
Occurrence and Phylogenetic Analysis of Zoonotic Enteropathogenic Protist Parasites in Asymptomatic Domestic Ruminants from Portugal

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
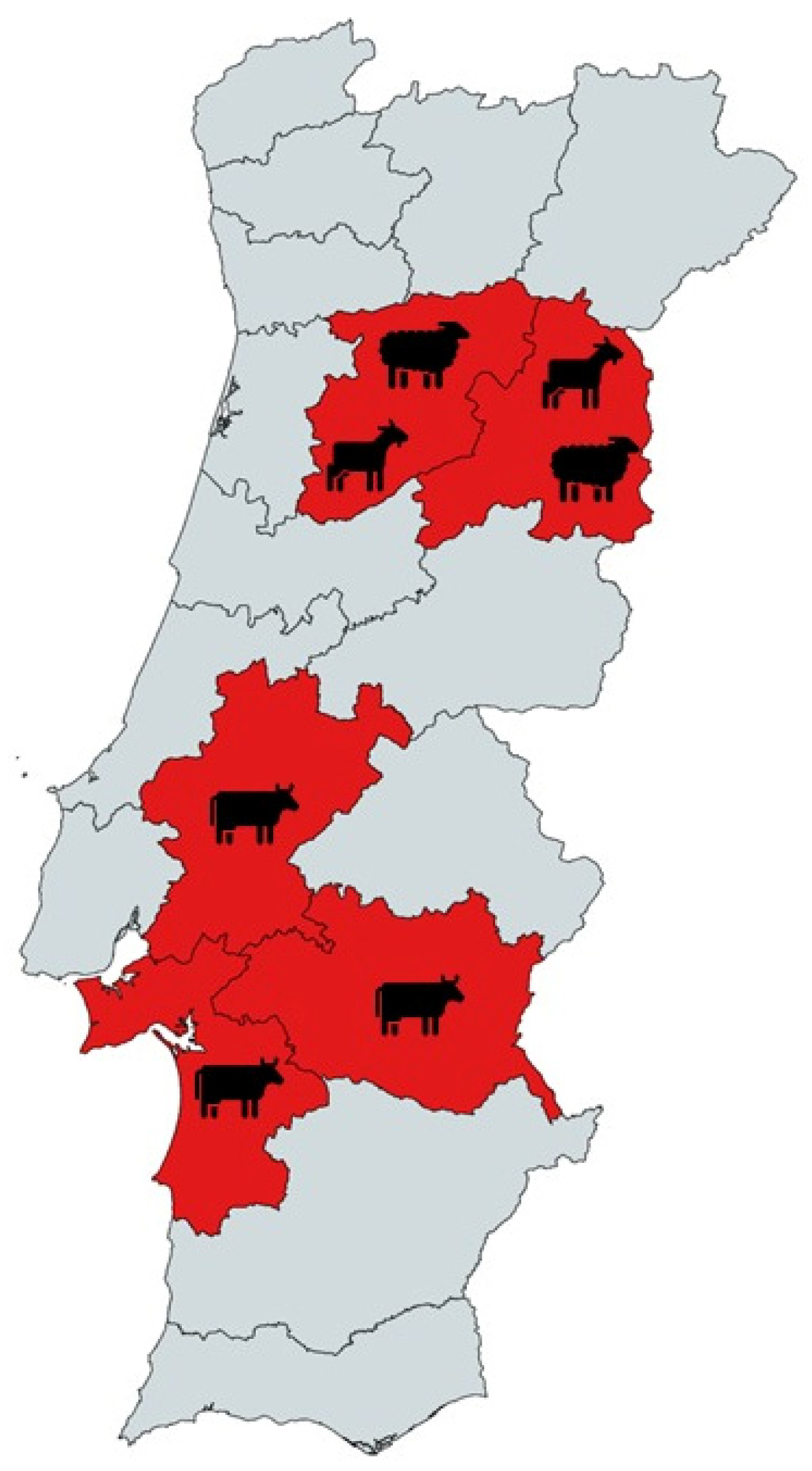
2.1. Sample Collection
2.2. DNA Extraction
2.3. Molecular Detection of Cryptosporidium spp.
2.4. Molecular Detection of Giardia duodenalis
2.5. Molecular Detection of Enterocytozoon bieneusi
2.6. Molecular Detection of Blastocystis sp.
2.7. Molecular Detection of Balantioides coli
2.8. General Procedures
2.9. Statistical Analysis
2.10. Sequencing and Phylogenetic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fayer, R.; Santin, M.; Macarisin, D. Detection of Concurrent Infection of Dairy Cattle with Blastocystis, Cryptosporidium, Giardia, and Enterocytozoon by Molecular and Microscopic Methods. Parasitol. Res. 2012, 111, 1349–1355. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, L.; Liu, L.; Yu, F.; Jian, Y.; Wang, R.; Zhang, S.; Zhang, L.; Ning, C.; Jian, F. Genotyping of Cryptosporidium spp., Giardia duodenalis and Enterocytozoon bieneusi from Sheep and Goats in China. BMC Vet. Res. 2022, 18, 361. [Google Scholar] [CrossRef]
- Percival, S.L.; Yates, M.V.; Williams, D.W.; Chalmers, R.M.; Gray, N.F. Microbiology of Waterborne Diseases: Microbiological Aspects and Risks, 2nd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1–695. [Google Scholar] [CrossRef]
- Ponce-Gordo, F.; García-Rodríguez, J.J. Balantioides coli. Res. Vet. Sci. 2021, 135, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Rauff-Adedotun, A.A.; Meor Termizi, F.H.; Shaari, N.; Lee, I.L. The Coexistence of Blastocystis spp. in Humans, Animals and Environmental Sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Shu, F.; Song, S.; Wei, Y.; Li, F.; Guo, Y.; Feng, Y.; Xiao, L.; Li, N. High Zoonotic Potential of Cryptosporidium spp., Giardia duodenalis, and Enterocytozoon bieneusi in Wild Nonhuman Primates from Yunnan Province, China. Parasites Vectors 2022, 15, 85. [Google Scholar] [CrossRef] [PubMed]
- Abeywardena, H.; Jex, A.R.; Gasser, R.B. A Perspective on Cryptosporidium and Giardia, with an Emphasis on Bovines and Recent Epidemiological Findings. Adv. Parasitol. 2015, 88, 243–301. [Google Scholar] [CrossRef]
- Eurostat Bovine Population—Annual Data—Eurostat. Available online: https://ec.europa.eu/eurostat/databrowser/view/apro_mt_lscatl/default/table?lang=en (accessed on 31 August 2023).
- Eurostat Sheep Population—Annual Data—Eurostat. Available online: https://ec.europa.eu/eurostat/databrowser/view/APRO_MT_LSSHEEP__custom_2702601/bookmark/bar?lang=en&bookmarkId=3f4109cd-c586-4bfe-a21c-4ce42be4ce9a (accessed on 13 October 2023).
- Eurostat Goat Population—Annual Data—Eurostat. Available online: https://ec.europa.eu/eurostat/databrowser/view/APRO_MT_LSGOAT__custom_2702677/bookmark/table?lang=en&bookmarkId=31b55a67-960f-4e32-93bb-e7eecc272b5f (accessed on 13 October 2023).
- Reif, J.S.; Wimmer, L.; Smith, J.A.; Dargatz, D.A.; Cheney, J.M. Human Cryptosporidiosis Associated with an Epizootic in Calves. Am. J. Public Health 1989, 79, 1528–1530. [Google Scholar] [CrossRef]
- Guo, Y.; Li, N.; Ryan, U.; Feng, Y.; Xiao, L. Small Ruminants and Zoonotic Cryptosporidiosis. Parasitol. Res. 2021, 120, 4189–4198. [Google Scholar] [CrossRef]
- Santin, M. Cryptosporidium and Giardia in Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 223–238. [Google Scholar] [CrossRef]
- Thomson, S.; Hamilton, C.A.; Hope, J.C.; Katzer, F.; Mabbott, N.A.; Morrison, L.J.; Innes, E.A. Bovine Cryptosporidiosis: Impact, Host-Parasite Interaction and Control Strategies. Vet. Res. 2017, 48, 42. [Google Scholar] [CrossRef]
- Ayele, A.; Seyoum, Z.; Leta, S. Cryptosporidium Infection in Bovine Calves: Prevalence and Potential Risk Factors in Northwest Ethiopia. BMC Res. Notes 2018, 11, 105. [Google Scholar] [CrossRef]
- Mensah, G.T.; Ayeh-Kumi, P.F.; Annang, A.K.; Owusu-Frimpong, I.; Niampoma, S.; Brown, C.A. Molecular Epidemiology of Cryptosporidium Species in Kpong and Its Environs, Ghana. PLoS ONE 2023, 18, e0281216. [Google Scholar] [CrossRef] [PubMed]
- Malekifard, F.; Ahmadpour, M. Molecular Detection and Identification of Giardia duodenalis in Cattle of Urmia, Northwest of Iran. In Veterinary Research Forum; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2018; Volume 9, p. 81. [Google Scholar]
- Zhao, L.; Zhang, Z.-S.; Han, W.-X.; Yang, B.; Chai, H.-L.; Wang, M.-Y.; Wang, Y.; Zhang, S.; Zhao, W.-H.; Ma, Y.-M. Prevalence and Molecular Characterization of Giardia duodenalis in Dairy Cattle in Central Inner Mongolia, Northern China. Sci. Rep. 2023, 13, 13960. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ma, L.; Gou, J.-M.; Yao, H.-Z.; Ren, M.; Yang, B.-K.; Lin, Q. Seasonal Distribution of Cryptosporidium spp., Giardia duodenalis and Enterocytozoon bieneusi in Tibetan Sheep in Qinghai, China. Parasites Vectors 2022, 15, 394. [Google Scholar] [CrossRef]
- Rojas-López, L.; Marques, R.C.; Svärd, S.G. Giardia duodenalis. Trends Parasitol. 2022, 38, 605–606. [Google Scholar] [CrossRef]
- Sprong, H.; Cacciò, S.M.; van der Giessen, J.W.B.; Partners, Z.N. Identification of Zoonotic Genotypes of Giardia duodenalis. PLoS Negl. Trop. Dis. 2009, 3, e558. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, A.; Sharbatkhori, M.; Tohidi, F.; Ghanbari, M.R.; Karanis, P.; Olfatifar, M.; Majidiani, H.; Khazaei, S.; Bahadory, S.; Javanmard, E. Global Prevalence of Giardia duodenalis in Cattle: A Systematic Review and Meta-Analysis. Prev. Vet. Med. 2022, 203, 105632. [Google Scholar] [CrossRef]
- Adam, R.D. Giardia duodenalis: Biology and Pathogenesis. Clin. Microbiol. Rev. 2021, 34, e00024-19. [Google Scholar] [CrossRef]
- Qin, Y.; Chen, C.; Qin, Y.-F.; Yang, X.-B.; Li, M.-H.; Meng, X.-Z.; Zhao, Z.-Y.; Ma, N.; Cai, Y.; Zhang, Y. Prevalence and Related Factors of Enterocytozoon bieneusi in Cattle: A Global Systematic Review and Meta-Analysis. Prev. Vet. Med. 2022, 208, 105775. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.-S.; Zou, Y.; Li, Z.; Xie, S.-C.; Shi, L.-Q.; Zou, F.-C.; Zhu, X.-Q.; Yang, J.-F.; Zhao, G.-H. Prevalence and Multi-Locus Genotypes of Enterocytozoon bieneusi in Black-Boned Sheep and Goats in Yunnan Province, Southwestern China. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 65, 385–391. [Google Scholar] [CrossRef]
- Zhang, Y.; Koehler, A.V.; Wang, T.; Haydon, S.R.; Gasser, R.B. Enterocytozoon bieneusi Genotypes in Cattle on Farms Located within a Water Catchment Area. J. Eukaryot. Microbiol. 2019, 66, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Feng, Y.; Xiao, L. Enterocytozoon bieneusi. Trends Parasitol. 2022, 38, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Fayer, R. Microsporidiosis: Enterocytozoon bieneusi in Domesticated and Wild Animals. Res. Vet. Sci. 2011, 90, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Greige, S.; El Safadi, D.; Khaled, S.; Gantois, N.; Baydoun, M.; Chemaly, M.; Benamrouz-Vanneste, S.; Chabé, M.; Osman, M.; Certad, G. First Report on the Prevalence and Subtype Distribution of Blastocystis sp. in Dairy Cattle in Lebanon and Assessment of Zoonotic Transmission. Acta Trop. 2019, 194, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, S.-H.; Seo, M.-G.; Kim, H.-Y.; Kim, J.W.; Lee, Y.-R.; Kim, J.H.; Kwon, O.-D.; Kwak, D. Occurrence and Genetic Diversity of Blastocystis in Korean Cattle. Vet. Parasitol. 2018, 258, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Higuera, A.; Herrera, G.; Jimenez, P.; García-Corredor, D.; Pulido-Medellín, M.; Bulla-Castañeda, D.M.; Pinilla, J.C.; Moreno-Pérez, D.A.; Maloney, J.G.; Santín, M.; et al. Identification of Multiple Blastocystis Subtypes in Domestic Animals From Colombia Using Amplicon-Based Next Generation Sequencing. Front. Vet. Sci. 2021, 8, 732129. [Google Scholar] [CrossRef] [PubMed]
- Li, W.C.; Wang, K.; Gu, Y. Occurrence of Blastocystis sp. and Pentatrichomonas Hominis in Sheep and Goats in China. Parasites Vectors 2018, 11, 93. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Tan, K.S.W.; Clark, C.G. Blastocystis. Trends Parasitol. 2020, 36, 315–316. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Karanis, P. Blastocystis spp., ubiquitous parasite of human, animals and environment. In Encyclopedia of Environmental Health; Elsevier: Oxford, UK, 2019; pp. 429–435. ISBN 9780444639523. [Google Scholar]
- Ahmed, A.; Ijaz, M.; Ayyub, R.M.; Ghaffar, A.; Ghauri, H.N.; Aziz, M.U.; Ali, S.; Altaf, M.; Awais, M.; Naveed, M. Balantidium coli in Domestic Animals: An Emerging Protozoan Pathogen of Zoonotic Significance. Acta Trop. 2020, 203, 105298. [Google Scholar] [CrossRef]
- Tiangtip, R.; Jongwutiwes, S. Molecular Analysis of Cryptosporidium Species Isolated from HIV-Infected Patients in Thailand. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Spierling, N.G.; Schmidt, S.; Rosenfeld, U.M.; Reil, D.; Imholt, C.; Jacob, J.; Ulrich, R.G.; Aebischer, T.; Klotz, C. Occurrence and Distribution of Giardia Species in Wild Rodents in Germany. Parasites Vectors 2018, 11, 213. [Google Scholar] [CrossRef] [PubMed]
- Buckholt, M.A.; Lee, J.H.; Tzipori, S. Prevalence of Enterocytozoon bieneusi in Swine: An 18-Month Survey at a Slaughterhouse in Massachusetts. Appl. Environ. Microbiol. 2002, 68, 2595–2599. [Google Scholar] [CrossRef]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA Barcoding of Blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic Heterogeneity in Internal Transcribed Spacer Genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Fan, Y.; Huang, X.; Guo, S.; Yang, F.; Yang, X.; Guo, Y.; Feng, Y.; Xiao, L.; Li, N. Subtyping Cryptosporidium Xiaoi, a Common Pathogen in Sheep and Goats. Pathogens 2021, 10, 800. [Google Scholar] [CrossRef]
- Papanikolopoulou, V.; Lafi, S.Q.; Papadopoulos, E.; Diakou, A.; Xiao, L.; Giadinis, N.D. Risk Factors for Cryptosporidium Infection in Small Ruminants in Northern Greece. Vet. Parasitol. 2022, 309, 109769. [Google Scholar] [CrossRef]
- Delafosse, A.; Castro-Hermida, J.A.; Baudry, C.; Ares-Mazás, E.; Chartier, C. Herd-Level Risk Factors for Cryptosporidium Infection in Dairy-Goat Kids in Western France. Prev. Vet. Med. 2006, 77, 109–121. [Google Scholar] [CrossRef]
- Sanz Ceballos, L.; Illescas Gómez, P.; Sanz Sampelayo, M.R.; Gil Extremera, F.; Rodríguez Osorio, M. Prevalence of Cryptosporidium Infection in Goats Maintained under Semi-Extensive Feeding Conditions in the Southeast of Spain. Parasite 2009, 16, 315–318. [Google Scholar] [CrossRef]
- Li, W.-C.; Wang, K.; Gu, Y.-F. Detection and Genotyping Study of Enterocytozoon bieneusi in Sheep and Goats in East-Central China. Acta Parasitol. 2019, 64, 44–50. [Google Scholar] [CrossRef]
- Wang, S.-S.; Wang, R.-J.; Fan, X.-C.; Liu, T.-L.; Zhang, L.-X.; Zhao, G.-H. Prevalence and Genotypes of Enterocytozoon bieneusi in China. Acta Trop. 2018, 183, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Udonsom, R.; Prasertbun, R.; Mahittikorn, A.; Chiabchalard, R.; Sutthikornchai, C.; Palasuwan, A.; Popruk, S. Identification of Enterocytozoon bieneusi in Goats and Cattle in Thailand. BMC Vet. Res. 2019, 15, 308. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, W.; Yang, D.; Zhang, L.; Wang, R.; Liu, A. Prevalence of Enterocytozoon bieneusi and Genetic Diversity of ITS Genotypes in Sheep and Goats in China. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 32, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, Y.; Wu, Y.; Niu, Z.; Li, J.; Zhang, S.; Wang, R.; Jian, F.; Ning, C.; Zhang, L. Molecular Characterization of Giardia duodenalis and Enterocytozoon bieneusi Isolated from Tibetan Sheep and Tibetan Goats Under Natural Grazing Conditions in Tibet. J. Eukaryot. Microbiol. 2020, 67, 100–106. [Google Scholar] [CrossRef]
- Salehi, R.; Rostami, A.; Mirjalali, H.; Stensvold, C.R.; Haghighi, A. Genetic Characterization of Blastocystis from Poultry, Livestock Animals and Humans in the Southwest Region of Iran-Zoonotic Implications. Transbound. Emerg. Dis. 2022, 69, 1178–1185. [Google Scholar] [CrossRef]
- Onder, Z.; Yildirim, A.; Pekmezci, D.; Duzlu, O.; Pekmezci, G.Z.; Ciloglu, A.; Simsek, E.; Kokcu, N.D.; Yetismis, G.; Ercan, N.; et al. Molecular Identification and Subtype Distribution of Blastocystis sp. in Farm and Pet Animals in Turkey. Acta Trop. 2021, 220, 105939. [Google Scholar] [CrossRef]
- Wei, C.-N.; Qin, R.-L.; Zhang, Z.-H.; Zheng, W.-B.; Liu, Q.; Gao, W.-W.; Zhu, X.-Q.; Xie, S.-C. Prevalence and Genetic Characterization of Blastocystis in Sheep and Pigs in Shanxi Province, North China: From a Public Health Perspective. Animals 2023, 13, 2843. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Berg, R.P.K.D.; Maloney, J.G.; Molokin, A.; Santin, M. Molecular Characterization of Blastocystis and Entamoeba of Muskoxen and Sheep in Greenland. Int. J. Parasitol. 2023, 53, 673–685. [Google Scholar] [CrossRef]
- Tan, T.C.; Tan, P.C.; Sharma, R.; Sugnaseelan, S.; Suresh, K.G. Genetic Diversity of Caprine Blastocystis from Peninsular Malaysia. Parasitol. Res. 2013, 112, 85–89. [Google Scholar] [CrossRef]
- Gabrielli, S.; Palomba, M.; Furzi, F.; Brianti, E.; Gaglio, G.; Napoli, E.; Rinaldi, L.; Alburqueque, R.A.; Mattiucci, S. Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy. Microorganisms 2021, 9, 1656. [Google Scholar] [CrossRef]
- Yu, X.; Wang, H.; Li, Y.; Mu, X.; Yuan, K.; Wu, A.; Guo, J.; Hong, Y.; Zhang, H. Occurrence and Genotypic Identification of Blastocystis spp., Enterocytozoon bieneusi, and Giardia duodenalis in Leizhou Black Goats in Zhanjiang City, Guangdong Province, China. Animals 2023, 13, 2777. [Google Scholar] [CrossRef] [PubMed]
- Al-Herrawy, A.Z.; Gad, M.A. Microsporidial Spores in Fecal Samples of Some Domesticated Animals Living in Giza, Egypt. Iran. J. Parasitol. 2016, 11, 195–203. [Google Scholar]
- Oikarinen, S.; Tauriainen, S.; Viskari, H.; Simell, O.; Knip, M.; Virtanen, S.; Hyöty, H. PCR Inhibition in Stool Samples in Relation to Age of Infants. J. Clin. Virol. 2009, 44, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Dessì, G.; Tamponi, C.; Varcasia, A.; Sanna, G.; Pipia, A.P.; Carta, S.; Salis, F.; Díaz, P.; Scala, A. Cryptosporidium Infections in Sheep Farms from Italy. Parasitol. Res. 2020, 119, 4211–4218. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Navarro, E.; Remesar, S.; García-Dios, D.; Martínez-Calabuig, N.; Prieto, A.; López-Lorenzo, G.; López, C.M.; Panadero, R.; Fernández, G.; et al. The Age-Related Cryptosporidium Species Distribution in Asymptomatic Cattle from North-Western SPAIN. Animals 2021, 11, 256. [Google Scholar] [CrossRef]
- Rzeżutka, A.; Kaupke, A. Occurrence and Molecular Identification of Cryptosporidium Species Isolated from Cattle in Poland. Vet. Parasitol. 2013, 196, 301–306. [Google Scholar] [CrossRef]
- Smith, R.P.; Clifton-Hadley, F.A.; Cheney, T.; Giles, M. Prevalence and Molecular Typing of Cryptosporidium in Dairy Cattle in England and Wales and Examination of Potential On-Farm Transmission Routes. Vet. Parasitol. 2014, 204, 111–119. [Google Scholar] [CrossRef]
- Abarca, N.; Santín, M.; Ortega, S.; Maloney, J.G.; George, N.S.; Molokin, A.; Cardona, G.A.; Dashti, A.; Köster, P.C.; Bailo, B. Molecular Detection and Characterization of Blastocystis sp. and Enterocytozoon bieneusi in Cattle in Northern Spain. Vet. Sci. 2021, 8, 191. [Google Scholar] [CrossRef]
- Abdo, S.M.; El-Adawy, H.; Farag, H.F.; El-Taweel, H.A.; Elhadad, H.; El-Badry, A.A.-M. Detection and Molecular Identification of Blastocystis Isolates from Humans and Cattle in Northern Egypt. J. Parasites Dis. Off. organ Indian Soc. Parasitol. 2021, 45, 738–745. [Google Scholar] [CrossRef]
- Zhu, W.; Tao, W.; Gong, B.; Yang, H.; Li, Y.; Song, M.; Lu, Y.; Li, W. First Report of Blastocystis Infections in Cattle in China. Vet. Parasitol. 2017, 246, 38–42. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Yang, C.; Santin, M.; Matos, O.; Xiao, L. Molecular Characterization of Enterocytozoon bieneusi in Cattle Indicates That Only Some Isolates Have Zoonotic Potential. Parasitol. Res. 2004, 92, 328–334. [Google Scholar] [CrossRef] [PubMed]
- da Silva Fiuza, V.R.; Lopes, C.W.G.; de Oliveira, F.C.R.; Fayer, R.; Santin, M. New Findings of Enterocytozoon bieneusi in Beef and Dairy Cattle in Brazil. Vet. Parasitol. 2016, 216, 46–51. [Google Scholar] [CrossRef]
- Fayer, R.; Santín, M.; Trout, J.M. Enterocytozoon bieneusi in Mature Dairy Cattle on Farms in the Eastern United States. Parasitol. Res. 2007, 102, 15–20. [Google Scholar] [CrossRef]
- Mendonça, C.; Almeida, A.; Castro, A.; de Lurdes Delgado, M.; Soares, S.; da Costa, J.M.C.; Canada, N. Molecular Characterization of Cryptosporidium and Giardia Isolates from Cattle from Portugal. Vet. Parasitol. 2007, 147, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Cardona, G.A.; de Lucio, A.; Bailo, B.; Cano, L.; de Fuentes, I.; Carmena, D. Unexpected Finding of Feline-Specific Giardia duodenalis Assemblage F and Cryptosporidium Felis in Asymptomatic Adult Cattle in Northern Spain. Vet. Parasitol. 2015, 209, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Minetti, C.; Taweenan, W.; Hogg, R.; Featherstone, C.; Randle, N.; Latham, S.M.; Wastling, J.M. Occurrence and Diversity of Giardia duodenalis Assemblages in Livestock in the UK. Transbound. Emerg. Dis. 2014, 61, e60–e67. [Google Scholar] [CrossRef]
- Meng, X.-Z.; Kang, C.; Wei, J.; Ma, H.; Liu, G.; Zhao, J.-P.; Zhang, H.-S.; Yang, X.-B.; Wang, X.-Y.; Yang, L.-H. Meta-Analysis of the Prevalence of Giardia duodenalis in Cattle in China. Foodborne Pathog. Dis. 2023, 20, 17–31. [Google Scholar] [CrossRef]
- Bilal, C.Q.; Khan, M.S.; Avais, M.; Ijaz, M.; Khan, J.A. Prevalence and Chemotherapy of Balantidium coli in Cattle in the River Ravi Region, Lahore (Pakistan). Vet. Parasitol. 2009, 163, 15–17. [Google Scholar] [CrossRef]
- Dey, A.R.; Begum, N.; Biswas, H.; Alam, M.Z. Prevalence and Factors Influencing Gastrointestinal Parasitic Infections in Sheep in Bangladesh. Ann. Parasitol. 2021, 67, 187–194. [Google Scholar] [CrossRef]
- Jamil, M.; Ijaz, M.; Muddassir Ali, M. Prevalence, Hematology and Treatment of Balantidium coli among Small Ruminants in and Around Lahore, Pakistan. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 123–126. [Google Scholar] [CrossRef]
- Sultan, K.; Elmonir, W.; Hegazy, Y. Gastrointestinal Parasites of Sheep in Kafrelsheikh Governorate, Egypt: Prevalence, Control and Public Health Implications. Beni-Suef Univ. J. Basic Appl. Sci. 2016, 5, 79–84. [Google Scholar] [CrossRef][Green Version]
- Stensvold, C.R.; Beser, J.; Ljungström, B.; Troell, K.; Lebbad, M. Low Host-Specific Enterocytozoon bieneusi Genotype BEB6 Is Common in Swedish Lambs. Vet. Parasitol. 2014, 205, 371–374. [Google Scholar] [CrossRef]
- Fiuza, V.R.D.S.; Lopes, C.W.G.; Cosendey, R.I.J.; de Oliveira, F.C.R.; Fayer, R.; Santín, M. Zoonotic Enterocytozoon bieneusi Genotypes Found in Brazilian Sheep. Res. Vet. Sci. 2016, 107, 196–201. [Google Scholar] [CrossRef]
- Valenčáková, A.; Danišová, O. Molecular Characterization of New Genotypes Enterocytozoon bieneusi in Slovakia. Acta Trop. 2019, 191, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Castro-Hermida, J.A.; Almeida, A.; González-Warleta, M.; Correia da Costa, J.M.; Rumbo-Lorenzo, C.; Mezo, M. Occurrence of Cryptosporidium Parvum and Giardia duodenalis in Healthy Adult Domestic Ruminants. Parasitol. Res. 2007, 101, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.-L.; Yan, W.-L.; Wang, J.-M.; Meng, J.-X.; Zhang, M.; Zhao, J.-X.; Shang, K.-M.; Liu, J.; Liu, W.-H. Meta-Analysis of the Prevalence of Giardia duodenalis in Sheep and Goats in China. Microb. Pathog. 2023, 179, 106097. [Google Scholar] [CrossRef] [PubMed]
- Tzanidakis, N.; Sotiraki, S.; Claerebout, E.; Ehsan, A.; Voutzourakis, N.; Kostopoulou, D.; Stijn, C.; Vercruysse, J.; Geurden, T. Occurrence and Molecular Characterization of Giardia duodenalis and Cryptosporidium spp. in Sheep and Goats Reared under Dairy Husbandry Systems in Greece. Parasite 2014, 21, 45. [Google Scholar] [CrossRef]
- Kiani-Salmi, N.; Fattahi-Bafghi, A.; Astani, A.; Sazmand, A.; Zahedi, A.; Firoozi, Z.; Ebrahimi, B.; Dehghani-Tafti, A.; Ryan, U.; Akrami-Mohajeri, F. Molecular Typing of Giardia duodenalis in Cattle, Sheep and Goats in an Arid Area of Central Iran. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 75, 104021. [Google Scholar] [CrossRef]
- Elmadawy, R.S.; Diab, M.S. Prevalence of Balantidium coli and Molecular Analysis of Isospora Oocysts Found in Goats in Qalyobia Governorate, Egypt. Ann. Vet. Anim. Sci. 2017, 4, 8–17. [Google Scholar]
- Ghimire, T.R.; Bhattarai, N. A Survey of Gastrointestinal Parasites of Goats in a Goat Market in Kathmandu, Nepal. J. Parasites Dis. Off. Organ Indian Soc. Parasitol. 2019, 43, 686–695. [Google Scholar] [CrossRef]
- Kanyari, P.W.N.; Kagira, J.M.; Mhoma, R.J. Prevalence and Intensity of Endoparasites in Small Ruminants Kept by Farmers in Kisumu Municipality, Kenya. Livest. Res. Rural Dev. 2009, 21, 202. [Google Scholar]
- Mhoma, J.; Kanyari, P.; Kagira, J. The Prevalence of Gastro-Intestinal Parasites in Goats in Urban and Peri-Urban Areas of Mwanza City, Tanzania. Sci. PArasitol. 2011, 12, 191–196. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Organism | Locus | Oligonucleotide | Sequence (5′–3′) | Reference |
|---|---|---|---|---|
| Blastocystis sp. | ssu rRNA | RD5 | GAGCTTTTTAACTGCAACAACG | [39] |
| BhRDr | ATCTGGTTGATCCTGCCAGT | |||
| Cryptosporidium spp. | ssu rRNA | CR-P1 | CAGGGAGGTAGTGACAAGAA | [36] |
| CRP2 | TCAGCCTTGCGACCATACTC | |||
| CR-P3 | ATTGGAGGGCAAGTCTGGTG | |||
| CPB-DIAGR | TAAGGTGCTGAAGGAGTAAGG | |||
| Giardia duodenalis | ssu rRNA | RH11 | CATCCGGTCGAT CCT GCC and CAT CCG GTT GAT CCT GCC | [37] |
| Gia2150c | CTGCTGCCGTCCTTGGATGT | |||
| RH4 | AGTCGAACCCTGATTCTCCGCCAGG and AGTCAAACCCTGATCCTCCGCCAGG and AGTCGAACCCTGATTCTCCGTCAGG | |||
| Enterocytozoon bieneusi | ITS | EBITS3 | GGTCATAGGGATGAAGAG | [38] |
| EBITS4 | TTCGAGTTCTTTCGCGCTC | |||
| EBITS1 | GCTCTGAATATCTATGGCT | |||
| EBITS2.4 | ATCGCCGACGGATCCAAGTG | |||
| Balantioides coli | ITS | B5D | GAGCTTTTTAACTGCAACAACG | [40] |
| RD5 | ATCTGGTTGATCCTGCCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes-Gonçalves, S.; Palmeira, J.D.; Ferreira, H.; Santos-Silva, S.; Mesquita, J.R. Occurrence and Phylogenetic Analysis of Zoonotic Enteropathogenic Protist Parasites in Asymptomatic Domestic Ruminants from Portugal. Pathogens 2023, 12, 1341. https://doi.org/10.3390/pathogens12111341
Gomes-Gonçalves S, Palmeira JD, Ferreira H, Santos-Silva S, Mesquita JR. Occurrence and Phylogenetic Analysis of Zoonotic Enteropathogenic Protist Parasites in Asymptomatic Domestic Ruminants from Portugal. Pathogens. 2023; 12(11):1341. https://doi.org/10.3390/pathogens12111341
Chicago/Turabian StyleGomes-Gonçalves, Sara, Josman Dantas Palmeira, Helena Ferreira, Sérgio Santos-Silva, and João R. Mesquita. 2023. "Occurrence and Phylogenetic Analysis of Zoonotic Enteropathogenic Protist Parasites in Asymptomatic Domestic Ruminants from Portugal" Pathogens 12, no. 11: 1341. https://doi.org/10.3390/pathogens12111341
APA StyleGomes-Gonçalves, S., Palmeira, J. D., Ferreira, H., Santos-Silva, S., & Mesquita, J. R. (2023). Occurrence and Phylogenetic Analysis of Zoonotic Enteropathogenic Protist Parasites in Asymptomatic Domestic Ruminants from Portugal. Pathogens, 12(11), 1341. https://doi.org/10.3390/pathogens12111341