Ethics Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Approval no. 2020-022). Every effort was made to reduce any suffering of the animals.
2.2. Parasite Culture
Toxoplasma gondii tachyzoites of the parental RH∆ku80 (referred as RH) stain, PYS (ToxoDB#9, Chinese I) strain and gene mutant strains were maintained in confluent monolayers of human foreskin fibroblasts (HFFs, ATCC SCRC-1041TM) maintained in DMEM supplemented with 2% fetal bovine serum (FBS, Gibco, Auckland, New Zealand), 10 mM HEPES (pH 7.2, Solarbio, Beijing, China), 100 U/mL of penicillin (Solarbio, China) and 100 μg/mL of streptomycin (Solarbio, China) as described previously [20,21]. For further study, the tachyzoites were isolated and purified by 27-gauge needles (BD Medical, Franklin Lake, WI, USA) and Millipore filters (Merck-Millipore, Darmstadt, Germany) with a pore size of 5 µm. Tachyzoites were counted by hemocytometer measurement and diluted to required number of tachyzoites in 200 μL PBS or DMEM [22].
2.3. Construction of had2a Gene Knockout Strain
The CRISPR-Cas9 mediated homologous gene recombination was used to delete the had2a gene in the wild-type RH strain [23]. The CRISPR plasmid pSAG1::CAS9-U6::Sg HAD2a and homologous dihydrofolate reductase (DHFR) drug-selective plasmid were constructed. To generate the had2a gene knockout CRISPR plasmid pSAG1::CAS9-U6::Sg HAD2a, the SgRNA of had2a gene was used to replace the SgRNA of the uracil phosphoribosyl transferase (uprt) gene in the pSAG1::CAS9-U6::SgUPRT template plasmid [24]. The 5′ and 3′ homologous arms of the had2a gene (1000~1300 bp) were fused to the DHFR fragment and the pUC19 backbone fragment to construct the homologous DHFR-selective plasmid [25]. The sequence-validated plasmid was used as a template to amplify the homologous DHFR fragment. To construct the had2a gene knockout strain (RHΔhad2a), the purified homologous drug-selective cassette and sequencing-validated CRISPR plasmid were co-transfected into the wild-type RH tachyzoites. After pyrimethamine selection, single clones of RHΔhad2a obtained by modified limiting dilution were confirmed by designed PCRs [20]. The primers used are listed in Table 1.

Table 1.
Primers used in this study.
2.4. Immunofluorescence Assay (IFA)
To visualize the effect of had2a gene deletion on parasite daughter budding, the purified RH and RHΔhad2a tachyzoites were used to infect the HFF monolayers. After fixed with 4% paraformaldehyde for 20 min, the samples were permeabilized with 0.2% Triton X-100 for 15 min and incubated with polyclonal rabbit anti-IMC1 (1:500, available in our lab) [20] and goat anti-rabbit IgG (H+L) conjugated with Alexa Fluor 488 (1:500, Thermo Fisher Scientific, Waltham, MA, USA) for 1 h at 37 °C, each of which was followed by five washes with PBS [26]. A Leica confocal microscope system (TCS SP8, Leica, Wetzlar, Germany) was used to image the samples.
2.5. Plaque Assay
Confluent HFF monolayers grown in 12-well plates (Thermo Fisher Scientific, USA) were infected with 300 freshly egressed tachyzoites of wild-type RH or mutant RHΔhad2a strains for 7 days. After the culture medium was removed, samples were washed with PBS, fixed with 4% paraformaldehyde for 20 min and stained with 0.2% crystal violet for 20 min, which was followed by two washes with PBS [25]. Three independent replicates were performed. The plaque size and number were analyzed with the ImageJ software version 1.53a.
2.6. Intracellular Replication Assay
Freshly egressed tachyzoites (1 × 105) of RHΔhad2a or RH strain were used to infect the confluent HFF monolayers cultured in 12-well plates for 1 h prior to washing the uninvaded tachyzoites. The number of tachyzoites in at least 100 parasitophorous vacuoles of samples was counted using IFA after 24 h incubation, in which mouse anti-SAG1 (1:500, Thermo Fisher Scientific, USA) and goat-anti mouse conjugated with Alexa Fluor 488 were used to visualize intracellular parasites [27].
2.7. Virulence Assessment in Mice
Female Kunming mice were randomly divided into six groups (10 mice/group). Different doses of RH (1 × 102) or RHΔhad2a (1 × 102, 1 × 103, 1 × 104, 1 × 105, 1 × 106) tachyzoites diluted in 200 μL PBS were used to infect mice intraperitoneally. During 30 days of infection, the survival and clinical signs of all infected mice group were observed twice daily [28]. Mice reaching a humane endpoint were euthanized immediately.
2.8. Antibodies Assessment in Immunized Mice
Kunming mice were intraperitoneally immunized with 5 × 104 RHΔhad2a tachyzoites, which was diluted in 200 μL PBS and quantified by plaque assay to ensure its number and viability used in vaccination. Naive mice were intraperitoneally injected with 200 μL PBS. Serum samples of the naive and immunized mice (six mice per group) were collected at 30 days post-immunization. To detect whether anti-T. gondii antibodies were induced, subclass antibodies IgG1 and IgG2a along with total IgG were determined using ELISA [13,15].
2.9. Protection of RHΔhad2a Immunization against T. gondii Infection
Thirty days post-vaccination with 5 × 104 RHΔhad2a tachyzoites, 1 × 103 T. gondii tachyzoites of the wild-type RH strain or PYS strain were used to intraperitoneally infect each of the immunized mice and naive mice (ten mice per group). For chronic infection, the naive and immunized mice (ten mice per group) were orally administrated with 20 Pru cysts or 50 Pru oocysts. The Pru cysts were collected and purified from the brain tissues of Kunming mice infected with cysts according to previous descriptions [13,29], whereas the Pru oocysts were collected and purified from feces of kittens orally infected with cysts [13,30,31]. The survival time and toxoplasmosis clinical signs of all infected mice group were observed twice daily. Mice were euthanized immediately when reaching the humane endpoint. Brain cysts of all chronically survived mice were counted as previously [20,25]. For the detection of brain cyst burden, the brains of the mice that survived for 30 days post-chronic challenge were collected, homogenized, and used to count brain cyst number, as described previously [15].
2.10. Statistical Analysis
All data obtained from three independent replicates were represented as means ± standard deviations (SD). Group difference was analyzed by a two-tailed, unpaired Student’s t test using GraphPad Prism version 9.0. Difference was considered significant when the p value < 0.05.
3. Results
3.1. Successful Construction of RHΔhad2a
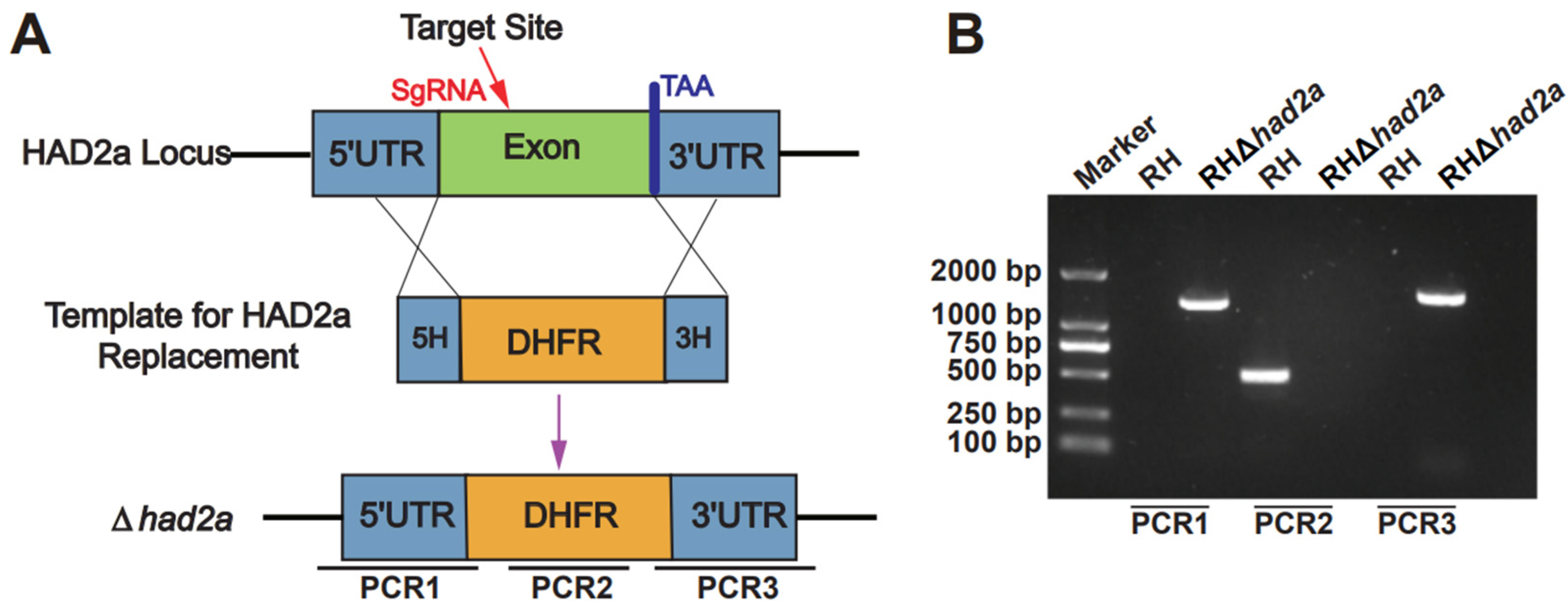
To investigate the role of HAD2a in T. gondii virulence, the had2a gene was knocked out in the T. gondii-type I RH strain by replacing the had2a gene with a homologous DHFR fragment using CRISPR-Cas9 (Figure 1A). The successful construction of gene knockout strain RHΔhad2a was confirmed by PCRs. In PCR1 and PCR3, recombinant 5′ and 3′ homologous fragments (~1300 bp) were amplified in the RHΔhad2a strain but not in the RH strain (Figure 1B), validating the successful insertion of 5′ and 3′ homologous fragments in RHΔhad2a. The small had2a coding gene fragment was amplified in PCR2 of the RH strain but not in PCR2 of the RHΔhad2a strain (Figure 1B), confirming the disruption of the had2a gen in RHΔhad2a. These results demonstrated that had2a gene knockout strain RHΔhad2a was successfully generated by CRISPR-Cas9 mediated homologous recombination.
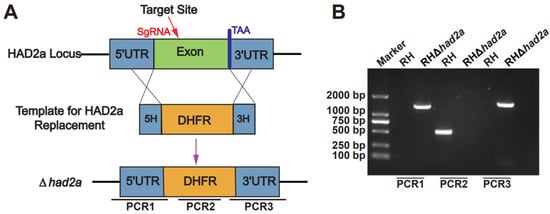
Figure 1.
Construction and validation of RHΔhad2a. (A) Schematic illustration of had2a gene deletion by CRISPR-Cas9 mediated homologous gene replacement. (B) PCRs of RHΔhad2a. Recombinant homologous fragments of 5′ and 3′ in RHΔhad2a were detected by PCR1 and PCR3. Deletion of had2a gene was detected by PCR2.
3.2. Knockout of had2a Gene Severely Attenuated T. gondii Virulence in Mice
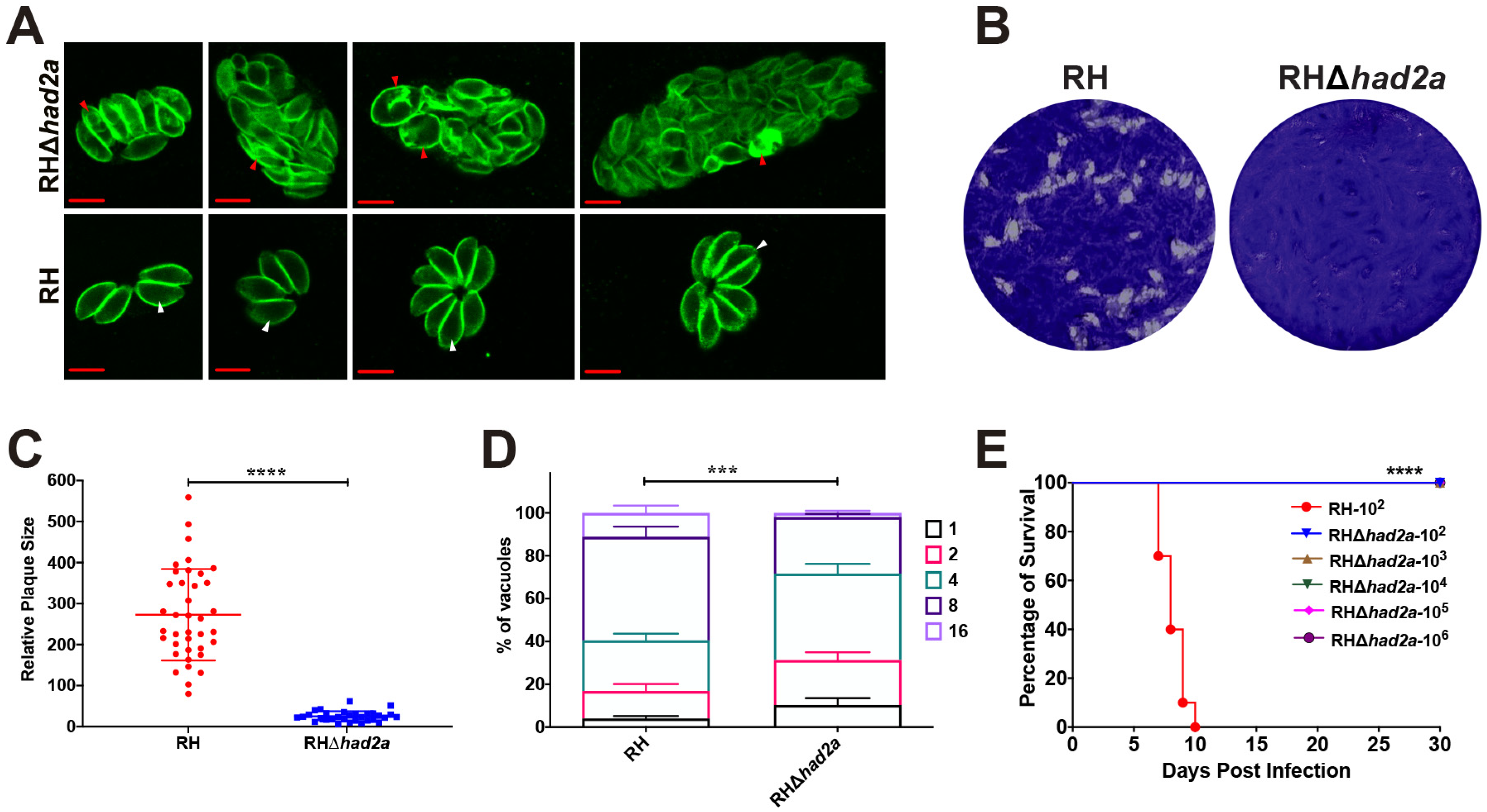
To assess the effect of had2a disruption on the replication and virulence of T. gondii, we performed IFA, in vitro plaque and in vivo virulence assays using RHΔhad2a and wild-type RH strains. In IFA, a division defect was observed in RHΔhad2a (Figure 2A). Some intravacuolar tachyzoites of RHΔhad2a were enlarged without daughter budding or the separation of dividing tachyzoites, lacking the rosette appearance of wild-type RH parasites. These results confirmed the role of HAD2a in parasite daughter budding as previously described [19]. In an in vitro plaque assay, the same dose of RHΔhad2a or RH tachyzoites were used to infect the HFF monolayer. After 7 days of incubation, the HFF samples were stained with crystal violet. The results showed that knockout of the had2a gene caused a significant defect in the plaque-forming ability (Figure 2B,C). The in vitro parasite replication rates were also monitored 24 h post infection. The deletion of had2a led to a significant decrease in the replication of T. gondii type I strain (Figure 2D). The significant defect of RHΔhad2a in plaque-forming ability and replication efficacy demonstrates that HAD2a plays an important role in the in vitro proliferation of T. gondii.
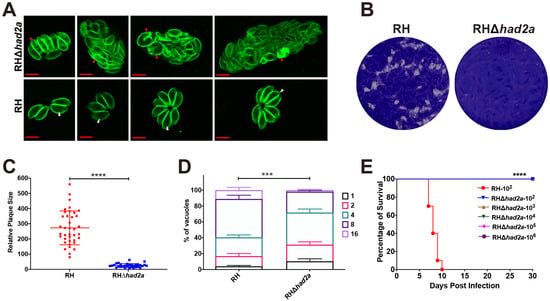
Figure 2.
Knockout of had2a affected T. gondii daughter budding, replication, and virulence. (A) Division defect of intracellular RHΔhad2a dividing parasites. Wild-type RH parasites show normal daughter parasite budding (white arrows), while some RHΔhad2a parasites show the aberrant morphology of intercellular parasites and improper segregation of daughter cells (red arrows). (B–D) Replication defect of RHΔhad2a parasites in vitro. Plaques comparison of wild-type RH and RHΔhad2a parasites (B). Relative size and number of plaques were significantly affected by had2a gene deletion (**** p < 0.0001) (C). (D) Intracellular replication of RH and RHΔhad2a parasites at 24 h post infection of HFFs determined in three independent experiments. RHΔhad2a parasites had a slower replication compared to wild-type RH strain (*** p < 0.001). (E) Attenuated virulence of RHΔhad2a parasites. All mice infected with different doses of RHΔhad2a tachyzoites survived, while all mice infected with wild-type RH tachyzoites were sacrificed (**** p < 0.0001).
To test whether HAD2a is important for T. gondii virulence, different doses of RHΔhad2a or RH tachyzoites were used to infect Kunming mice. All mice infected with different doses (102, 103, 104, 105, 106) of RHΔhad2a tachyzoites survived without any clinical symptoms of toxoplasmosis, whereas mice infected with 102 tachyzoites of the wild-type RH strain were sacrificed within 10 days post-infection (Figure 2E). These results suggest that HAD2a is important for T. gondii virulence and that RHΔhad2a may be a potential candidate for developing anti-T. gondii live-attenuated vaccine.
3.3. RHΔhad2a Vaccination Induces a Th1-Dominated Immune Response
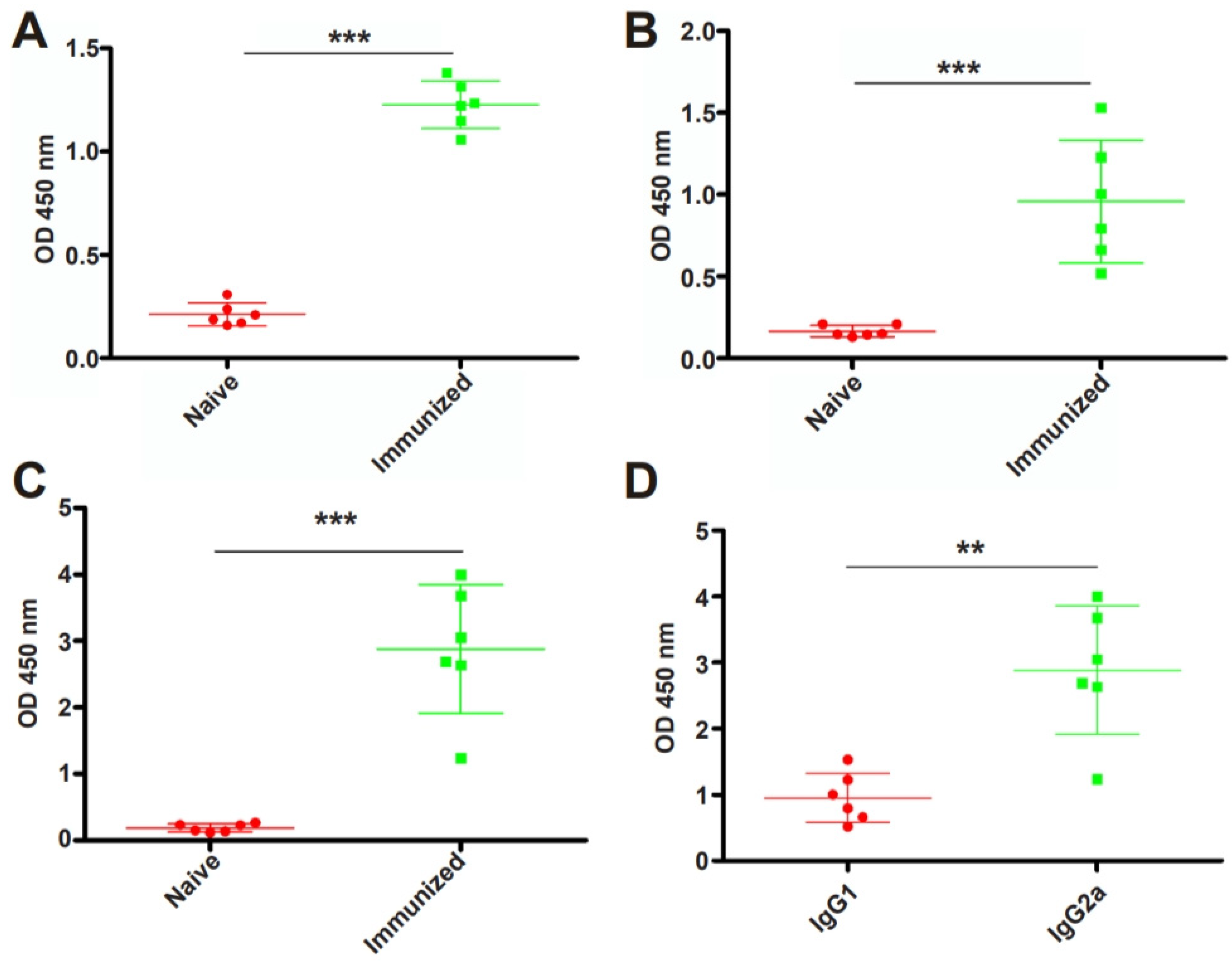
To examine whether RHΔhad2a vaccination induces the production of anti-T. gondii-specific IgG, Kunming mice were immunized with 5 × 104 RHΔhad2a tachyzoites (Figure 3A) [25]. The sera of RHΔhad2a-vaccinated and unvaccinated mice were isolated at 30 days post-infection. The total IgG, subclass antibodies IgG1 and IgG2a were determined by ELISA (Figure 3B). These results showed that the sera obtained from RHΔhad2a-vaccinated mice had significantly higher levels of IgG, IgG1 and IgG2a compared with the sera of non-vaccinated mice (Figure 4A–C). The IgG2a level was higher than the IgG1 level in the sera of RHΔhad2a-vaccinated mice (Figure 4D). These results illuminated that RHΔhad2a vaccination induced a Th1-dominated immune response and that RHΔhad2a has the potential to be a live-attenuated vaccine candidate.
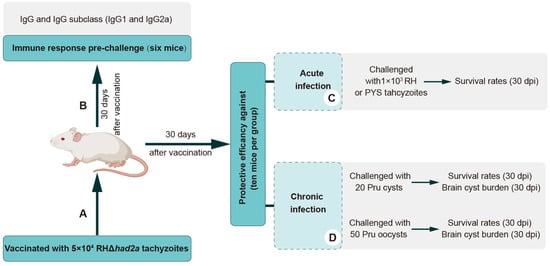
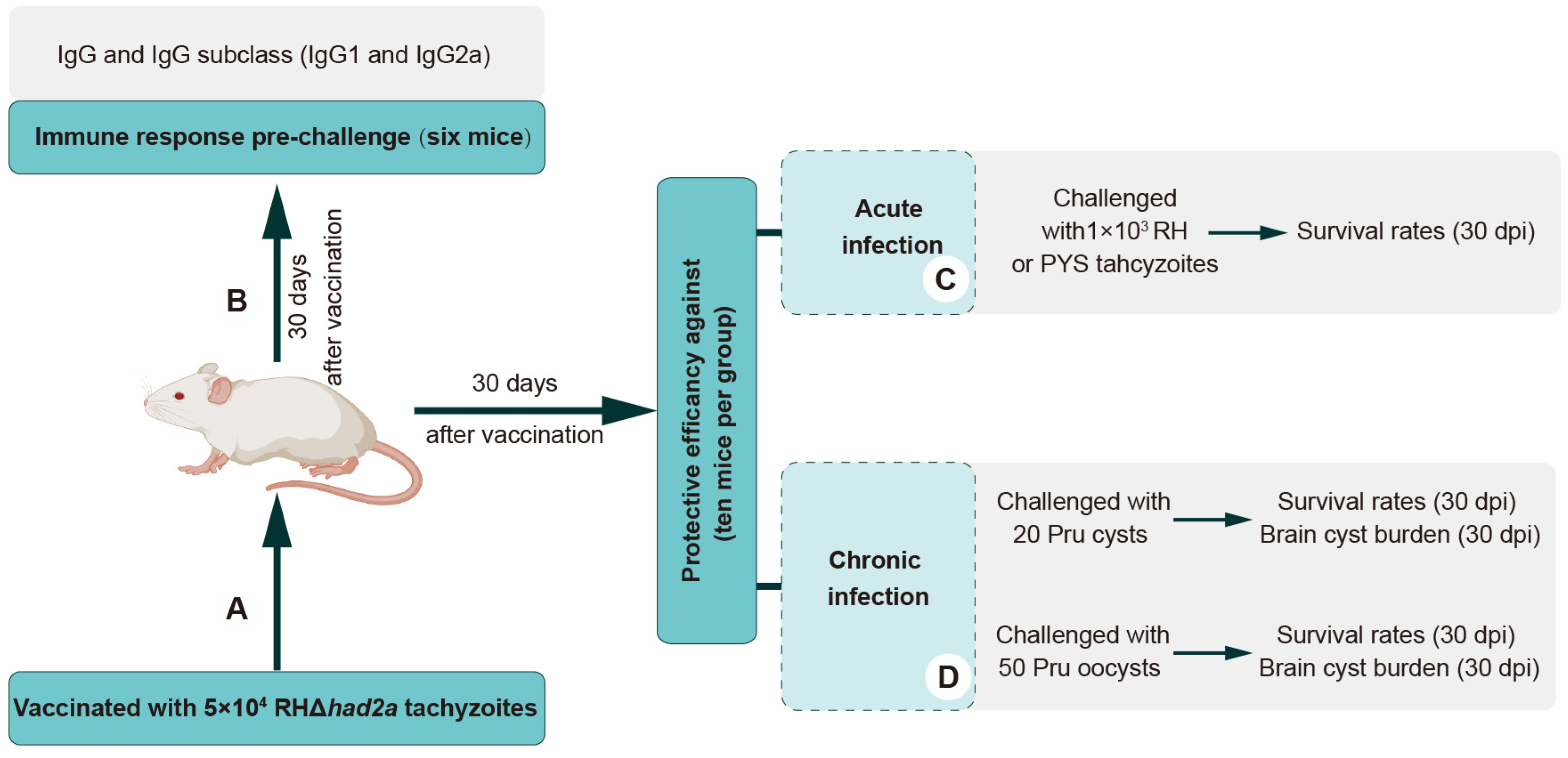
Figure 3.
Schematic illustration of the study design of RHΔhad2a vaccination. Experimental overview of Kunming mice vaccination with 5 × 104 RHΔhad2a tachyzoites (A), and determination of anti-T. gondii antibodies of vaccinated mice prior to challenge (B) at acute (C) and chronic stages of infection (D).
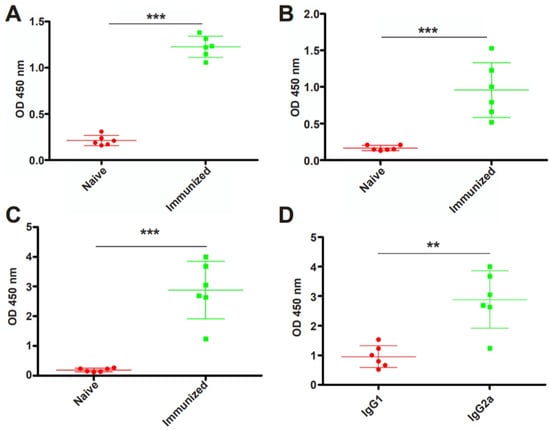
Figure 4.
Immunization with RHΔhad2a induced a significant increased level of anti-T. gondii-specific antibodies. Mice were vaccinated with 5 × 104 RHΔhad2a tachyzoites for 30 days. Total IgG (A), IgG1 (B) and IgG2a (C) in sera of the naive and immunized mice (*** p < 0.001). Comparison of IgG1 and IgG2a levels in immunized mice (** p < 0.01) (D).
3.4. Vaccination of RHΔhad2a Provides Partial Protection in Mice against Acute Infection of Virulent Strains
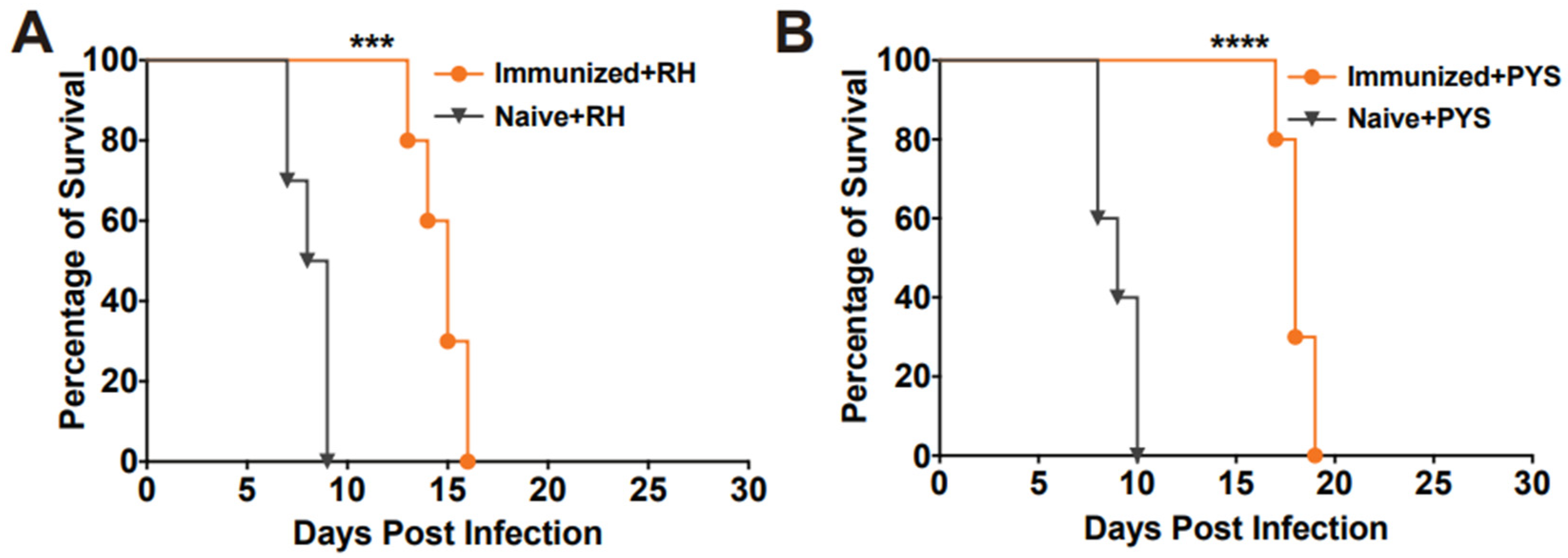
To assess the protective efficacy of RHΔhad2a vaccination against tachyzoite infection, Kunming mice were immunized with 5 × 104 RHΔhad2a tachyzoites (Figure 3A) [25]. After 30 days post-immunization, the vaccinated mice and unvaccinated mice were infected by 103 wild-type RH or PYS tachyzoites (Figure 3C). The results showed that all unvaccinated mice were sacrificed within 10 days when challenged with RH or PYS tachyzoites (Figure 5A,B). Although the vaccinated mice were also sacrificed, the survival time was significantly prolonged in the vaccinated group (sacrificed within 18 days) (Figure 5A,B). These results indicate that immunization of the live-attenuated RHΔhad2a confers partial protection against acute T. gondii infection by virulent strains.
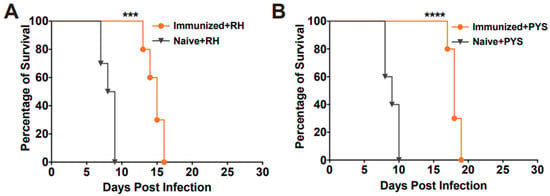
Figure 5.
RHΔhad2a immunization significantly prolonged the survival time of mice with acute T. gondii infection. The naive and immunized mice were challenged with 1 × 103 tachyzoites of type I RH strain (A) or PYS (ToxoDB#9, Chinese I) strain (B) 30 days after immunization (*** p < 0.001, **** p < 0.0001). The survival time of the vaccinated mice was significantly longer compared to that of the naive mice.
3.5. RHΔhad2a Vaccination Confers Partial Protection against Chronic Infection
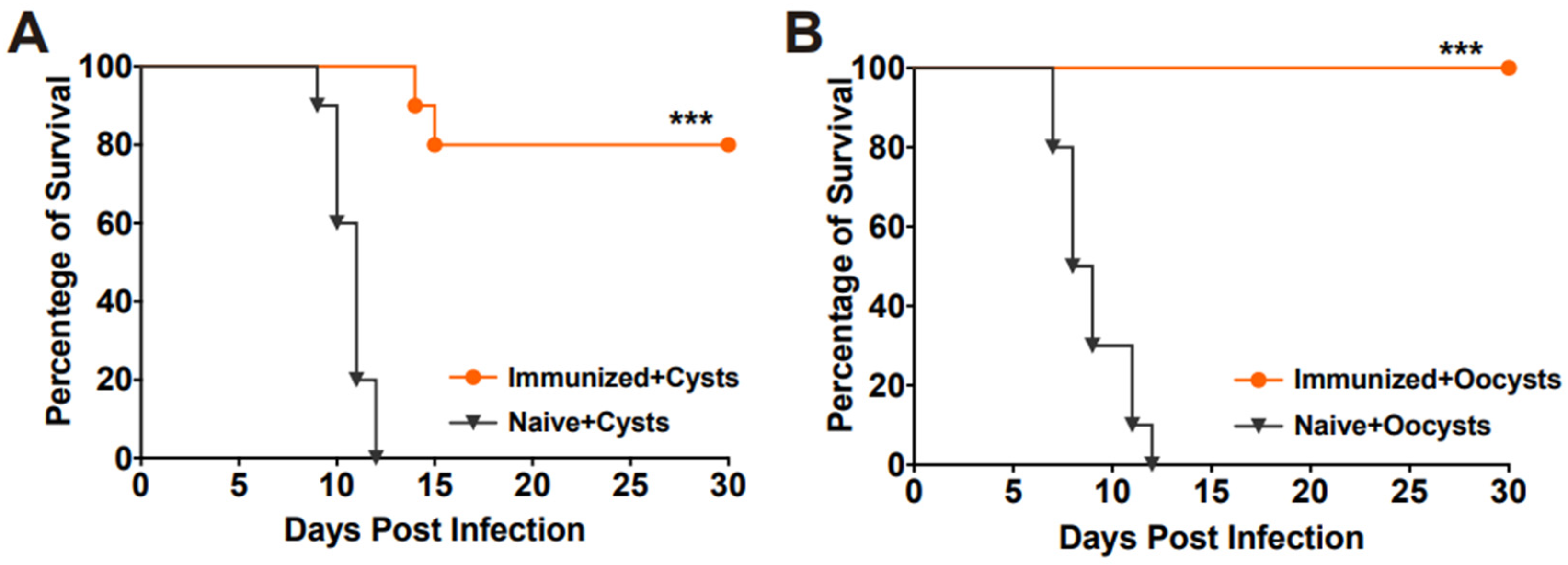
To evaluate the protective efficacy of RHΔhad2a vaccination against chronic infection, mice were orally challenged with 20 Pru cysts or 50 Pru oocysts after 30 days post-vaccination (Figure 3D). The results showed that all unimmunized mice were sacrificed during the chronic stage of challenge infection by cysts or oocysts of the Pru strain (Figure 6A,B), whereas 80% of the vaccinated mice in the cysts-challenged group and 100% of the vaccinated mice in the oocysts-challenged group survived (Figure 6A,B). At 30 days post-challenge, the brain cyst numbers of the surviving mice were determined. In immunized groups, two out of ten mice in the Pru cysts-challenged group were sacrificed, and brain cysts were detected in only three out of the eight surviving mice (median = 45 ± 32). In the oocysts-challenged group, only three out of ten surviving mice had brain cysts (median = 20 ± 22). The numbers of brain cysts in the surviving vaccinated mice were significantly lower than that previously reported [15,25]. These results suggest that RHΔhad2a vaccination induced partial protection against chronic T. gondii infection.
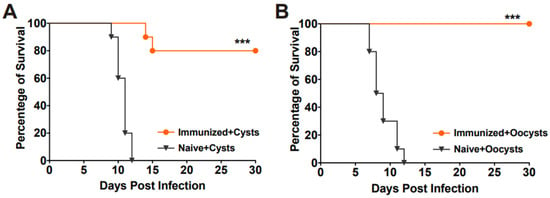
Figure 6.
Vaccination with RHΔhad2a protected mice from chronic T. gondii infection. Survival time of the naive and immunized mice orally infected with 20 cysts (A) or 50 oocysts (B) of type II T. gondii Pru strain, 30 days after immunization with RHΔhad2a tachyzoites (*** p < 0.001). Survival percentages of the vaccinated mice were significantly higher than that of the naive mice.
4. Discussion
Toxoplasmosis caused by T. gondii infection is a significant public health threat to humans and animals [5,32]. Due to its elaborate immune evasion system and complex life cycle of T. gondii, it is difficult to prevent and treat toxoplasmosis [5,32]. Although the combined use of pyrimethamine and sulfadiazine is the primary therapeutic regimen, failure of treatment happened quite often [8]. Side effects of current therapeutics may occur in treated patients [33]. Vaccination of humans and animals with effective vaccines would reduce chemical therapeutic consequences and reliance, and it offers a better alternative for the efficient and long-term control of zoonotic toxoplasmosis [8,34].
HAD2a is essential for T. gondii daughter budding, which is an important part of the parasite lytic cycle [19]. In this study, we successfully knocked out the had2a gene in the RH strain and constructed an RHΔhad2a mutant strain. These results of IFA, plaque assay and intravacuolar replication assay demonstrated the important role of HAD2a in daughter budding, the separation of dividing tachyzoites and intracellular proliferation, which was consistent with a previous study [19]. We used 102, 103, 104, 105, and 106 RHΔhad2a tachyzoites and 102 RH tachyzoites to infect Kunming mice and monitored the survival rates of the mice. All infectious doses of RHΔhad2a tachyzoites were not lethal to mice and caused no clinical symptoms of toxoplasmosis, suggesting that the deletion of had2a attenuated the virulence of the RH strain, indicating the possible application of RHΔhad2a as a toxoplasmosis vaccine candidate. Infection of mice with the wild-type RH strain can cause histopathological changes in the liver, spleen, brain and lung of the infected mice [35]. Thus, future studies should examine the histopathological changes in the mice infected with RHΔhad2a tachyzoites to evaluate its safety in mice.
Toxoplasma gondii infection induces a complex protective immunity, including the innate immune response, adaptive immune response, and humoral immunity [36,37]. In this study, we found that RHΔhad2a vaccination induced a significant increase in anti-T. gondii specific antibodies, with significantly higher IgG2a levels than IgG1 levels, indicating that the vaccination of mice with RHΔhad2a provokes a Th1-biased immunity. The increased IgG induced by RHΔhad2a immunization may opsonize T. gondii by phagocytosis, inhibiting the attachment of the parasites to host cells and blocking the parasite invasion to exert their protective effect against T. gondii infection [38]. The host immunity is primarily dependent on T helper 1 (Th1) cell-mediated immunity [8,36,37]. High levels of interleukin-12 (IL-12) and interferon-γ (IFN-γ) are critical for the clearance of tachyzoites during acute infection, and it is essential for the formation of tissue cysts and sustainment of latent infection [8,36,37,39]. Whether RHΔhad2a vaccination induces the production of specific cytokines, such as IFN-γ, TNF-a, IL-12 or MCP-1, requires further studies. In addition, the determination of cytokines and antiparasitic factors, such as reactive nitrogen species and oxygen species, in mice infected with different doses of tachyzoites may help to understand whether the possible protective effect of a live-attenuated strain is dependent on the inoculated dose.
Tachyzoites and bradyzoites are the main infectious stages of T. gondii’s asexual reproductive cycle. Rapidly dividing tachyzoites can disseminate to enormous and distant host tissues and provoke significant immune responses [5]. Host immune defense and drug application can limit tachyzoite growth; however, some tachyzoites overcoming these challenges transform into slowly replicating bradyzoites [5]. Cyst walls protect slowly dividing bradyzoites within tissue cysts to remain dormant in hosts by evading host immune responses, facilitating the establishment of long-term persistent infection [5,40]. Toxoplasma gondii infection commonly occurs in humans via the oral consumption of water, vegetables, fruits contaminated with T. gondii oocysts or undercooked meat containing T. gondii tissue cysts [41]. Thus, challenge infection with tachyzoites, cysts and oocysts after the immunization of mice with live-attenuated T. gondii vaccine strains is important in anti-T. gondii vaccine research.
In this study, we found that the survival rate was significantly higher in vaccinated mice challenged with Pru cysts compared with that of unvaccinated mice. Remarkably, RHΔhad2a vaccination protected all mice from Pru oocyst infection. The brain cyst number of vaccinated mice challenged with Pru cysts and oocysts was significantly decreased comparing to previous data [15,25]. However, T. gondii might be detected in other organs of the infected mice, in which no brain cyst was detected [17]. In addition, the T. gondii load in vaccinated mice should be assessed before and after re-infection, in future studies, to rule out the residual infection and to verify that parasites are effectively eliminated by immunization.
Comparing to the naive group, mice vaccinated with RHΔhad2a tachyzoites had a significantly longer survival time when challenged with tachyzoites of the wild-type I RH strain and PYS strain (ToxoDB#9). These results demonstrate that vaccination with the RHΔhad2a strain conferred partial protection against acute infection and partially prevented the establishment of parasite infection in mice challenged with the Pru strain. Unlike RHΔtkl1 [13], ME49Δα-amy [14] and RHΔgra17Δnpt1 [15], RHΔhad2a did not fully protect mice from the virulent RH and PYS strains. Thus, RHΔhad2a may not be individually applied as an effective live-attenuated vaccine. However, RHΔhad2a may be applied to generate a multiple-attenuated knockout vaccine strain in further studies. One limitation of RHΔhad2a constructed in this study is that it exhibits resistance to pyrimethamine, which is one of the primary drugs for treating toxoplasmosis. Thus, RHΔhad2a should not be used for humans.
5. Conclusions
The findings of the present study demonstrates that the deletion of had2a suppressed the daughter budding and in vitro growth, and it attenuated the virulence of the T. gondii-type I RH strain. A single vaccination intraperitoneally of 5 × 104 RHΔhad2a tachyzoites induces a Th1-skewed immune response in mice. Immunization of the RHΔhad2a mutant strain provides a significantly longer survival time for mice against acute infection by highly virulent tachyzoites compared to the naive mice, partial protection against chronic infection by Pru cysts, and strong protection against Pru oocysts. These findings show that RHΔhad2a could be a promising parent strain to generate a double- or triple- gene knockout mutant strain, or it could be cooperatively applied with another live-attenuated strain as live-attenuated vaccines.
Author Contributions
Conceptualization, X.-Q.Z., X.-N.Z. and M.W.; Formal analysis, H.-S.Z., H.C., C.-X.L., Z.-W.Z. and X.-N.Z.; Funding acquisition, X.-Q.Z. and M.W.; Investigation, H.-S.Z., H.C., C.-X.L. and Z.-W.Z.; Methodology, H.-S.Z. and X.-N.Z.; Project administration, M.W. and X.-Q.Z.; Resources, X.-N.Z.; Supervision, M.W. and X.-Q.Z.; Writing—original draft, H.-S.Z., H.C., C.-X.L. and X.-N.Z.; Writing—review and editing, X.-N.Z. and X.-Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Shanxi Provincial Agricultural and Rural Research Program (Grant No. LXXMsxnd202101), National Natural Science Foundation of China (Grant No. 32172887), the NSFC-Yunnan Joint Fund (Grant No. U2202201), the Fundamental Research Funds of the Chinese Academy of Agricultural Sciences (Grant No. Y2022XK19) and the Special Research Fund of Shanxi Agricultural University for High-level Talents (Grant No. 2021XG001).
Institutional Review Board Statement
The animal study was reviewed and approved by the Animal Ethics Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Permit No. 2020-022).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, Z.D.; Wang, S.C.; Liu, H.H.; Ma, H.Y.; Li, Z.Y.; Wei, F.; Zhu, X.Q.; Liu, Q. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: A systematic review and meta-analysis. Lancet HIV 2017, 4, e177–e188. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Mendez, O.A.; Koshy, A.A. Toxoplasma gondii: Entry, association, and physiological influence on the central nervous system. PLoS Pathog. 2017, 13, e1006351. [Google Scholar] [CrossRef]
- Elmore, S.A.; Jones, J.L.; Conrad, P.A.; Patton, S.; Lindsay, D.S.; Dubey, J.P. Toxoplasma gondii: Epidemiology, feline clinical aspects, and prevention. Trends Parasitol. 2010, 26, 190–196. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Marra, C.M.; Zhu, X.Q. Epidemiology, pathophysiology, diagnosis, and management of cerebral toxoplasmosis. Clin. Microbiol. Rev. 2021, 34, e00115–e00119. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Pappas, G.; Roussos, N.; Falagas, M.E. Toxoplasmosis snapshots: Global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 2009, 39, 1385–1394. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhang, N.Z.; Li, T.T.; He, J.J.; Elsheikha, H.M.; Zhu, X.Q. Advances in the development of anti-Toxoplasma gondii vaccines: Challenges, opportunities, and perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef]
- Elsheikha, H.M. Congenital toxoplasmosis: Priorities for further health promotion action. Public Health 2008, 122, 335–353. [Google Scholar] [CrossRef]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110 Suppl, S11–S16. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Chen, J.; Wang, M.; Petersen, E.; Zhu, X.Q. Vaccines against Toxoplasma gondii: New developments and perspectives. Expert. Rev. Vaccines 2013, 12, 1287–1299. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, H. Moving towards improved vaccines for Toxoplasma gondii. Expert. Opin. Biol. Ther. 2018, 18, 273–280. [Google Scholar] [CrossRef]
- Wang, J.L.; Liang, Q.L.; Li, T.T.; He, J.J.; Bai, M.J.; Cao, X.Z.; Elsheikha, H.M.; Zhu, X.Q. Toxoplasma gondii tkl1 deletion mutant is a promising vaccine against acute, chronic, and congenital toxoplasmosis in mice. J. Immunol. 2020, 204, 1562–1570. [Google Scholar] [CrossRef]
- Yang, J.; Yang, C.; Qian, J.; Li, F.; Zhao, J.; Fang, R. Toxoplasma gondii α-amylase deletion mutant is a promising vaccine against acute and chronic toxoplasmosis. Microb. Biotechnol. 2020, 13, 2057–2069. [Google Scholar] [CrossRef]
- Liang, Q.L.; Sun, L.X.; Elsheikha, H.M.; Cao, X.Z.; Nie, L.B.; Li, T.T.; Li, T.S.; Zhu, X.Q.; Wang, J.L. RHΔgra17Δnpt1 strain of Toxoplasma gondii elicits protective immunity against acute, chronic and congenital Toxoplasmosis in mice. Microorganisms 2020, 8, 352. [Google Scholar] [CrossRef]
- Shen, Y.; Zheng, B.; Sun, H.; Wu, S.; Fan, J.; Ding, J.; Gao, M.; Kong, Q.; Lou, D.; Ding, H.; et al. A live attenuated RHΔompdcΔuprt mutant of Toxoplasma gondii induces strong protective immunity against toxoplasmosis in mice and cats. Infect. Dis. Poverty 2023, 12, 60. [Google Scholar] [CrossRef]
- Wu, M.; Liu, S.; Chen, Y.; Liu, D.; An, R.; Cai, H.; Wang, J.; Zhou, N.; Obed, C.; Han, M.; et al. Live-attenuated ME49Δcdpk3 strain of Toxoplasma gondii protects against acute and chronic toxoplasmosis. NPJ Vaccines 2022, 7, 98. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.B.; Bai, M.J.; Huang, S.Y.; Zhu, X.Q. Live attenuated Pru:Δcdpk2 strain of Toxoplasma gondii protects against acute, chronic, and congenital toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Engelberg, K.; Ivey, F.D.; Lin, A.; Kono, M.; Lorestani, A.; Faugno-Fusci, D.; Gilberger, T.W.; White, M.; Gubbels, M.J. A MORN1-associated HAD phosphatase in the basal complex is essential for Toxoplasma gondii daughter budding. Cell. Microbiol. 2016, 18, 1153–1171. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Liang, Q.L.; Zhang, Z.W.; Wang, M.; Sibley, L.D.; Zhu, X.Q. The protein phosphatase 2A holoenzyme is a key regulator of starch metabolism and bradyzoite differentiation in Toxoplasma gondii. Nat. Commun. 2022, 13, 7560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Li, T.T.; Wang, J.L.; Liang, Q.L.; Zhang, H.S.; Sun, L.X.; Zhu, X.Q. Functional characterization of two thioredoxin proteins of Toxoplasma gondii using the CRISPR-Cas9 system. Front. Vet. Sci. 2020, 7, 614759. [Google Scholar] [CrossRef]
- Cao, X.Z.; Wang, J.L.; Elsheikha, H.M.; Li, T.T.; Sun, L.X.; Liang, Q.L.; Zhang, Z.W.; Lin, R.Q. Characterization of the role of amylo-alpha-1,6-glucosidase protein in the infectivity of Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2019, 9, 418. [Google Scholar] [CrossRef]
- Zheng, X.N.; Wang, J.L.; Elsheikha, H.M.; Wang, M.; Zhang, Z.W.; Sun, L.X.; Wang, X.C.; Zhu, X.Q.; Li, T.T. Functional characterization of 15 novel dense granule proteins in Toxoplasma gondii using the CRISPR-Cas9 system. Microbiol. Spectr. 2023, 11, e0307822. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Zhao, D.Y.; Liang, Q.L.; Elsheikha, H.M.; Wang, M.; Sun, L.X.; Zhang, Z.W.; Chen, X.Q.; Zhu, X.Q.; Wang, J.L. The antioxidant protein glutaredoxin 1 is essential for oxidative stress response and pathogenicity of Toxoplasma gondii. FASEB J. 2023, 37, e22932. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.N.; Sun, L.X.; Elsheikha, H.M.; Li, T.T.; Gao, J.; Wu, X.J.; Zhang, Z.W.; Wang, M.; Fu, B.Q.; Zhu, X.Q.; et al. A newly characterized dense granule protein (GRA76) is important for the growth and virulence of Toxoplasma gondii. Int. J. Parasitol. 2023, 54, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Bai, M.J.; Elsheikha, H.M.; Liang, Q.L.; Li, T.T.; Cao, X.Z.; Zhu, X.Q. Novel roles of dense granule protein 12 (GRA12) in Toxoplasma gondii infection. FASEB J. 2020, 34, 3165–3178. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.L.; Nie, L.B.; Li, T.T.; Elsheikha, H.M.; Sun, L.X.; Zhang, Z.W.; Zhao, D.Y.; Zhu, X.Q.; Wang, J.L. Functional characterization of 17 protein serine/threonine phosphatases in Toxoplasma gondii using CRISPR-Cas9 system. Front. Cell Dev. Biol. 2021, 9, 738794. [Google Scholar] [CrossRef]
- Gao, J.; Wu, X.J.; Zheng, X.N.; Li, T.T.; Kou, Y.J.; Wang, X.C.; Wang, M.; Zhu, X.Q. Functional characterization of eight zinc finger motif-containing proteins in Toxoplasma gondii type I RH strain using the CRISPR-Cas9 system. Pathogens 2023, 12, 1232. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, N.Z.; Zhang, F.K.; Wang, M.; Hu, L.Y.; Zhu, X.Q. Immune response and protective effect against chronic Toxoplasma gondii infection induced by vaccination with a DNA vaccine encoding profilin. BMC Infect. Dis. 2018, 18, 117. [Google Scholar] [CrossRef]
- Villena, I.; Aubert, D.; Gomis, P.; Ferte, H.; Inglard, J.C.; Denis-Bisiaux, H.; Dondon, J.M.; Pisano, E.; Ortis, N.; Pinon, J.M. Evaluation of a strategy for Toxoplasma gondii oocyst detection in water. Appl. Environ. Microbiol. 2004, 70, 4035–4039. [Google Scholar] [CrossRef]
- Yang, W.B.; Wang, J.L.; Gui, Q.; Zou, Y.; Chen, K.; Liu, Q.; Liang, Q.L.; Zhu, X.Q.; Zhou, D.H. Immunization with a live-attenuated RH:ΔNPT1 strain of Toxoplasma gondii induces strong protective immunity against toxoplasmosis in mice. Front. Microbiol. 2019, 10, 1875. [Google Scholar] [CrossRef]
- Matta, S.K.; Rinkenberger, N.; Dunay, I.R.; Sibley, L.D. Toxoplasma gondii infection and its implications within the central nervous system. Nat. Rev. Microbiol. 2021, 19, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Huang, S.Y.; Behnke, M.S.; Chen, K.; Shen, B.; Zhu, X.Q. The past, present, and future of genetic manipulation in Toxoplasma gondii. Trends Parasitol. 2016, 32, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.M.; El-Tantawy, N.L.; Elsawey, A.M.; Abdelsalam, R.A.; Azab, M.S. The course of infection with Toxoplasma gondii RH strain in mice pre-vaccinated with gamma irradiated tachyzoites. Exp. Parasitol. 2019, 205, 107733. [Google Scholar] [CrossRef]
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Pittman, K.J.; Knoll, L.J. Long-term relationships: The complicated interplay between the host and the developmental stages of Toxoplasma gondii during acute and chronic infections. Microbiol. Mol. Biol. Rev. 2015, 79, 387–401. [Google Scholar] [CrossRef]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef]
- Tachibana, Y.; Hashizaki, E.; Sasai, M.; Yamamoto, M. Host genetics highlights IFN-γ-dependent Toxoplasma genes encoding secreted and non-secreted virulence factors in in vivo CRISPR screens. Cell Rep. 2023, 42, 112592. [Google Scholar] [CrossRef]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The bradyzoite: A key developmental stage for the persistence and pathogenesis of toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef]
- Jones, J.L.; Dubey, J.P. Foodborne toxoplasmosis. Clin. Infect. Dis. 2012, 55, 845–851. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).