

Current Perspectives and Challenges of MAIT Cell-Directed Therapy for Tuberculosis Infection

{kind=link}
Abstract
:1. Introduction
2. MAIT Cell Development and the Microbiota
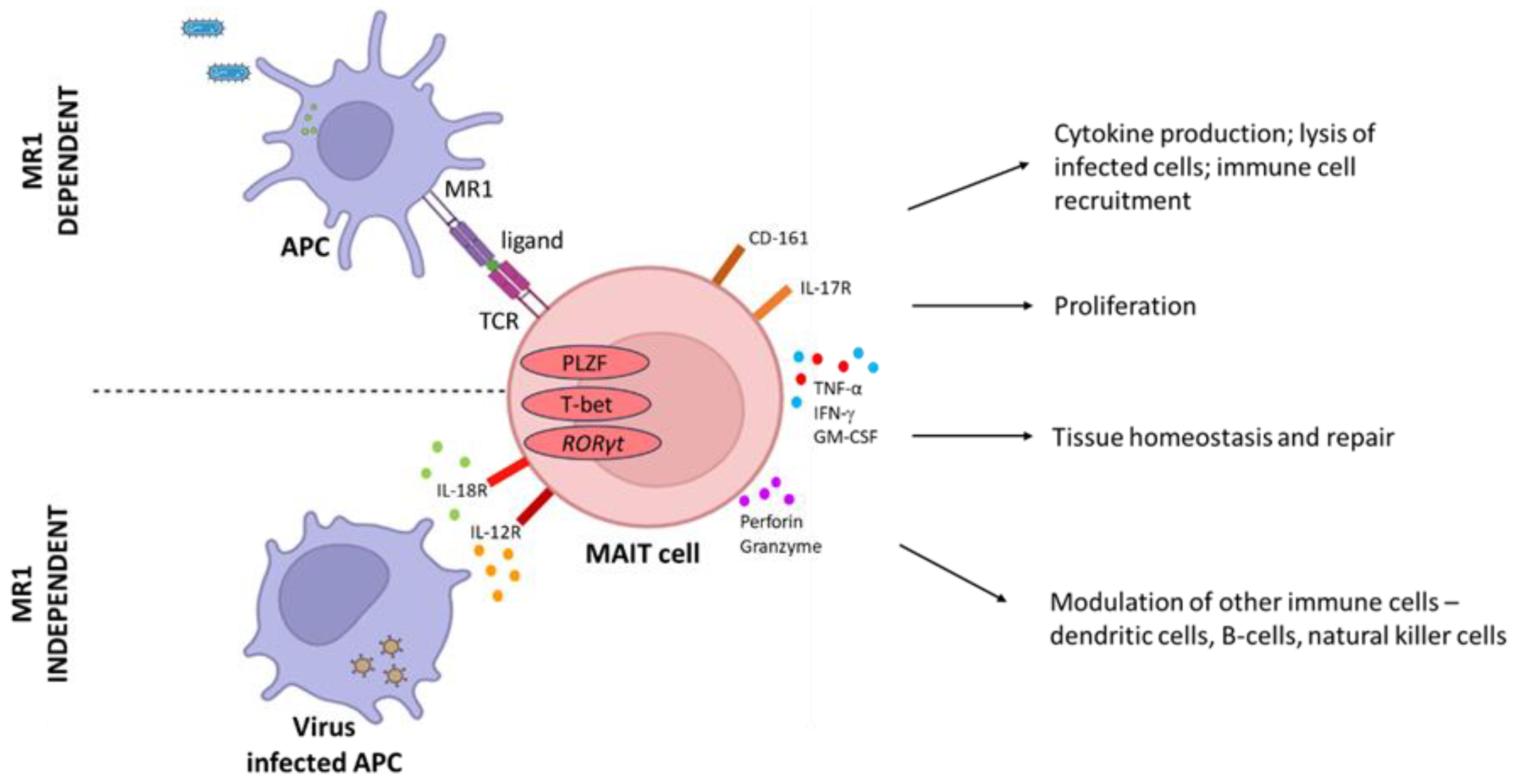
3. Features of MAIT Cells
4. MR1-Bound Antigens
5. MAIT Cell Response to Infection and Disease
6. MAIT Cells and Vaccine Development
7. MAIT Cell Directed Therapy and TB Infection
8. Concluding Remarks and Future Perspectives
Funding
Conflicts of Interest
References
- Gold, M.C.; Lewinsohn, D.M. Co-dependents: MR1-restricted MAIT cells and their antimicrobial function. Nat. Rev. Microbiol. 2013, 11, 14–19. [Google Scholar] [CrossRef]
- Uldrich, A.P.; Le Nours, J.; Pellicci, D.G.; Gherardin, N.A.; McPherson, K.G.; Lim, R.T.; Patel, O.; Beddoe, T.; Gras, S.; Rossjohn, J.; et al. CD1d-lipid antigen recognition by the γδ TCR. Nat. Immunol. 2013, 14, 1137–1145. [Google Scholar] [CrossRef]
- Lantz, O.; Bendelac, A. An invariant T cell receptor alpha chain is used by a unique subset of major histocompatibility complex class I-specific CD4+ and CD4-8- T cells in mice and humans. J. Exp. Med. 1994, 180, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Constant, P.; Davodeau, F.; Peyrat, M.A.; Poquet, Y.; Puzo, G.; Bonneville, M.; Fournié, J.J. Stimulation of human gamma delta T cells by nonpeptidic mycobacterial ligands. Science 1994, 264, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Zinser, M.E.; Highton, A.J.; Kurioka, A.; Kronsteiner, B.; Hagel, J.; Leng, T.; Marchi, E.; Phetsouphanh, C.; Willberg, C.B.; Dunachie, S.J.; et al. Human MAIT cells show metabolic quiescence with rapid glucose-dependent upregulation of granzyme B upon stimulation. Immunol. Cell Biol. 2018, 96, 666–674. [Google Scholar] [CrossRef]
- Lanier, L.L. Shades of grey--the blurring view of innate and adaptive immunity. Nat. Rev. Immunol. 2013, 13, 73–74. [Google Scholar] [CrossRef]
- Gutierrez-Arcelus, M.; Teslovich, N.; Mola, A.R.; Polidoro, R.B.; Nathan, A.; Kim, H.; Hannes, S.; Slowikowski, K.; Watts, G.F.M.; Korsunsky, I.; et al. Lymphocyte innateness defined by transcriptional states reflects a balance between proliferation and effector functions. Nat. Commun. 2019, 10, 687. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.K.; Constantinides, M.G.; Han, J.; Picard, D.; Martin, E.; Li, B.; Lantz, O.; Bendelac, A. The transcription factor PLZF directs the effector program of the NKT cell lineage. Immunity 2008, 29, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.P.; Ives, M.L.; Rao, G.; Lau, A.; Payne, K.; Kobayashi, M.; Arkwright, P.D.; Peake, J.; Wong, M.; Adelstein, S.; et al. STAT3 is a critical cell-intrinsic regulator of human unconventional T cell numbers and function. J. Exp. Med. 2015, 212, 855–864. [Google Scholar] [CrossRef]
- Koay, H.F.; Godfrey, D.I.; Pellicci, D.G. Development of mucosal-associated invariant T cells. Immunol. Cell Biol. 2018, 96, 598–606. [Google Scholar] [CrossRef]
- Dusseaux, M.; Martin, E.; Serriari, N.; Péguillet, I.; Premel, V.; Louis, D.; Milder, M.; Le Bourhis, L.; Soudais, C.; Treiner, E.; et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood 2011, 117, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Legoux, F.; Bellet, D.; Daviaud, C.; El Morr, Y.; Darbois, A.; Niort, K.; Procopio, E.; Salou, M.; Gilet, J.; Ryffel, B.; et al. Microbial metabolites control the thymic development of mucosal-associated invariant T cells. Science 2019, 366, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Leeansyah, E.; Loh, L.; Nixon, D.F.; Sandberg, J.K. Acquisition of innate-like microbial reactivity in mucosal tissues during human fetal MAIT-cell development. Nat. Commun. 2014, 5, 3143. [Google Scholar] [CrossRef]
- Constantinides, M.G.; Link, V.M.; Tamoutounour, S.; Wong, A.C.; Perez-Chaparro, P.J.; Han, S.-J.; Chen, Y.E.; Li, K.; Farhat, S.; Weckel, A.; et al. MAIT cells are imprinted by the microbiota in early life and promote tissue repair. Science 2019, 366, eaax6624. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.Z.; Jo, J.; Tan, A.T.; Sandalova, E.; Chia, A.; Tan, K.C.; Lee, K.H.; Gehring, A.J.; De Libero, G.; Bertoletti, A. IL-7 licenses activation of human liver intrasinusoidal mucosal-associated invariant T cells. J. Immunol. 2013, 190, 3142–3152. [Google Scholar] [CrossRef] [PubMed]
- Reantragoon, R.; Corbett, A.J.; Sakala, I.G.; Gherardin, N.A.; Furness, J.B.; Chen, Z.; Eckle, S.B.; Uldrich, A.P.; Birkinshaw, R.W.; Patel, O.; et al. Antigen-loaded MR1 tetramers define T cell receptor heterogeneity in mucosal-associated invariant T cells. J. Exp. Med. 2013, 210, 2305–2320. [Google Scholar] [CrossRef]
- Tilloy, F.; Treiner, E.; Park, S.H.; Garcia, C.; Lemonnier, F.; de la Salle, H.; Bendelac, A.; Bonneville, M.; Lantz, O. An invariant T cell receptor alpha chain defines a novel TAP-independent major histocompatibility complex class Ib-restricted alpha/beta T cell subpopulation in mammals. J. Exp. Med. 1999, 189, 1907–1921. [Google Scholar] [CrossRef]
- Eckle, S.B.; Birkinshaw, R.W.; Kostenko, L.; Corbett, A.J.; McWilliam, H.E.; Reantragoon, R.; Chen, Z.; Gherardin, N.A.; Beddoe, T.; Liu, L.; et al. A molecular basis underpinning the T cell receptor heterogeneity of mucosal-associated invariant T cells. J. Exp. Med. 2014, 211, 1585–1600. [Google Scholar] [CrossRef]
- Jalali, S.; Harpur, C.M.; Piers, A.T.; Auladell, M.; Perriman, L.; Li, S.; An, K.; Anderson, J.; Berzins, S.P.; Licciardi, P.V.; et al. A high-dimensional cytometry atlas of peripheral blood over the human life span. Immunol. Cell Biol. 2022, 100, 805–821. [Google Scholar] [CrossRef] [PubMed]
- Ussher, J.E.; Bilton, M.; Attwod, E.; Shadwell, J.; Richardson, R.; de Lara, C.; Mettke, E.; Kurioka, A.; Hansen, T.H.; Klenerman, P.; et al. CD161++ CD8+ T cells, including the MAIT cell subset, are specifically activated by IL-12+IL-18 in a TCR-independent manner. Eur. J. Immunol. 2014, 44, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Hackstein, C.P.; Klenerman, P. Emerging features of MAIT cells and other unconventional T cell populations in human viral disease and vaccination. Semin. Immunol. 2022, 61–64, 101661. [Google Scholar] [CrossRef]
- Shilpi, C.; Gabriel, A.; Thomas, R.; Ashu, C.; Ciro, R.-S.; Viankail Cedillo, C.; Gregory, S.; Hayley, S.; Mallory Paynich, M.; Goo-Young, S.; et al. Transcriptomes and metabolism define mouse and human MAIT cell heterogeneity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lu, B.; Liu, M.; Wang, J.; Fan, H.; Yang, D.; Zhang, L.; Gu, X.; Nie, J.; Chen, Z.; Corbett, A.J.; et al. IL-17 production by tissue-resident MAIT cells is locally induced in children with pneumonia. Mucosal Immunol. 2020, 13, 824–835. [Google Scholar] [CrossRef]
- Salio, M.; Gasser, O.; Gonzalez-Lopez, C.; Martens, A.; Veerapen, N.; Gileadi, U.; Verter, J.G.; Napolitani, G.; Anderson, R.; Painter, G.; et al. Activation of Human Mucosal-Associated Invariant T Cells Induces CD40L-Dependent Maturation of Monocyte-Derived and Primary Dendritic Cells. J. Immunol. 2017, 199, 2631–2638. [Google Scholar] [CrossRef]
- Bennett, M.S.; Trivedi, S.; Iyer, A.S.; Hale, J.S.; Leung, D.T. Human mucosal-associated invariant T (MAIT) cells possess capacity for B cell help. J. Leukoc. Biol. 2017, 102, 1261–1269. [Google Scholar] [CrossRef]
- Shimizu, Y.; Horigane-Konakai, Y.; Ishii, Y.; Sugimoto, C.; Wakao, H. Mucosal-associated invariant T cells repress group 2 innate lymphoid cells in Alternaria alternata-induced model of allergic airway inflammation. Front. Immunol. 2022, 13, 1005226. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Miyake, S.; Chiba, A.; Lantz, O.; Yamamura, T. Mucosal-associated invariant T cells regulate Th1 response in multiple sclerosis. Int. Immunol. 2011, 23, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.I.; Ninkov, M.; Haeryfar, S.M.M. Classic costimulatory interactions in MAIT cell responses: From gene expression to immune regulation. Clin. Exp. Immunol. 2023, 213, 50–66. [Google Scholar] [CrossRef]
- Leng, T.; Akther, H.D.; Hackstein, C.P.; Powell, K.; King, T.; Friedrich, M.; Christoforidou, Z.; McCuaig, S.; Neyazi, M.; Arancibia-Cárcamo, C.V.; et al. TCR and Inflammatory Signals Tune Human MAIT Cells to Exert Specific Tissue Repair and Effector Functions. Cell Rep. 2019, 28, 3077–3091.e3075. [Google Scholar] [CrossRef]
- Hinks, T.S.C.; Marchi, E.; Jabeen, M.; Olshansky, M.; Kurioka, A.; Pediongco, T.J.; Meehan, B.S.; Kostenko, L.; Turner, S.J.; Corbett, A.J.; et al. Activation and In Vivo Evolution of the MAIT Cell Transcriptome in Mice and Humans Reveals Tissue Repair Functionality. Cell Rep. 2019, 28, 3249–3262.e3245. [Google Scholar] [CrossRef]
- Rouxel, O.; Da silva, J.; Beaudoin, L.; Nel, I.; Tard, C.; Cagninacci, L.; Kiaf, B.; Oshima, M.; Diedisheim, M.; Salou, M.; et al. Cytotoxic and regulatory roles of mucosal-associated invariant T cells in type 1 diabetes. Nat. Immunol. 2017, 18, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- du Halgouet, A.; Darbois, A.; Alkobtawi, M.; Mestdagh, M.; Alphonse, A.; Premel, V.; Yvorra, T.; Colombeau, L.; Rodriguez, R.; Zaiss, D.; et al. Role of MR1-driven signals and amphiregulin on the recruitment and repair function of MAIT cells during skin wound healing. Immunity 2023, 56, 78–92.e76. [Google Scholar] [CrossRef] [PubMed]
- Kjer-Nielsen, L.; Patel, O.; Corbett, A.J.; Le Nours, J.; Meehan, B.; Liu, L.; Bhati, M.; Chen, Z.; Kostenko, L.; Reantragoon, R.; et al. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 2012, 491, 717–723. [Google Scholar] [CrossRef]
- Keller, A.N.; Eckle, S.B.; Xu, W.; Liu, L.; Hughes, V.A.; Mak, J.Y.; Meehan, B.S.; Pediongco, T.; Birkinshaw, R.W.; Chen, Z.; et al. Drugs and drug-like molecules can modulate the function of mucosal-associated invariant T cells. Nat. Immunol. 2017, 18, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Corbett, A.J.; Eckle, S.B.G.; Birkinshaw, R.W.; Liu, L.; Patel, O.; Mahony, J.; Chen, Z.; Reantragoon, R.; Meehan, B.; Cao, H.; et al. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 2014, 509, 361–365. [Google Scholar] [CrossRef]
- Patel, O.; Kjer-Nielsen, L.; Le Nours, J.; Eckle, S.B.G.; Birkinshaw, R.; Beddoe, T.; Corbett, A.J.; Liu, L.; Miles, J.J.; Meehan, B.; et al. Recognition of vitamin B metabolites by mucosal-associated invariant T cells. Nat. Commun. 2013, 4, 2142. [Google Scholar] [CrossRef]
- Mak, J.Y.; Xu, W.; Reid, R.C.; Corbett, A.J.; Meehan, B.S.; Wang, H.; Chen, Z.; Rossjohn, J.; McCluskey, J.; Liu, L.; et al. Stabilizing short-lived Schiff base derivatives of 5-aminouracils that activate mucosal-associated invariant T cells. Nat. Commun. 2017, 8, 14599. [Google Scholar] [CrossRef]
- Harriff, M.J.; McMurtrey, C.; Froyd, C.A.; Jin, H.; Cansler, M.; Null, M.; Worley, A.; Meermeier, E.W.; Swarbrick, G.; Nilsen, A.; et al. MR1 displays the microbial metabolome driving selective MR1-restricted T cell receptor usage. Sci. Immunol. 2018, 3, eaao2556. [Google Scholar] [CrossRef]
- Schmaler, M.; Colone, A.; Spagnuolo, J.; Zimmermann, M.; Lepore, M.; Kalinichenko, A.; Bhatia, S.; Cottier, F.; Rutishauser, T.; Pavelka, N.; et al. Modulation of bacterial metabolism by the microenvironment controls MAIT cell stimulation. Mucosal Immunol. 2018, 11, 1060–1070. [Google Scholar] [CrossRef]
- Jabeen, M.F.; Hinks, T.S.C. MAIT cells and the microbiome. Front. Immunol. 2023, 14, 1127588. [Google Scholar] [CrossRef]
- Gold, M.C.; Cerri, S.; Smyk-Pearson, S.; Cansler, M.E.; Vogt, T.M.; Delepine, J.; Winata, E.; Swarbrick, G.M.; Chua, W.-J.; Yu, Y.Y.L.; et al. Human Mucosal Associated Invariant T Cells Detect Bacterially Infected Cells. PLoS Biol. 2010, 8, e1000407. [Google Scholar] [CrossRef] [PubMed]
- Booth, J.S.; Salerno-Goncalves, R.; Blanchard, T.G.; Patil, S.A.; Kader, H.A.; Safta, A.M.; Morningstar, L.M.; Czinn, S.J.; Greenwald, B.D.; Sztein, M.B. Mucosal-Associated Invariant T Cells in the Human Gastric Mucosa and Blood: Role in Helicobacter pylori Infection. Front. Immunol. 2015, 6, 466. [Google Scholar] [CrossRef] [PubMed]
- Le Bourhis, L.; Dusseaux, M.; Bohineust, A.; Bessoles, S.; Martin, E.; Premel, V.; Coré, M.; Sleurs, D.; Serriari, N.E.; Treiner, E.; et al. MAIT cells detect and efficiently lyse bacterially-infected epithelial cells. PLoS Pathog. 2013, 9, e1003681. [Google Scholar] [CrossRef]
- Kjer-Nielsen, L.; Corbett, A.J.; Chen, Z.; Liu, L.; Mak, J.Y.W.; Godfrey, D.I.; Rossjohn, J.; Fairlie, D.P.; McCluskey, J.; Eckle, S.B.G. An overview on the identification of MAIT cell antigens. Immunol. Cell Biol. 2018, 96, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Le Bourhis, L.; Martin, E.; Péguillet, I.; Guihot, A.; Froux, N.; Coré, M.; Lévy, E.; Dusseaux, M.; Meyssonnier, V.; Premel, V. Antimicrobial activity of mucosal-associated invariant T cells. Nat. Immunol. 2010, 11, 701–708. [Google Scholar] [CrossRef]
- Hartmann, N.; McMurtrey, C.; Sorensen, M.L.; Huber, M.E.; Kurapova, R.; Coleman, F.T.; Mizgerd, J.P.; Hildebrand, W.; Kronenberg, M.; Lewinsohn, D.M.; et al. Riboflavin Metabolism Variation among Clinical Isolates of Streptococcus pneumoniae Results in Differential Activation of Mucosal-associated Invariant T Cells. Am. J. Respir. Cell. Mol. Biol. 2018, 58, 767–776. [Google Scholar] [CrossRef]
- Meermeier, E.W.; Laugel, B.F.; Sewell, A.K.; Corbett, A.J.; Rossjohn, J.; McCluskey, J.; Harriff, M.J.; Franks, T.; Gold, M.C.; Lewinsohn, D.M. Human TRAV1-2-negative MR1-restricted T cells detect S. pyogenes and alternatives to MAIT riboflavin-based antigens. Nat. Commun. 2016, 7, 12506. [Google Scholar] [CrossRef]
- Meierovics, A.; Yankelevich, W.J.; Cowley, S.C. MAIT cells are critical for optimal mucosal immune responses during in vivo pulmonary bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, E3119–E3128. [Google Scholar] [CrossRef]
- Smith, D.J.; Hill, G.R.; Bell, S.C.; Reid, D.W. Reduced Mucosal Associated Invariant T-Cells Are Associated with Increased Disease Severity and Pseudomonas aeruginosa Infection in Cystic Fibrosis. PLoS ONE 2014, 9, e109891. [Google Scholar] [CrossRef]
- Georgel, P.; Radosavljevic, M.; Macquin, C.; Bahram, S. The non-conventional MHC class I MR1 molecule controls infection by Klebsiella pneumoniae in mice. Mol. Immunol. 2011, 48, 769–775. [Google Scholar] [CrossRef]
- Cho, Y.N.; Kee, S.J.; Kim, T.J.; Jin, H.M.; Kim, M.J.; Jung, H.J.; Park, K.J.; Lee, S.J.; Lee, S.S.; Kwon, Y.S.; et al. Mucosal-associated invariant T cell deficiency in systemic lupus erythematosus. J. Immunol. 2014, 193, 3891–3901. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; Beaudoin, L.; Da Silva, J.; Allatif, O.; Rossjohn, J.; et al. Mucosal-associated invariant T cell alterations in obese and type 2 diabetic patients. J. Clin. Invest. 2015, 125, 1752–1762. [Google Scholar] [CrossRef]
- Grimaldi, D.; Le Bourhis, L.; Sauneuf, B.; Dechartres, A.; Rousseau, C.; Ouaaz, F.; Milder, M.; Louis, D.; Chiche, J.D.; Mira, J.P.; et al. Specific MAIT cell behaviour among innate-like T lymphocytes in critically ill patients with severe infections. Intensive Care. Med. 2014, 40, 192–201. [Google Scholar] [CrossRef]
- Ling, L.; Lin, Y.; Zheng, W.; Hong, S.; Tang, X.; Zhao, P.; Li, M.; Ni, J.; Li, C.; Wang, L.; et al. Circulating and tumor-infiltrating mucosal associated invariant T (MAIT) cells in colorectal cancer patients. Sci. Rep. 2016, 6, 20358. [Google Scholar] [CrossRef] [PubMed]
- Treiner, E.; Liblau, R.S. Mucosal-Associated Invariant T Cells in Multiple Sclerosis: The Jury is Still Out. Front. Immunol. 2015, 6, 503. [Google Scholar] [CrossRef]
- Ussher, J.E.; Willberg, C.B.; Klenerman, P. MAIT cells and viruses. Immunol. Cell Biol. 2018, 96, 630–641. [Google Scholar] [CrossRef] [PubMed]
- van Wilgenburg, B.; Scherwitzl, I.; Hutchinson, E.C.; Leng, T.; Kurioka, A.; Kulicke, C.; de Lara, C.; Cole, S.; Vasanawathana, S.; Limpitikul, W.; et al. MAIT cells are activated during human viral infections. Nat. Commun. 2016, 7, 11653. [Google Scholar] [CrossRef]
- Wallington, J.C.; Williams, A.P.; Staples, K.J.; Wilkinson, T.M.A. IL-12 and IL-7 synergize to control mucosal-associated invariant T-cell cytotoxic responses to bacterial infection. J. Allergy Clin. Immunol. 2018, 141, 2182–2195.e2186. [Google Scholar] [CrossRef]
- Cosgrove, C.; Ussher, J.E.; Rauch, A.; Gärtner, K.; Kurioka, A.; Hühn, M.H.; Adelmann, K.; Kang, Y.H.; Fergusson, J.R.; Simmonds, P.; et al. Early and nonreversible decrease of CD161+ /MAIT cells in HIV infection. Blood 2013, 121, 951–961. [Google Scholar] [CrossRef]
- Leeansyah, E.; Ganesh, A.; Quigley, M.F.; Sönnerborg, A.; Andersson, J.; Hunt, P.W.; Somsouk, M.; Deeks, S.G.; Martin, J.N.; Moll, M.; et al. Activation, exhaustion, and persistent decline of the antimicrobial MR1-restricted MAIT-cell population in chronic HIV-1 infection. Blood 2013, 121, 1124–1135. [Google Scholar] [CrossRef]
- Serriari, N.E.; Eoche, M.; Lamotte, L.; Lion, J.; Fumery, M.; Marcelo, P.; Chatelain, D.; Barre, A.; Nguyen-Khac, E.; Lantz, O.; et al. Innate mucosal-associated invariant T (MAIT) cells are activated in inflammatory bowel diseases. Clin. Exp. Immunol. 2014, 176, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Toubal, A.; Kiaf, B.; Beaudoin, L.; Cagninacci, L.; Rhimi, M.; Fruchet, B.; da Silva, J.; Corbett, A.J.; Simoni, Y.; Lantz, O.; et al. Mucosal-associated invariant T cells promote inflammation and intestinal dysbiosis leading to metabolic dysfunction during obesity. Nat. Commun. 2020, 11, 3755. [Google Scholar] [CrossRef] [PubMed]
- Lezmi, G.; Abou Taam, R.; Dietrich, C.; Chatenoud, L.; de Blic, J.; Leite-de-Moraes, M. Circulating IL-17-producing mucosal-associated invariant T cells (MAIT) are associated with symptoms in children with asthma. Clin. Immunol. 2018, 188, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, M.; Cao, J.; Li, X.; Fan, D.; Xia, Y.; Lu, X.; Li, J.; Ju, D.; Zhao, H. The Dynamic Interplay between the Gut Microbiota and Autoimmune Diseases. J. Immunol. Res. 2019, 2019, 7546047. [Google Scholar] [CrossRef]
- Wong, E.B.; Gold, M.C.; Meermeier, E.W.; Xulu, B.Z.; Khuzwayo, S.; Sullivan, Z.A.; Mahyari, E.; Rogers, Z.; Kløverpris, H.; Sharma, P.K.; et al. TRAV1-2(+) CD8(+) T-cells including oligoconal expansions of MAIT cells are enriched in the airways in human tuberculosis. Commun. Biol. 2019, 2, 203. [Google Scholar] [CrossRef]
- Seshadri, C.; Thuong, N.T.; Mai, N.T.; Bang, N.D.; Chau, T.T.; Lewinsohn, D.M.; Thwaites, G.E.; Dunstan, S.J.; Hawn, T.R. A polymorphism in human MR1 is associated with mRNA expression and susceptibility to tuberculosis. Genes. Immun. 2017, 18, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.; Cords, O.; Liu, Q.; Acuna-Villaorduna, C.; Bonnet, M.; Fox, G.J.; Carvalho, A.C.C.; Chan, P.C.; Croda, J.; Hill, P.C.; et al. Infant BCG vaccination and risk of pulmonary and extrapulmonary tuberculosis throughout the life course: A systematic review and individual participant data meta-analysis. Lancet Glob. Health 2022, 10, e1307–e1316. [Google Scholar] [CrossRef]
- Angelidou, A.; Pittet, L.F.; Faustman, D.; Curtis, N.; Levy, O. BCG vaccine’s off-target effects on allergic, inflammatory, and autoimmune diseases: Worth another shot? J. Allergy Clin. Immunol. 2022, 149, 51–54. [Google Scholar] [CrossRef]
- Kumar, N.P.; Padmapriyadarsini, C.; Rajamanickam, A.; Bhavani, P.K.; Nancy, A.; Jayadeepa, B.; Selvaraj, N.; Asokan, D.; Renji, R.M.; Venkataramani, V.; et al. BCG vaccination induces enhanced frequencies of memory T cells and altered plasma levels of common γc cytokines in elderly individuals. PLoS ONE 2021, 16, e0258743. [Google Scholar] [CrossRef]
- Dey, R.J.; Dey, B.; Harriff, M.; Canfield, E.T.; Lewinsohn, D.M.; Bishai, W.R. Augmentation of the Riboflavin-Biosynthetic Pathway Enhances Mucosa-Associated Invariant T (MAIT) Cell Activation and Diminishes Mycobacterium tuberculosis Virulence. mBio 2021, 13, e0386521. [Google Scholar] [CrossRef]
- Howson, L.J.; Napolitani, G.; Shepherd, D.; Ghadbane, H.; Kurupati, P.; Preciado-Llanes, L.; Rei, M.; Dobinson, H.C.; Gibani, M.M.; Teng, K.W.W.; et al. MAIT cell clonal expansion and TCR repertoire shaping in human volunteers challenged with Salmonella Paratyphi A. Nat. Commun. 2018, 9, 253. [Google Scholar] [CrossRef]
- Wang, H.; D’Souza, C.; Lim, X.Y.; Kostenko, L.; Pediongco, T.J.; Eckle, S.B.G.; Meehan, B.S.; Shi, M.; Wang, N.; Li, S.; et al. MAIT cells protect against pulmonary Legionella longbeachae infection. Nat. Commun. 2018, 9, 3350. [Google Scholar] [CrossRef] [PubMed]
- Rashu, R.; Ninkov, M.; Wardell, C.M.; Benoit, J.M.; Wang, N.I.; Meilleur, C.E.; D’Agostino, M.R.; Zhang, A.; Feng, E.; Saeedian, N.; et al. Targeting the MR1-MAIT cell axis improves vaccine efficacy and affords protection against viral pathogens. PLoS Pathog. 2023, 19, e1011485. [Google Scholar] [CrossRef] [PubMed]
- Riffelmacher, T.; Paynich Murray, M.; Wientjens, C.; Chandra, S.; Cedillo-Castelán, V.; Chou, T.F.; McArdle, S.; Dillingham, C.; Devereaux, J.; Nilsen, A.; et al. Divergent metabolic programmes control two populations of MAIT cells that protect the lung. Nat. Cell Biol. 2023, 25, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Ledgerwood, J.E.; DeZure, A.D.; Stanley, D.A.; Coates, E.E.; Novik, L.; Enama, M.E.; Berkowitz, N.M.; Hu, Z.; Joshi, G.; Ploquin, A.; et al. Chimpanzee Adenovirus Vector Ebola Vaccine. N. Engl. J. Med. 2014, 376, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 2020, 396, 467–478. [Google Scholar] [CrossRef]
- Barouch, D.H.; Tomaka, F.L.; Wegmann, F.; Stieh, D.J.; Alter, G.; Robb, M.L.; Michael, N.L.; Peter, L.; Nkolola, J.P.; Borducchi, E.N.; et al. Evaluation of a mosaic HIV-1 vaccine in a multicentre, randomised, double-blind, placebo-controlled, phase 1/2a clinical trial (APPROACH) and in rhesus monkeys (NHP 13-19). Lancet 2018, 392, 232–243. [Google Scholar] [CrossRef]
- Provine, N.M.; Amini, A.; Garner, L.C.; Spencer, A.J.; Dold, C.; Hutchings, C.; Silva Reyes, L.; FitzPatrick, M.E.B.; Chinnakannan, S.; Oguti, B.; et al. MAIT cell activation augments adenovirus vector vaccine immunogenicity. Science 2021, 371, 521–526. [Google Scholar] [CrossRef]
- Hartmann, N.; Kronenberg, M. MAITs onstage in mice and men with three acts for development. Immunol. Cell Biol. 2017, 95, 3–4. [Google Scholar] [CrossRef]
- Nelson, A.G.; Wang, H.; Dewar, P.M.; Eddy, E.M.; Li, S.; Lim, X.Y.; Patton, T.; Zhou, Y.; Pediongco, T.J.; Meehan, L.J.; et al. Synthetic 5-amino-6-D-ribitylaminouracil paired with inflammatory stimuli facilitates MAIT cell expansion in vivo. Front. Immunol. 2023, 14, 1109759. [Google Scholar] [CrossRef]
- Cui, Y.; Franciszkiewicz, K.; Mburu, Y.K.; Mondot, S.; Le Bourhis, L.; Premel, V.; Martin, E.; Kachaner, A.; Duban, L.; Ingersoll, M.A. Mucosal-associated invariant T cell–rich congenic mouse strain allows functional evaluation. J. Clin. Investig. 2015, 125, 4171–4185. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.M.; Dash, P.; Roy, S.; McMurtrey, C.; Awad, W.; Reed, J.S.; Hammond, K.B.; Abdulhaqq, S.; Wu, H.L.; Burwitz, B.J.; et al. MR1-restricted mucosal-associated invariant T (MAIT) cells respond to mycobacterial vaccination and infection in nonhuman primates. Mucosal Immunol. 2017, 10, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Kauffman, K.D.; Oh, S.; Nelson, C.E.; Barry, C.E.; Barber, D.L. MAIT cell-directed therapy of Mycobacterium tuberculosis infection. Mucosal Immunol. 2021, 14, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Vorkas, C.K.; Levy, O.; Skular, M.; Li, K.; Aubé, J.; Glickman, M.S. Efficient 5-OP-RU-Induced Enrichment of Mucosa-Associated Invariant T Cells in the Murine Lung Does Not Enhance Control of Aerosol Mycobacterium tuberculosis Infection. Infect. Immun. 2020, 89, 10–1128. [Google Scholar] [CrossRef]
- Yu, H.; Yang, A.; Derrick, S.; Mak, J.Y.W.; Liu, L.; Fairlie, D.P.; Cowley, S. Artificially induced MAIT cells inhibit M. bovis BCG but not M. tuberculosis during in vivo pulmonary infection. Sci. Rep. 2020, 10, 13579. [Google Scholar] [CrossRef]
- Salou, M.; Legoux, F.; Lantz, O. MAIT cell development in mice and humans. Mol. Immunol. 2021, 130, 31–36. [Google Scholar] [CrossRef]
- Bucsan, A.N.; Rout, N.; Foreman, T.W.; Khader, S.A.; Rengarajan, J.; Kaushal, D. Mucosal-activated invariant T cells do not exhibit significant lung recruitment and proliferation profiles in macaques in response to infection with Mycobacterium tuberculosis CDC1551. Tuberculosis 2019, 116s, S11–S18. [Google Scholar] [CrossRef]
- Sakai, S.; Lora, N.E.; Kauffman, K.D.; Dorosky, D.E.; Oh, S.; Namasivayam, S.; Gomez, F.; Fleegle, J.D.; Arlehamn, C.S.L.; Sette, A.; et al. Functional inactivation of pulmonary MAIT cells following 5-OP-RU treatment of non-human primates. Mucosal Immunol. 2021, 14, 1055–1066. [Google Scholar] [CrossRef]
- Balfour, A.; Schutz, C.; Goliath, R.; Wilkinson, K.A.; Sayed, S.; Sossen, B.; Kanyik, J.P.; Ward, A.; Ndzhukule, R.; Gela, A.; et al. Functional and Activation Profiles of Mucosal-Associated Invariant T Cells in Patients With Tuberculosis and HIV in a High Endemic Setting. Front. Immunol. 2021, 12, 648216. [Google Scholar] [CrossRef]
- Wakao, H.; Yoshikiyo, K.; Koshimizu, U.; Furukawa, T.; Enomoto, K.; Matsunaga, T.; Tanaka, T.; Yasutomi, Y.; Yamada, T.; Minakami, H.; et al. Expansion of Functional Human Mucosal-Associated Invariant T Cells via Reprogramming to Pluripotency and Redifferentiation. Cell Stem Cell 2013, 12, 546–558. [Google Scholar] [CrossRef]
- McSharry, B.P.; Samer, C.; McWilliam, H.E.G.; Ashley, C.L.; Yee, M.B.; Steain, M.; Liu, L.; Fairlie, D.P.; Kinchington, P.R.; McCluskey, J.; et al. Virus-Mediated Suppression of the Antigen Presentation Molecule MR1. Cell Rep. 2020, 30, 2948–2962.e2944. [Google Scholar] [CrossRef]
- Shaler, C.R.; Choi, J.; Rudak, P.T.; Memarnejadian, A.; Szabo, P.A.; Tun-Abraham, M.E.; Rossjohn, J.; Corbett, A.J.; McCluskey, J.; McCormick, J.K.; et al. MAIT cells launch a rapid, robust and distinct hyperinflammatory response to bacterial superantigens and quickly acquire an anergic phenotype that impedes their cognate antimicrobial function: Defining a novel mechanism of superantigen-induced immunopathology and immunosuppression. PLoS Biol. 2017, 15, e2001930. [Google Scholar] [CrossRef]
- Jiang, J.; Yang, B.; An, H.; Wang, X.; Liu, Y.; Cao, Z.; Zhai, F.; Wang, R.; Cao, Y.; Cheng, X. Mucosal-associated invariant T cells from patients with tuberculosis exhibit impaired immune response. J. Infect. 2016, 72, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wang, X.; An, H.; Yang, B.; Cao, Z.; Liu, Y.; Su, J.; Zhai, F.; Wang, R.; Zhang, G.; et al. Mucosal-associated invariant T-cell function is modulated by programmed death-1 signaling in patients with active tuberculosis. Am. J. Respir. Crit. Care. Med. 2014, 190, 329–339. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chengalroyen, M.D. Current Perspectives and Challenges of MAIT Cell-Directed Therapy for Tuberculosis Infection. Pathogens 2023, 12, 1343. https://doi.org/10.3390/pathogens12111343
Chengalroyen MD. Current Perspectives and Challenges of MAIT Cell-Directed Therapy for Tuberculosis Infection. Pathogens. 2023; 12(11):1343. https://doi.org/10.3390/pathogens12111343
Chicago/Turabian StyleChengalroyen, Melissa D. 2023. "Current Perspectives and Challenges of MAIT Cell-Directed Therapy for Tuberculosis Infection" Pathogens 12, no. 11: 1343. https://doi.org/10.3390/pathogens12111343
APA StyleChengalroyen, M. D. (2023). Current Perspectives and Challenges of MAIT Cell-Directed Therapy for Tuberculosis Infection. Pathogens, 12(11), 1343. https://doi.org/10.3390/pathogens12111343